Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Sarfuddin Azmi | -- | 3875 | 2023-02-01 06:42:38 | | | |
| 2 | Jessie Wu | -1 word(s) | 3874 | 2023-02-02 02:16:50 | | | | |
| 3 | Jessie Wu | Meta information modification | 3874 | 2023-02-02 02:19:42 | | | | |
| 4 | Jessie Wu | + 11 word(s) | 3885 | 2023-02-02 02:25:59 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Shoaib, S.; Ansari, M.A.; Kandasamy, G.; Vasudevan, R.; Hani, U.; Chauhan, W.; Alhumaidi, M.S.; Altammar, K.A.; Azmi, S.; Ahmad, W.; et al. Plant-Based Compounds Against SARS-CoV-2. Encyclopedia. Available online: https://encyclopedia.pub/entry/40705 (accessed on 26 July 2026).
Shoaib S, Ansari MA, Kandasamy G, Vasudevan R, Hani U, Chauhan W, et al. Plant-Based Compounds Against SARS-CoV-2. Encyclopedia. Available at: https://encyclopedia.pub/entry/40705. Accessed July 26, 2026.
Shoaib, Shoaib, Mohammad Azam Ansari, Geetha Kandasamy, Rajalakshimi Vasudevan, Umme Hani, Waseem Chauhan, Maryam S. Alhumaidi, Khadijah A. Altammar, Sarfuddin Azmi, Wasim Ahmad, et al. "Plant-Based Compounds Against SARS-CoV-2" Encyclopedia, https://encyclopedia.pub/entry/40705 (accessed July 26, 2026).
Shoaib, S., Ansari, M.A., Kandasamy, G., Vasudevan, R., Hani, U., Chauhan, W., Alhumaidi, M.S., Altammar, K.A., Azmi, S., Ahmad, W., Wahab, S., & Islam, N. (2023, February 01). Plant-Based Compounds Against SARS-CoV-2. In Encyclopedia. https://encyclopedia.pub/entry/40705
Shoaib, Shoaib, et al. "Plant-Based Compounds Against SARS-CoV-2." Encyclopedia. Web. 01 February, 2023.
Copy Citation
Polyphenols and alkaloids are the most widespread plant-based products with prominent properties including anti-cancer, antioxidant, antimalarial, antiviral, antibacterial, antifungal, anti-diabetic, anti-inflammatory, and anti-dengue effects. Accordingly, these phytochemicals can be promising candidates for discovering effective therapeutic regimens for SARS-CoV-2.
polyphenols
SARS-CoV-2
antiviral
1. Plant Products with Anti-Inflammatory and Antioxidant Potential in Various Diseases
Curcumin has been known for centuries to have anti-inflammatory effects, and extensive studies have shown that curcumin may inhibit inflammation-associated signaling pathways in many pathological conditions [1]. Several studies have shown that IL-6 down-regulation is associated with the therapeutic effects of curcumin [2]. Another study demonstrated that curcumin showed anti-inflammatory effects through the inhibition of tumor necrosis factor (TNF)-α, serumIL-6, and c-reactive protein (CRP) [3]. Another study elucidated the important role of chloroquine and amodiaquine, which earlier had been implicated in the treatment and prevention of malaria and inflammatory disease (Table 1). Both chloroquine and amodiaquine have been shown to inhibit interferon (IFN) production [4]. tumor necrosis factor (TNF)-α, NF-, IL-1, and IL-6 are known key regulators in the pathogenesis of rheumatoid arthritis and other inflammatory diseases, and chloroquine has been shown to inhibit IL-1, IL-6, and TNF- production in human monocytes stimulated with lipopolysaccharide [5]. In contrast, hydroxychloroquine has been shown to inhibit IL-1 and IL-6 production in monocytes while leaving IL-2, IL-4, IFN-, and TNF- production unaffected [6]. In a study, chloroquine and hydroxychloroquine were found to exhibit anti-inflammatory potential against phytohemagglutinin and lipopolysaccharide-stimulated peripheral blood mononuclear cells (PBMC). They discovered that chloroquine and hydroxychloroquine both inhibited IFN- and TNF-production equally [7]. Epigallocatechin gallate (EGCG) attenuates SARS-CoV-2 infection by blocking the interaction of SARS-CoV-2 spike protein receptor-binding domain to human angiotensin-converting enzyme 2 [8].
Anti-inflammatory potential of ascorbic acid has been accomplished by its outstanding power to inhibit the production of IFN-γ while fucoxanthin decreased TNF-α production in human lymphocytes [9]. Ascorbic acid has also been shown to possess immunomodulatory activity, as it inhibits TNF-α and IL-6 production in lipopolysaccharide (LPS)-induced monocytes [10]. Previous studies also indicated the role of ascorbic acid in mitigating the production of TNF-α and IL-6 in LPS-induced macrophages and community-acquired pneumonia [11]. In a few young healthy males, the level of IL-6 was measured before and after the administration of ascorbic acid, and decreased IL-6 production was reported in endothelin-1-induced males [12]. Emodin, an active component of many plants, has previously been reported to prevent the binding of SARS-CoV to ACE2 receptors through the inhibition of spike glycoprotein [13]. Epigallocatechin-3-gallate (EGCG) is a polyphenol found in green tea, and it has been identified with anti-inflammatory, antioxidant, antiviral, and other health-enhancing properties. In vitro studies revealed that polyphenols inhibited the production of many anti-inflammatory cytokines, including IL-6, TNF-α, and IL-8, by inhibiting the cascade that leads to the activation of ERK1/2 and NF-κB [14]. Another study annotated the possible mechanism of EGCG to inhibit Ang II and CRP expression in macrophages [15]. Chemical structures of some of the bioactive compounds active against SARS-CoV-2 and 3C-like proteinase (3CL pro) are presented in Figure 1.
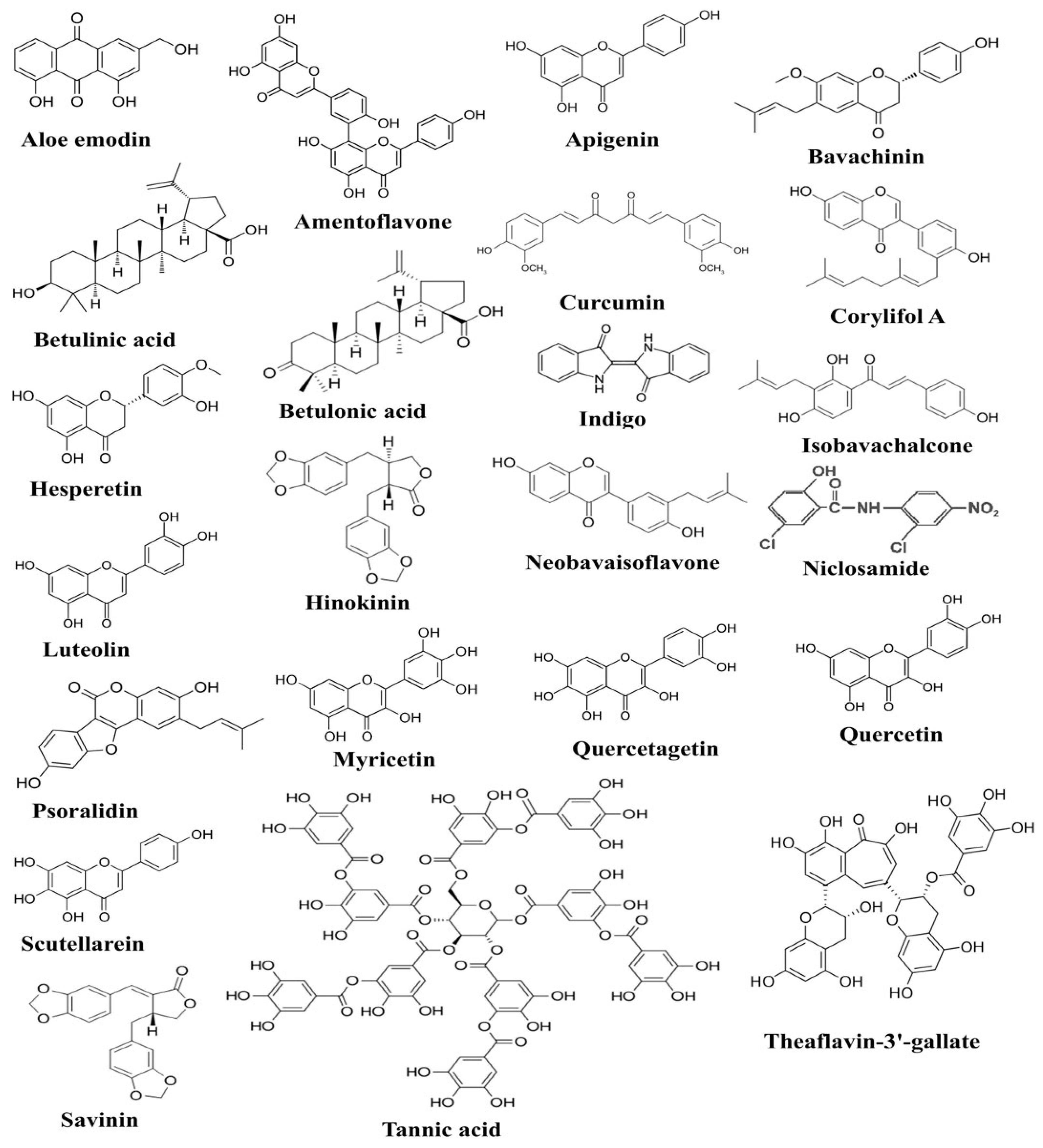
Figure 1. The figure shows chemical structures of plant-derived compounds active against SARS-CoV-2, which have been documented to exert anti-SARS-CoV-2 activities through targeting various pathogenesis-related proteins of SARS-CoV-2 [16].
Polyphenols such as punicalagin, gallic acid, and ellagic acid are well-known antioxidants and have been demonstrated previously to inhibit LPS-stimulated IL-6 production [17]. The recent evidence has shown that punicalagin, a component of pomegranate attenuated the production of IL-6 and TNF-α in lipopolysaccharide (LPS)-induced macrophages [18]. Triphala compounds chebulagic acid, chebulinic acid and gallic acid were also reported to display anti-TNF-α activity in addition to the decreased expression levels of IL-6, IL-8 and MCP-1 which is mediated by inhibition of p38, ERK and NFκB phosphorylation [19]. The anti-inflammatory and antioxidant effects of the phenolic compound resveratrol have been known for decades. In vitro studies reported that resveratrol inhibited TNF-α, IL-8 and IL-6 along with the reduced expression of MCP-1 in human coronary artery smooth muscle cells through the attenuation of extracellular signal-regulated kinase (ERK) [20]. The effect of resveratrol on inflammation and oxidative stress in COPD rats showed that the serum levels of IL-6 and IL-8 were significantly decreased upon treatment with resveratrol [21]. Moreover, the flavonoid quercetin was demonstrated to possess anti-inflammatory and antioxidant potential. Quercetin has been shown to reduce IL-6, TNF-α, and IL-1 production in the LPS-induced macrophage cell line RAW264.77 [22].
Luteolin, a plant product naturally present in several medicinal plants, has attracted wide attention for its health benefits. It significantly reduced the production of TNF-α-induced MCP-1, intercellular cell adhesion molecule (ICAM)-1 and vascular cell adhesion molecule (VCAM)-1 through the inhibition of nuclear factor kappa B (NF-κB) signaling [23]. In vivo studies showed that luteolin and curcumin synergistically inhibited protein expression of MCP-1 and VCAM-1 in TNF-α-induced monocytes [24]. Apigenin, a flavonoid found in many plants, possesses various health benefits, and several studies have elucidated its anti-inflammatory and antioxidant properties. Several studies found its anti-inflammatory effects in various diseases, including diabetes, sepsis, cancer, atherosclerosis, and other inflammation-based diseases. A study demonstrated that apigenin potentially reduced the expression of many inflammatory molecules such as TNF-α, IL-6, IL-8, and GM-CSF through modulating the expression of NF-κB [25][26]. Fisetin, a flavone found in many plants, was discovered to inhibit IL-6, TNF-α, MCP-1, and IL-8 production in IL-1β-stimulated human lung epithelial cells by inhibiting NF-κB and ERK1/2 [27]. Rutin, a flavonoid present in several fruits, vegetables and medicinal plants has been reported to alleviate the expression of TNF-α and nitric oxide in LPS-induced macrophages [28]. In vitro studies of 3,4-dihydroxytoluene, a Rutin metabolite, revealed that it inhibited nitric oxide (NO), cyclooxygenase (COX)-2, and inducible nitric oxide synthase (iNOS) production, as well as the expression of inflammatory cytokines such as IL-6, TNF-α, and IL-1 in LPS-stimulated macrophages [29].
Kaempferol, a flavonoid present in several plant species, has been shown to mollify the secretion of MCP-1 in LPS-induced macrophages [30]. Kaempferol7-O-β-D-glucoside is a natural flavonol glucoside. In vitro studies show that it has the potential to reduce TNF-α, IL-6, IL-1β, iNOS, and COX-2 expression by inhibiting the NF-κB, AP-1, and JAK-STAT signaling pathways in LPS-induced macrophages [31]. Myricetin, a member of the flavonoid class of polyphenols, exhibits antioxidant potential, and the polyphenolic compound has been investigated for its anti-inflammatory properties. A recent study illustrated that myricetin might be responsible for reducing the production of IL-6, TNF-α, NO, and iNOS by suppressing the NF-κB and STAT-1 activation and Nrf-2 mediated Heme oxygenase-1(HO-1) induction in LPS-treated macrophages [32]. Hesperetin, a naturally occurring flavonoid in a few plant species, has been investigated for various health-improving benefits in addition to its antioxidant property. The polyphenolic compound inhibited IL-1β, IL-6 and TNF-α production by modulating the expression of peroxisome proliferator-activated receptor (PPAR)-γ and NF-κB in the bronchoalveolar lavage fluid of rats with ventilator-induced acute lung injury [33]. Another in vitro study suggested that hesperetin treatment may suppress IL-6, TNF-α and IL-1β production and iNOS and COX-2 expression which might be through the inhibition of NF-κB and activation of nuclear factor erythroid 2–related factor 2 (Nrf-2)/HO-1 pathways [34].
Naringenin, a polyphenolic flavonoid found in many citrus fruits and herbs, has been shown to reduce acute inflammation in macrophages by inhibiting IL-6 and TNF-αsecretion [35]. The other study looked into its anti-inflammatory properties and discovered that naringenin inhibited the production of IL-6, TNF-α, and IL-1β. Additionally, abated neutrophil and mononuclear cell recruitment and abrogated myeloperoxidase (MPO) activity were shown to reduce LPS-stimulated inflammatory pain, which possibly occurred through inhibition of the NF-κB pathway [36]. Isorhamnetin was also found to inhibit IL-6 production through the inhibition of NF-κB and STAT-1 signaling pathways in LPS-stimulated macrophages [37]. Some of the flavonoids including chrysin, galangin, quercetin, kaempferol and myricetin were also reported to inhibit IL-1β expression in LPS- and IFN-γ-treated macrophages [38]. Theaflavin, a tea polyphenol, has been known for its antioxidant and anti-cancer properties, and the plant product was also investigated for its anti-inflammatory effects. Theaflavin inhibited LPS-stimulated expression of IL-6, MCP-1, and ICAM-1 by modulating the expression of NF-κB and MAPK [39]. Genistein, an isoflavone mainly present in soybeans, exerts beneficial effects. The isoflavones inhibited IL-6 and TNF-α through suppressing NF-κB pathway activation in LPS-stimulated macrophages [40]. However, attenuated iNOS-derived NO expression was demonstrated in LPS-stimulated macrophages [41]. Baicalein treatment significantly reduced STAT1, STAT3, JAK1 and JAK2 phosphorylation in LPS-induced macrophages which alleviated the production of IL-6, IL-1β and TNF-α and NO [42]. Ferulic acid modulated LPS-induced macrophages expression of anti-inflammatory cytokines such as TNF-α, IL-6 and IL-10 by targeting NF-κB and Nrf-2 pathways [43]. Table 1 shows the anti-inflammatory effects of various phytochemicals.
Table 1. This table shows various plant products which exert anti-inflammatory potential through targeting various inflammation-related signaling molecules. Several phytochemicals and plant extract have been reported to modulate activities of TNF-α, ILs and IFN-γ. Additionally, they also inhibit the binding of spike protein (ACE2 receptor), cyclooxygenase-2, iNOS and CRP production. Thus, plant extracts and phytochemicals can achieve its anti-inflammatory effects against SARS-CoV-2.
| Class | Plant Product | Effects | Reference |
|---|---|---|---|
| Phenolics | Curcumin | Inhibits TNF-α, serum IL-6 and CRP production. | [1][3] |
| Phenolics | Chloroquine | Inhibits IFN-γ, IL-1, IL-6 and TNF-α production. | [4][5] |
| Phenolics | Punicalagin | Inhibits IL-6 and TNF-α production. | [17][18] |
| Phenolics | Ascorbic acid | Inhibits IFN-γ, TNF-α and IL-6 production. | [9][10] |
| Anthraquinone | Emodin | Inhibits binding of spike protein to ACE2 receptor. | [13] |
| Flavanol | EGCG | Inhibits IL-6, TNF-α and IL-8 production. | [14] |
| Phenolics | Hydroxychloroquine | Inhibits IL-1, IL-6 IFN-γ and TNF-α production. | [6][7] |
| Flavone | Luteolin | Inhibits production of TNF-α-induced MCP-1, ICAM-1 and VCAM-1. | [23] |
| Flavone | Apigenin | Inhibits TNF-α, IL-6, IL-8, and GM-CSF expression. | [25][26] |
| Carotenoid | Fucoxanthin | Decreases TNF-α production. | [9] |
| Phenolics | Resveratrol | Inhibits TNF-α, IL-8 and IL-6 and MCP-1 expression. | [20] |
| Phenolics | Gallic acid | Inhibits IL-6 production. | [17] |
| Phenolics | Chebulagic acid (Triphala) | Inhibits IL-6, IL-8 and MCP-1 expression. | [19] |
| Phenolics | Chebulinic acid (Triphala) | Inhibits IL-6, IL-8 and MCP-1 expression. | [19] |
| Flavonol | Quercetin | Inhibits IL-6, TNF-α and IL-1β production. | [22] |
| Flavonol | Fisetin | Inhibits IL-6, TNF-α, MCP-1 and IL-8 production. | [27] |
| Flavone | Rutin | Inhibits expression of NO and TNF-α. | [28] |
| Flavonol | Kaempferol | Inhibits secretion of MCP-1. | [30] |
| Phenolics | Ellagic acid | Inhibits IL-6 production. | [17] |
| Flavonol | Myricetin | Inhibits IL-6, TNF-α, NO, and iNOS. | [32] |
| Flavanone | Hesperetin | Inhibits IL-1β, IL-6 and TNF-α production. | [33][34] |
| Flavanone | Naringenin | Inhibits IL-6 and TNF-α production. | [35] |
| Glycoside | Isorhamnetin | Inhibits IL-6 production. | [37] |
| Flavone | Chrysin | Inhibits IL-1β expression. | [38] |
| Flavonol | Galangin | Inhibits IL-1β expression. | [38] |
| Phenolics | Theaflavin | Inhibits expression of IL-6, MCP-1 and ICAM-1 | [39] |
| Isoflavonoid | Genistein | Inhibits IL-6 and TNF-α production. | [40] |
| Flavone | Baicalein | Inhibits production of IL-6, TNF-α, IL-1β and NO. | [42] |
| Phenolics | Ferulic acid | Inhibits production TNF-α, IL-6 and IL-10 | [43] |
| Flavonol | Kaempferol7-o-β-d-glucoside | Inhibits expression of TNF-α, IL-6, IL-1β, iNOS and COX-2. | [31] |
2. Antiviral Activities of Plant Products: An Insight into Molecular Mechanisms
Various antiviral plant products and their mechanisms of action have been suggested in this section. Several studies have shown the therapeutic potential of many plant products, which include flavonoids, alkaloids, terpenoids, chalcones, glycosides, phenolics, tannins, and lignins. There are many molecular targets that had been anticipated in view of finding a potent target that may lead to the development of a novel strategy for the prevention and treatment of coronaviruses. Various possible molecular targets were identified in order to treat and prevent the previously occurring SARS-CoV and MERS-CoV outbreaks through the identification and application of antiviral plant products. These molecular targets include spike glycoproteins S1 and S2 (domains), virus-encoded proteases such as 3C-like cysteine protease (3CL pro) and papain-like cysteine protease, and virus-encoded enzymes such as RNA-dependent RNA polymerase and NTPase/helicase. Herein, researchers repurposed several plant products to consider them as the possible hope to controlling the novel pandemic, as earlier many plant products such as chloroquine and hydroxychloroquine have proven their anti-malarial potential.
Hesperidin is commonly found in citrus fruits, and its aglycone metabolite is known as hesperetin. In a recent study, hesperetin and hesperedin were considered to determine whether they exhibit inhibitory activities against SARS-CoV-2. The results of the in-silico study showed that both compounds have the ability to bind with TMPRSS2 and ACE2, and they suppressed the infection of VeroE6 cells with SARS-CoV-2 by inhibiting the interaction between S protein and ACE2, which indicates that both compounds can be used for the treatment and prevention of SARS-CoV-2 [44]. Honokiol, the main compound found in Magnolia trees, has previously been shown to have anti-cancer and anti-inflammatory properties. In an interesting study, honokiol was shown to inhibit viral growth and proliferation in cell culture. Notably, honokiol protected VeroE6 cells from the SARS-CoV-2 infection with an EC50 value of 7.8 µM, decreasing the viral RNA copies and viral titers. Further investigations showed that honokiol inhibited SARS-CoV-2 replication, and they also suggested that honokiol is also active against many variants of SARS-CoV-2, including the Omicron variant and other human coronaviruses [45].
In a recent study, Allium sativum, Nigella sativa, Camellia sinensis, Curcuma longa, and Eucalyptus sp. were analyzed for phytochemicals, which revealed that naringenin and saikosaponins show promising anti-SARS-CoV-2 effects [46]. Another study investigated a library of plant products and identified myricetin and scutellarein as the novel inhibitors of the SARS-CoV helicase, nsP13 [47]. Quercetin and epigallocatechin gallate are some of the important flavonoids which played pivotal roles in improving the health status, as well as fighting against many pathogens. They were investigated for their antiviral potential due to antioxidant and anti-inflammation properties and were found to stop SARS-CoV replication via the amelioration of 3CLpro expressed in Pichia pastoris [48]. Ricinus communis, a medicinal plant, is known for its various pharmacological properties, including antioxidation, antibacterial, antiviral, and anti-inflammatory actions, which were studied to determine its anti-SARS-CoV-2 effects. The studies showed that Ricinus communis containsricinine and lupeol as their chief phytoconstituents, and a methylene chloride extract of this plant exerts prominent virucidal activity against SARS-CoV-2. However, they further showed that ricinine exhibited superior anti-SARS-CoV-2 effects with an IC50 value of 2.5 µg/mL, followed by lupeol with an IC50 value of 19.5 µg/mL [49].
In an in-silico study, luteolin was reported to bind with the spike glycoprotein, RdRp, and ADP phosphatase of non-structural protein-3, suggesting luteolin as an anti-SARS-CoV-2 compound [50]. Four flavonoids, amentoflavone, luteolin, apigenin, and quercetin, have been demonstrated to inhibit the SARS-CoV; however, amentoflavone and luteolin significantly inhibited 3CL proteinase (3CL pro) activity at lower doses, comparatively [51]. Another study screened several phytochemicals for their potential activities against 3CL pro and PL pro, and out of 32 phytocompounds, only amentoflavone and gallocatechin gallate were shown to possess the best binding with 3CL pro and PL pro, which may help in the development of amentoflavone and gallocatechin gallate as drug candidates against 3CL pro and PL pro as therapeutics for SARS-CoV-2 [52]. An aqueous extract of Houttuynia cordata was found to be enriched with flavonoids such as quercetin, quercitrin, and isoquercitrin, which demonstrated the inhibition of herpes simplex virus (HSV) and SARS-CoV [53]. Five Isatisindigotica root-extracted compounds and seven plant-derived compounds were evaluated for their anti-SARS-CoV activities, and in vitro studies suggested that hesperetin, sinigrin, and aloe emodin were observed as the most effective inhibitors of cleavage activity of the 3CL pro [54]. Another in silico study has screened 41 antiviral compounds from Indian medicinal plants, and among them, amentoflavone, hypericin, and torvoside H were suggested to serve as potential inhibitors against SARS-CoV-2 [55].
Glycyrrhizin, a compound mainly found in Glycyrrhiza glabra, exerts anti-inflammatory activities and has also been investigated for its anti-SARS-CoV effects. One of the research articles has indicated that glycyrrhizin may inhibit the replication of SARS-CoV at higher doses, which may cause toxicity to normal cells as well [56]. Another study demonstrated that glycyrrhizin and licorice extract possess multiple health benefits, including antioxidant, antibacterial, and anti-inflammatory activities. Furthermore, the researchers showed that glycyrrhizin and licorice extract exhibited significant inhibitory activity against SARS-CoV-2, which was due to the inhibition of viral entry through the inhibition of viral interaction with the cell membrane receptor ACE2 [57]. Cinanserin, a compound naturally available in Houttuynia cordata plant species, inhibited SARS-CoV replication, most likely through the inhibition of 3CL proteinase activity [58]. Lycorine, a compound extracted from the medicinal herb Lycoris radiata, demonstrated anti-SARS-CoV activities [59]. The same research article has explored many medicinal herbs for their effectiveness against the SARS-CoV and reported that out of several, only four Chinese medicinal plants, including Artemisia annua L. (ethanolic extract), Lycoris radiata (ethanolic extract), Pyrrosia lingua (chloroform extract), and Lindera aggregate (sims) kasterm (ethanolic extract) inhibited the activity of the SARS-CoV [59]. Emodin, a compound specifically extracted from the genera Rheum and Polygonum, was found to impede the growth of SARS-CoV through the inhibition of spike proteins and ACE2 interaction [13]. Aqueous extracts of Rheum officinale Baill and Polygonum multiflorum Thunb (Chinese medicinal herbs) significantly inhibited the host–pathogen interaction through the inhibition of spike proteins and ACE2 interactions [13].
Zingiber officinale, Scutellariabaicalensis, Nigella sativa, Allium sativum, Camellia sinensis, Hypericum perforatum, and Glycyrrhiza glabra were studied to identify phytochemicals and their anti-SARS-CoV-2 activities. The results of the study showed that they possess different types of terpenoids, out of which emodin and baicalin inhibited S protein, iguesterin inhibited 3CL pro, cryptotanshinone targeted PL pro and sotetsuflavone inhibited RdRp, thus they exhibited potent anti-SARS-CoV-2 activities [60]. Houttuynia cordata Thunb, a member of the Saururaceae family, was screened to check whether the plant displays anti-SARS-CoV activities. As a result, an aqueous extract of this plant inhibited both 3CL pro and RdRp of SARS-CoV [61]. In order to identify the potential inhibitory effects against SARS-CoV, more than 200 Chinese medicinal herbs were screened, but only a few of them showed antiviral activities. Gentiana scabra, Dioscorea batatas, Cassia tora, Taxillus chinensis, and Cibotium barometz inhibited Vero E6 cell proliferation and viral replication; however, Cibotium barometz and Dioscorea batatas were found to be significant inhibitors of 3CL pro [62]. Rheum palmatum, a Chinese medicinal herb, was also investigated and was found to prevent the replication of SARS-CoV by inhibiting 3CL pro [63]. Toona sinensis Roem, a plant species in the Meliaceae family, was studied for antiviral effects, and the results showed that the plant inhibited SARS-CoV replication in a promising way [64]. Prunella vulgaris and Saussurea lappa were studied among 121 Chinese herbs, and the two compounds tetra-o-galloyl-D-glucose and luteolin extracted from these plants demonstrated antiviral potential by inhibiting spike protein S2 and preventing SARS-CoV entry into Vero E6 cells [65]. Anthemis hyaline, Nigella sativa, and Citrus sinensis extracts have been proven to inhibit the replication of the coronavirus, but the molecular mechanism of the inhibiting replication machinery was not clearly understood [66]. Moreover, recently, two important phytochemicals, curcumin and catechin, were studied for their inhibitory activities against SARS-CoV-2. The results of the in-silico study revealed a high potential to bind with S protein and ACE2, implying that both polyphenolic compounds could be used as anti-SARS-CoV-2 agents [67]. In addition to immunomodulatory, antioxidant, and anti-inflammatory actions, oxidized EGCG also displayed inhibitory activity against SARS-CoV-2 [68]. Furthermore, the flavonoids EGCG and theaflavin were shown to block the virus particle from attaching to the ACE2 and glucose-regulated protein (GRP) 78 receptors. In particular, an in-silico approach revealed that EGCG binds to 3CL pro and both possessed anti-inflammatory properties to control the actions of inflammatory cytokines. All of these experimental results suggest that both flavonoids could be used in the treatment and prevention of SARS-CoV-2 [69]. The impact of bioactive metabolites derived from medicinal plants on molecular targets of various steps of the multiplication process of SARS-CoV-2 has been presented in Figure 2.
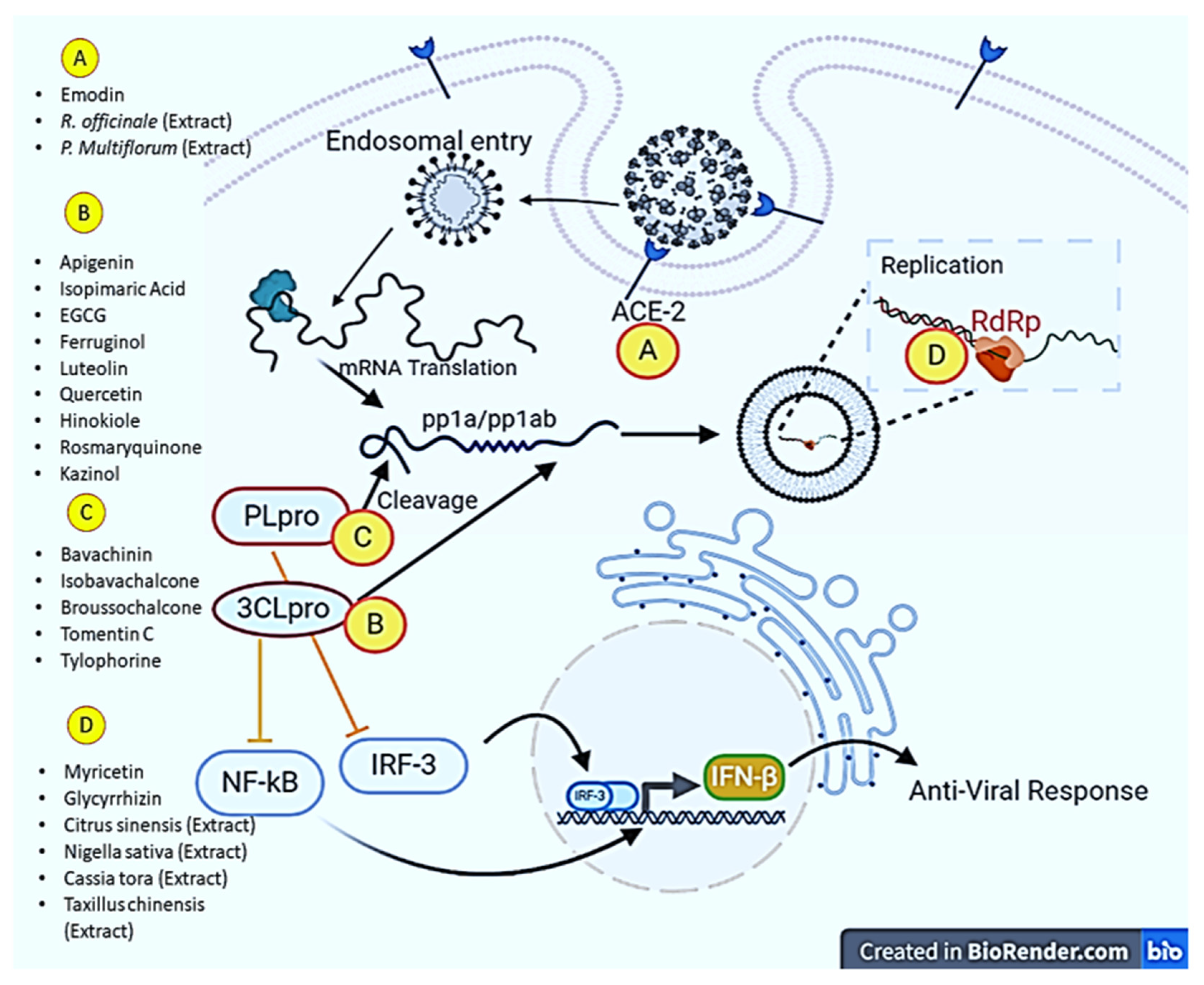
Figure 2. Major molecular targets of phytochemicals in combating SARS-CoV-2. Phytochemicals and plant extracts can target angiotensin-converting enzyme (ACE-2), RNA dependent RNA polymerase (RdRp), Papain-like protease (PL pro) and 3-chymotrypsin-like cysteine protease (3CL pro). The illustration was created at BioRender.com (an online available tool for illustrations).
Recently, an in-silico study revealed that flavonoids such as apigenin, quercetin, and kaempferol bind with the spike receptor protein of SARS-CoV-2; however, quercetin was found to be the most active phytochemical against SARS-CoV-2 and could be a better inhibitor in combating SARS-CoV-2 infections [70]. In silico and in vitro studies suggested that out of many studied phytochemicals, curcumin, EGCG, theaflavin and resveratrol were found to be effective in impeding the coronavirus [71]. Resveratrol, a polyphenolic compound, has been investigated for its antiviral activities against SARS-CoV-2. The results of the study showed that this compound exhibited anti-inflammatory potential and inhibited SARS-CoV-2 replication in human primary bronchial epithelial cell cultures. The low bioavailability (oral administration) of this compound limits its use in clinical practices; however, a topical administration through inhaled formulations may provide a sufficient amount of resveratrol in the lung airways (the site for the entry of SARS-CoV-2) [72]. In an interesting study, black garlic and 49 polyphenolic compounds were screened for their anti-SARS-CoV-2 activities. The results of the study showed that the black garlic extract exerted its inhibitory effect on purified main protease enzyme (Mpro) with an IC50 value of 137 μg/mL, whereas the mixture of tannic acid, daidzein, and puerarin and/or myricetin was found to enhance the inhibitory effects on Mpro [73]. A recent study found that naringenin can be a promising novel pharmacological compound with safe and efficient therapy for SARS-CoV-2 infections, as naringenin strongly inhibited SARS-CoV-2 replication in VeroE6 cells through targeting endo-lysosomal two-pore channels [74].
Some of the biflavonoids and diterpenoids extracted from a traditionally used medicinal plant, Torreya nucifera, were predicted to inhibit the 3CL pro of SARS-CoV. The diterpenoids, ferruginol, o-acetyl-18-hydroxyferruginol, kayadiol and 18-oxoferruginol also demonstrated inhibitory potential against 3CL pro activity of SAS-CoV, slightly more than other isolated diterpenoids such as methyl dehydroabietate, 18-hydroxyferruginol, and hinokiol andisopimaric acid [51]. The geranylated flavonoids extracted from Paulownia tomentosa were found capable of targeting the SARS-CoV papain-like protease (PL pro) in which the compounds tomentin E, tomentin B, and tomentin A were demonstrated as potent inhibitors more comparatively than tomentin C and tomentin D [75]. Many polyphenolic compounds were isolated from the plant species Broussonetia papyrifera for the evaluation of anti-PL pro and anti-3CL pro activities in coronaviruses. Among all of the evaluated compounds, papyriflavonol A, broussochalcone A, broussochalcone B, and kazinol J inhibited SARS-CoV-PL pro more effectively in contrast to other polyphenols including 4-hydroxyisolonchocarpin, 3′-(3-methylbut-2-enyl)-3′,4,7-trihydroxyflavane, kazinol A, kazinol B, broussoflavan A, and kazinol F. However, all the polyphenols were also effective against SARS-CoV 3-chymotripsin-like protease [76]. The ethanolic seed extracts from the medicinal plant Psoralea corylifolia had earlier demonstrated the significant anti-SARS-CoV PL pro potential. The purification process identified many aromatic compounds such as bavachinin, neobavaisoflavone, isobavachalcone, 4′-O-methylbavachalcone, psoralidin and corylifol A, and was reported to be endowed with good PL pro activities. Both the aromatic compounds psoralidin and isobavachalcone displayed potent efficacies in deterioration of the virus replication [77]. Anti-SARS-CoV plant products also comprise of tylophorine compounds purified from Tylophora indica that inhibit the SARS-CoV papain-like protease to curb the viral load via targeting virus replication [78].
With an extending approach in view of repurposing the plant products, Salvia miltiorrhiza-derived tanshinones have been reported as inhibiting the coronaviral cysteine proteases. The plant derived tanshinones include tanshinone IIA, tanshinone IIB, methyl tanshinonate, cryptotanshinone, tanshinone I, dihydrotanshinone I, and rosmariquinone. Dihydrotanshinone I, rosmariquinone, and tanshinone IIB suppressed the viral replication by inhibiting the 3CL pro while cryptotanshinone, tanshinone IIA, and dihydrotanshinone I inhibited 3PL pro activity specifically [79]. Anti-SARS-CoV effects of many medicinal herbs including Cinnamomi cortex (Cinnamomum verum), Forsythiae Fructus (Forsythia suspensa), Scutellariae Radix (Scutellaria baicalensis), Astragali Radix (Astragalus propinquus), Bupleuri Radix (Bupleurum chinensis) and Glycyrrhizae Radix (Glycyrrhiza uralensis) were also envisaged and only a purified compound, procyanidin, and a Cinnamomum verum extract were found to be significantly effective in decreasing coronaviral load while the molecular mechanism was not clearly understood [80]. In an in vitro study, plant-synthesized terpenoids and lignoids were investigated against SARS-CoV replication and proliferation in VeroE6 cells in addition to SARS-CoV 3CL pro. Of the hundreds of plants tested for derived compounds, only ferruginol, 8β-hydroxyabieta-9(11),13-dien-12-one, 7β-hydroxydeoxycryptojaponol, 3β-12-diacetoxyabieta-6,8,11,13-tetraene, betulonic acid, and savinin were found to promote exacerbation of SARS-CoV replication [81].
References
- Fan, Z.; Yao, J.; Li, Y.; Hu, X.; Shao, H.; Tian, X. Anti-inflammatory and antioxidant effects of curcumin on acute lung injury in a rodent model of intestinal ischemia reperfusion by inhibiting the pathway of NF-Kb. Int. J. Clin. Exp. Pathol. 2015, 8, 3451.
- Ghandadi, M.; Sahebkar, A. Curcumin: An effective inhibitor of interleukin-6. Curr. Pharm. Des. 2017, 23, 921–931.
- Mohammadi, S.; Kayedpoor, P.; Karimzadeh-Bardei, L.; Nabiuni, M. The effect of curcumin on TNF-α, IL-6 and CRP expression in a model of polycystic ovary syndrome as an inflammation state. J. Reprod. Infertil. 2017, 18, 352.
- Oh, S.; Shin, J.H.; Jang, E.J.; Won, H.Y.; Kim, H.K.; Jeong, M.G.; Hwang, E.S. Anti-inflammatory activity of chloroquine and amodiaquine through p21-mediated suppression of T cell proliferation and Th1 cell differentiation. Biochem. Biophys. Res. Commun. 2016, 474, 345–350.
- Jang, C.H.; Choi, J.H.; Byun, M.S.; Jue, D.M. Chloroquin inhibits production of TNFalpha, interleukin-1 and interleukin-6 from lipopolysaccharide-stimulated human monocytes/macrophages by different modes. Rheumatology 2006, 45, 703–710.
- Sperber, K.; Quraishi, H.U.M.A.; Kalb, T.H.; Panja, A.S.I.T.; Stecher, V.; Mayer, L. Selective regulation of cytokine secretion by hydroxychloroquine: Inhibition of interleukin 1 alpha (IL-1-alpha) and IL-6 in human monocytes and T cells. J. Rheumatol. 1993, 20, 803–808.
- Dijkmans, B.A.; Verweij, C.L. Chloroquine and hydroxychloroquine equally affect tumor necrosis factor-alpha, interleukin 6, and interferon-gamma production by peripheral blood mononuclear cells. J. Rheumatol. 1997, 24, 55–60.
- Ohishi, T.; Hishiki, T.; Baig, M.S.; Rajpoot, S.; Saqib, U.; Takasaki, T.; Hara, Y. Epigallocatechin gallate (EGCG) attenuates severe acute respiratory coronavirus disease 2 (SARS-CoV-2) infection by blocking the interaction of SARS-CoV-2 spike protein receptor-binding domain to human angiotensin-converting enzyme 2. PLoS ONE 2022, 17, e0271112.
- Molina, N.; Morandi, A.C.; Bolin, A.P.; Otton, R. Comparative effect of fucoxanthin and vitamin C on oxidative and functional parameters of human lymphocytes. Int. Immunol. 2014, 22, 41–50.
- Härtel, C.; Strunk, T.; Bucsky, P.; Schultz, C. Effects of vitamin C on intracytoplasmic cytokine production in human whole blood monocytes and lymphocytes. Cytokine 2004, 27, 101–106.
- Chen, Y.; Luo, G.; Yuan, J.; Wang, Y.; Yang, X.; Wang, X.; Zhong, N. Vitamin C mitigates oxidative stress and tumor necrosis factor-alpha in severe community-acquired pneumonia and LPS-induced macrophages. Mediat. Inflamm. 2014, 2014, 426740.
- Böhm, F.; Settergren, M.; Pernow, J. Vitamin C blocks vascular dysfunction and release of interleukin-6 induced by endothelin-1 in humans in vivo. Atherosclerosis 2007, 190, 408–415.
- Ho, T.Y.; Wu, S.L.; Chen, J.C.; Li, C.C.; Hsiang, C.Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101.
- Shin, H.Y.; Kim, S.H.; Jeong, H.J.; Kim, S.Y.; Shin, T.Y.; Um, J.Y.; Kim, H.M. Epigallocatechin-3-gallate inhibits secretion of TNF-α, IL-6 and IL-8 through the attenuation of ERK and NF-κB in HMC-1 cells. Int. Arch. Allergy Immunol. 2007, 142, 335–344.
- Li, M.; Liu, J.T.; Pang, X.M.; Han, C.J.; Mao, J.J. Epigallocatechin-3-gallate inhibits angiotensin II and interleukin-6-induced C-reactive protein production in macrophages. Pharmacol. Rep. 2012, 64, 912–918.
- Mandal, A.; Jha, A.K.; Hazra, B. Plant products as inhibitors of coronavirus 3CL protease. Front. Pharmacol. 2021, 12, 583387.
- Ben Saad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A&B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47.
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in LPS-Induced RAW264. 7 Macrophages by Inhibiting FoxO3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 2794.
- Shanmuganathan, S.; Angayarkanni, N. Chebulagic acid Chebulinic acid and Gallic acid, the active principles of Triphala, inhibit TNFα induced pro-angiogenic and pro-inflammatory activities in retinal capillary endothelial cells by inhibiting p38, ERK and NF-kB phosphorylation. Vasc. Pharmacol. 2018, 108, 23–35.
- Wakabayashi, I.; Takeda, Y. Inhibitory effects of resveratrol on MCP-1, IL-6, and IL-8 production in human coronary artery smooth muscle cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2013, 386, 835–839.
- Wang, X.L.; Li, T.; Li, J.H.; Miao, S.Y.; Xiao, X.Z. The effects of resveratrol on inflammation and oxidative stress in a rat model of chronic obstructive pulmonary disease. Molecules 2017, 22, 1529.
- Tang, J.; Diao, P.; Shu, X.; Li, L.; Xiong, L. Quercetin and Quercitrin Attenuates the Inflammatory Response and Oxidative Stress in LPS-Induced RAW264. 7 Cells: In vitro Assessment and a Theoretical Model. BioMed Res. Int. 2019, 2019, 7039802.
- Jia, Z.; Nallasamy, P.; Liu, D.; Shah, H.; Li, J.Z.; Chitrakar, R.; Li, Y. Luteolin protects against vascular inflammation in mice and TNF-alpha-induced monocyte adhesion to endothelial cells via suppressing IΚBα/NF-κB signaling pathway. J. Nutr. Biochem. 2015, 26, 293–302.
- Zhang, L.; Wang, X.; Zhang, L.; Virgous, C.; Si, H. Combination of curcumin and luteolin synergistically inhibits TNF-α-induced vascular inflammation in human vascular cells and mice. J. Nutr. Biochem. 2019, 73, 108222.
- Kang, O.H.; Lee, J.H.; Kwon, D.Y. Apigenin inhibits release of inflammatory mediators by blocking the NF-κB activation pathways in the HMC-1 cells. Immunopharmacol. Immunotoxicol. 2011, 33, 473–479.
- Zhang, X.; Wang, G.; Gurley, E.C.; Zhou, H. Flavonoid apigenin inhibits lipopolysaccharide-induced inflammatory response through multiple mechanisms in macrophages. PLoS ONE 2014, 9, e107072.
- Peng, H.L.; Huang, W.C.; Cheng, S.C.; Liou, C.J. Fisetin inhibits the generation of inflammatory mediators in interleukin-1β–induced human lung epithelial cells by suppressing the Nf-κb and Erk1/2 pathways. Int. Immunopharmacol. 2018, 60, 202–210.
- Guruvayoorappan, C.; Kuttan, G. Rutin inhibits nitric oxide and tumor necrosis factor-α production in lipopolysaccharide and concanavalin-A stimulated macrophages. Drug Metab. Drug Interact. 2007, 22, 263–278.
- Su, K.Y.; Yu, C.Y.; Chen, Y.P.; Hua, K.F.; Chen, Y.L.S. 3, 4-Dihydroxytoluene, a metabolite of rutin, inhibits inflammatory responses in lipopolysaccharide-activated macrophages by reducing the activation of NF-κBsignaling. BMC Complement. Altern. Med. 2014, 14, 21.
- Wilczok, T. Effect of kaempferol on the production and gene expression of monocyte chemoattractant protein-1 in J774. 2 macrophages. Pharmacol. Rep. 2005, 57, 107–112.
- Lee, S.B.; Shin, J.S.; Han, H.S.; Lee, H.H.; Park, J.C.; Lee, K.T. Kaempferol 7-O-β-D-glucoside isolated from the leaves of Cudraniatricuspidata inhibits LPS-induced expression of pro-inflammatory mediators through inactivation of NF-κB, AP-1, and JAK-STAT in RAW 264.7 macrophages. Chem.-Biol. Interact. 2018, 284, 101–111.
- Cho, B.O.; Yin, H.H.; Park, S.H.; Byun, E.B.; Ha, H.Y.; Jang, S.I. Anti-inflammatory activity of myricetin from Diospyros lotus through suppression of NF-κB and STAT1 activation and Nrf2-mediated HO-1 induction in lipopolysaccharide-stimulated RAW264. 7 macrophages. Biosci. Biotechnol. Biochem. 2016, 80, 1520–1530.
- Ma, H.; Feng, X.; Ding, S. Hesperetin attenuates ventilator-induced acute lung injury through inhibition of NF-κB-mediated inflammation. Eur. J. Pharmacol. 2015, 769, 333–341.
- Ren, H.; Hao, J.; Liu, T.; Zhang, D.; Lv, H.; Song, E.; Zhu, C. Hesperetin suppresses inflammatory responses in lipopolysaccharide-induced RAW 264.7 cells via the inhibition of NF-κB and activation of Nrf2/HO-1 pathways. Inflammation 2016, 39, 964–973.
- Jin, L.; Zeng, W.; Zhang, F.; Zhang, C.; Liang, W. Naringenin ameliorates acute inflammation by regulating intracellular cytokine degradation. J. Immunol. 2017, 199, 3466–3477.
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Mizokami, S.S.; Borghi, S.M.; Bordignon, J.; Silva, R.L.; VerriJr, W.A. The citrus flavonone naringenin reduces lipopolysaccharide-induced inflammatory pain and leukocyte recruitment by inhibiting NF-κB activation. J. Nutr. Biochem. 2016, 33, 8–14.
- Jin, J.Y.; Choi, E.Y.; Park, H.R.; Choi, J.I.; Choi, I.S.; Kim, S.J. Isorhamnetin inhibits P revotella intermedia lipopolysaccharide-induced production of interleukin-6 in murine macrophages via anti-inflammatory heme oxygenase-1 induction and inhibition of nuclear factor-κB and signal transducer and activator of transcription 1 activation. J. Periodontal Res. 2013, 48, 687–695.
- Bronska, M.; Czuba, Z.P.; Krol, W. Effect of flavone derivatives on interleukin 1β mRNA expression and IL-1β protein synthesis in stimulated RAW 264.7 macrophages. Scand. J. Immunol. 2003, 57, 162–166.
- Kim, S.; Joo, Y.E. Theaflavin inhibits LPS-induced IL-6, MCP-1, and ICAM-1 expression in bone marrow-derived macrophages through the blockade of NF-κB and MAPK signaling pathways. Chonnam Med. J. 2011, 47, 104–110.
- Ji, G.; Zhang, Y.; Yang, Q.; Cheng, S.; Hao, J.; Zhao, X.; Jiang, Z. Genistein suppresses LPS-induced inflammatory response through inhibiting NF-κB following AMP kinase activation in RAW 264.7 macrophages. PLoS ONE 2012, 7, e53101.
- Choi, E.Y.; Bae, S.H.; Ha, M.H.; Choe, S.H.; Hyeon, J.Y.; Choi, J.I.; Kim, S.J. Genistein suppresses Prevotella intermedia lipopolysaccharide-induced inflammatory response in macrophages and attenuates alveolar bone loss in ligature-induced periodontitis. Arch. Oral Biol. 2016, 62, 70–79.
- Qi, Z.; Yin, F.; Lu, L.; Shen, L.; Qi, S.; Lan, L.; Yin, Z. Baicalein reduces lipopolysaccharide-induced inflammation via suppressing JAK/STATs activation and ROS production. Inflamm. Res. 2013, 62, 845–855.
- Lampiasi, N.; Montana, G. The molecular events behind ferulic acid mediated modulation of IL-6 expression in LPS-activated Raw 264.7 cells. Immunobiology 2016, 221, 486–493.
- Cheng, F.J.; Huynh, T.K.; Yang, C.S.; Hu, D.W.; Shen, Y.C.; Tu, C.Y.; Wu, Y.-C.; Tang, C.-H.; Huang, W.-C.; Chen, Y.; et al. Hesperidin Is a Potential Inhibitor against SARS-CoV-2 Infection. Nutrients 2021, 13, 2800.
- Salgado-Benvindo, C.; Leijs, A.A.; Thaler, M.; Tas, A.; Arbiser, J.L.; Snijder, E.J.; van Hemert, M.J. Honokiol inhibits SARS-CoV-2 replication in cell culture. bioRxiv 2022.
- Mitra, S.S.; Nandy, S.; Dey, A. Promising Plant-Based Bioactive Natural Products in Combating SARS-CoV-2 Novel Corona (COVID-19) Virus Infection. In Medicinal Plants for Lung Diseases; Springer: Singapore, 2021; pp. 497–514.
- Yu, M.S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.W.; Jee, J.G.; Jeong, Y.J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorg. Med. Chem. Lett. 2012, 22, 4049–4054.
- Nguyen, T.T.H.; Woo, H.J.; Kang, H.K.; Kim, Y.M.; Kim, D.W.; Ahn, S.A.; Kim, D. Flavonoid-mediated inhibition of SARS coronavirus 3C-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838.
- Elkousy, R.H.; Said, Z.N.; Ali, M.A.; Kutkat, O. Anti-SARS-CoV-2 in vitro potential of Castor Oil Plant (Ricinus communis) Leaf Extract: In-Silico Virtual Evidence. Res. Sq. 2022, 1–34.
- Ansari, W.A.; Ahamad, T.; Khan, M.A.; Khan, Z.A.; Khan, M.F. Exploration of Luteolin as Potential Anti-COVID-19 Agent: Molecular Docking, Molecular Dynamic Simulation, ADMET and DFT Analysis. Lett. Drug Des. Discov. 2022, 19, 741–756.
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.Y.; Kim, D.; Rho, M.C. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947.
- Swargiary, A.; Mahmud, S.; Saleh, M.A. Screening of phytochemicals as potent inhibitor of 3-chymotrypsin and papain-like proteases of SARS-CoV-2: An in silico approach to combat COVID-19. J. Biomol. Struct. Dyn. 2022, 40, 2067–2081.
- Chen, X.; Wang, Z.; Yang, Z.; Wang, J.; Xu, Y.; Tan, R.X.; Li, E. Houttuynia cordata blocks HSV infection through inhibition of NF-κBactivation. Antivir. Res. 2011, 92, 341–345.
- Lin, C.W.; Tsai, F.J.; Tsai, C.H.; Lai, C.C.; Wan, L.; Ho, T.Y.; Chao, P.D.L. Anti-SARS coronavirus 3C-like protease effects of Isatisindigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42.
- Saravanan, K.M.; Zhang, H.; Senthil, R.; Vijayakumar, K.K.; Sounderrajan, V.; Wei, Y.; Shakila, H. Structural basis for the inhibition of SARS-CoV-2 main protease by Indian medicinal plant-derived antiviral compounds. J. Biomol. Struct. Dyn. 2022, 40, 1970–1978.
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046.
- Gomaa, A.A.; Abdel-Wadood, Y.A. The potential of glycyrrhizin and licorice extract in combating COVID-19 and associated conditions. Phytomed. Plus 2021, 1, 100043.
- Chen, L.; Gui, C.; Luo, X.; Yang, Q.; Günther, S.; Scandella, E.; Chen, J. Cinanserin is an inhibitor of the 3C-like proteinase of severe acute respiratory syndrome coronavirus and strongly reduces virus replication in vitro. J. Virol. 2005, 79, 7095–7103.
- Li, S.Y.; Chen, C.; Zhang, H.Q.; Guo, H.Y.; Wang, H.; Wang, L.; Li, R.S. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23.
- Boozari, M.; Hosseinzadeh, H. Natural products for COVID-19 prevention and treatment regarding to previous coronavirus infections and novel studies. Phytother. Res. 2021, 35, 864–876.
- Lau, K.M.; Lee, K.M.; Koon, C.M.; Cheung, C.S.F.; Lau, C.P.; Ho, H.M.; Tsui, S.K.W. Immunomodulatory and anti-SARS activities of Houttuynia cordata. J. Ethnopharmacol. 2008, 118, 79–85.
- Wen, C.C.; Shyur, L.F.; Jan, J.T.; Liang, P.H.; Kuo, C.J.; Arulselvan, P.; Yang, N.S. Traditional Chinese medicine herbal extracts of Cibotium barometz, Gentiana scabra, Dioscorea batatas, Cassia tora, and Taxillus chinensis inhibit SARS-CoVreplication. J. Tradit. Complement. Med. 2011, 1, 41–50.
- Luo, W.; Su, X.; Gong, S.; Qin, Y.; Liu, W.; Li, J.; Xu, Q. Anti-SARS coronavirus 3C-like protease effects of Rheum palmatum L. extracts. Biosci. Trends 2009, 3, 124–126.
- Chen, C.J.; Michaelis, M.; Hsu, H.K.; Tsai, C.C.; Yang, K.D.; Wu, Y.C.; Doerr, H.W. Toona sinensis Roem tender leaf extract inhibits SARS coronavirus replication. J. Ethnopharmacol. 2008, 120, 108–111.
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Chen, L. Small molecules blocking the entry of severe acute respiratory syndrome coronavirus into host cells. J. Virol. 2004, 78, 11334–11339.
- Ulasli, M.; Gurses, S.A.; Bayraktar, R.; Yumrutas, O.; Oztuzcu, S.; Igci, M.; Arslan, A. The effects of Nigella sativa (Ns), Anthemis hyalina(Ah) and Citrus sinensis (Cs) extracts on the replication of coronavirus and the expression of TRP genes family. Mol. Biol. Rep. 2014, 41, 1703–1711.
- Jena, A.B.; Kanungo, N.; Nayak, V.; Chainy, G.B.N.; Dandapat, J. Catechin and curcumin interact with S protein of SARS-CoV-2 and ACE2 of human cell membrane: Insights from computational studies. Sci. Rep. 2021, 11, 2043.
- Ungarala, R.; Munikumar, M.; Sinha, S.N.; Kumar, D.; Sunder, R.S.; Challa, S. Assessment of antioxidant, immunomodulatory activity of oxidised Epigallocatechin-3-Gallate (green tea polyphenol) and its action on the main protease of SARS-CoV-2—An in vitro and in silico approach. Antioxidants 2022, 11, 294.
- Imam, S.S.; Imam, S.T.; Mdwasifathar, K.; Ammar, M.Y. Interaction Between Ace 2 and SARS-CoV-2, And Use of Egcg And Theaflavin to Treat Covid 19 in Initial Phases. Int. J. Curr. Pharm. Res. 2022, 14, 5–10.
- Tuli, H.; Sood, S.; Pundir, A.; Choudhary, D.; Dhama, K.; Kaur, G.; Seth, P.; Vashishth, A.; Kumar, P. Molecular Docking studies of Apigenin, Kaempferol, and Quercetin as potential target against spike receptor protein of SARS COV. J. Exp. Biol. Agric. Sci. 2022, 10, 144–149.
- Flores-Félix, J.D.; Gonçalves, A.C.; Alves, G.; Silva, L.R. Consumption of phenolic-rich food and dietary supplements as a key tool in SARS-CoV-19 infection. Foods 2021, 10, 2084.
- Rossi, G.A.; Sacco, O.; Capizzi, A.; Mastromarino, P. Can resveratrol-inhaled formulations be considered potential adjunct treatments for COVID-19? Front. Immunol. 2021, 12, 670955.
- Nguyen, T.; Jung, J.-H.; Kim, M.-K.; Lim, S.; Choi, J.-M.; Chung, B.; Kim, D.-W.; Kim, D. The inhibitory effects of plant derivate polyphenols on the main protease of SARS coronavirus 2 and their structure–activity relationship. Molecules 2021, 26, 1924.
- Clementi, N.; Scagnolari, C.; D’Amore, A.; Palombi, F.; Criscuolo, E.; Frasca, F.; Pierangeli, A.; Mancini, N.; Antonelli, G.; Clementi, M.; et al. Naringenin is a powerful inhibitor of SARS-CoV-2 infection in vitro. Pharmacol. Res. 2021, 163, 105255.
- Cho, J.K.; Curtis-Long, M.J.; Lee, K.H.; Kim, D.W.; Ryu, H.W.; Yuk, H.J.; Park, K.H. Geranylated flavonoids displaying SARS-CoV papain-like protease inhibition from the fruits of Paulownia tomentosa. Bioorg. Med. Chem. 2013, 21, 3051–3057.
- Park, J.Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Lee, W.S. Evaluation of polyphenols from Broussonetiapapyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–512.
- Kim, D.W.; Seo, K.H.; Curtis-Long, M.J.; Oh, K.Y.; Oh, J.W.; Cho, J.K.; Park, K.H. Phenolic phytochemical displaying SARS-CoV papain-like protease inhibition from the seeds of Psoralea corylifolia. J. Enzym. Inhib. Med. Chem. 2014, 29, 59–63.
- Yang, C.W.; Lee, Y.Z.; Kang, I.J.; Barnard, D.L.; Jan, J.T.; Lin, D.; Lee, S.J. Identification of phenanthroindolizines and phenanthroquinolizidines as novel potent anti-coronaviral agents for porcine enteropathogenic coronavirus transmissible gastroenteritis virus and human severe acute respiratory syndrome coronavirus. Antivir. Res. 2010, 88, 160–168.
- Park, J.Y.; Kim, J.H.; Kim, Y.M.; Jeong, H.J.; Kim, D.W.; Park, K.H.; Ryu, Y.B. Tanshinones as selective and slow-binding inhibitors for SARS-CoV cysteine proteases. Bioorg. Med. Chem. 2012, 20, 5928–5935.
- Zhuang, M.; Jiang, H.; Suzuki, Y.; Li, X.; Xiao, P.; Tanaka, T.; Qin, C. Procyanidins and butanol extract of Cinnamomi Cortex inhibit SARS-CoVinfection. Antivir. Res. 2009, 82, 73–81.
- Wen, C.C.; Kuo, Y.H.; Jan, J.T.; Liang, P.H.; Wang, S.Y.; Liu, H.G.; Hou, C.C. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus. J. Med. Chem. 2007, 50, 4087–4095.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Entry Collection:
Biopharmaceuticals Technology
Revisions:
4 times
(View History)
Update Date:
02 Feb 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No