Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Amjad Shahzad Gondal | -- | 3279 | 2023-01-30 17:40:05 | | | |
| 2 | Lindsay Dong | Meta information modification | 3279 | 2023-01-31 10:37:54 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Khan, A.R.; Mustafa, A.; Hyder, S.; Valipour, M.; Rizvi, Z.F.; Gondal, A.S.; Yousuf, Z.; Iqbal, R.; Daraz, U. Bacillus spp. as Bioagents for Sustainable Agriculture. Encyclopedia. Available online: https://encyclopedia.pub/entry/40608 (accessed on 25 June 2026).
Khan AR, Mustafa A, Hyder S, Valipour M, Rizvi ZF, Gondal AS, et al. Bacillus spp. as Bioagents for Sustainable Agriculture. Encyclopedia. Available at: https://encyclopedia.pub/entry/40608. Accessed June 25, 2026.
Khan, Aimen Razzaq, Adeena Mustafa, Sajjad Hyder, Mohammad Valipour, Zarrin Fatima Rizvi, Amjad Shahzad Gondal, Zubaida Yousuf, Rashid Iqbal, Umar Daraz. "Bacillus spp. as Bioagents for Sustainable Agriculture" Encyclopedia, https://encyclopedia.pub/entry/40608 (accessed June 25, 2026).
Khan, A.R., Mustafa, A., Hyder, S., Valipour, M., Rizvi, Z.F., Gondal, A.S., Yousuf, Z., Iqbal, R., & Daraz, U. (2023, January 30). Bacillus spp. as Bioagents for Sustainable Agriculture. In Encyclopedia. https://encyclopedia.pub/entry/40608
Khan, Aimen Razzaq, et al. "Bacillus spp. as Bioagents for Sustainable Agriculture." Encyclopedia. Web. 30 January, 2023.
Copy Citation
Bacillus spp. improves crop growth in both direct and indirect ways through nitrogen fixation, P and K solubilization, phytohormones production, quorum quenching, biofilm formation, and lytic enzymes production. Moreover, Bacillus spp. boost plant resistance towards the notorious phytopathogens. As Bacillus spp. is eco-friendly, promotes plant growth, confers resistance against diseases, improves soil fertility, non-toxic, naturally occurring microbe, and supports sustainable agriculture, there is a need to explore the potential of native Bacillus spp. and to use them in bioproduct development to support sustainable agriculture.
Bacillus spp.
biocontrol agent
sustainable agriculture
biopesticide
1. Introduction
A range of plant diseases are caused by a variety of pathogenic microorganisms, such as fungi, bacteria, viruses, nematodes, and protozoa, which drastically lower agricultural production and cause significant yield losses [1]. Pathogenic diseases are responsible for between 20–40% of crop productivity losses [2]. Numerous methods have been employed to stop the spread of plant diseases, such as the use of pesticides, crop rotation, less susceptible crops, and other management techniques, but due to the resistance to pesticides and the persistence of soil-borne pathogens, their effectiveness is generally low [3]. Additionally, overusing chemically synthesized fertilizers has not only detrimental impacts on the biosphere but also affects the functioning of the ecosystem and diminishes the sustainability of agriculture [4]. Eco-friendly options for managing plant diseases and boosting crop yields are now being researched and advocated as part of an integrated crop management system—ICMS [5]. Biological control, which is a crucial component of ICMS, is described as the deployment of beneficial microorganisms to lessen the detrimental impacts of phytopathogens and encourage advantageous plant responses [6]. One of the most researched biocontrol agents, as biopesticides, is the Bacillus species, which inhibits phytopathogens by the mechanisms of competition and antagonism [7].
Various microorganisms, including Hypericum gramineum, Pseudomonas fluorescence, and Streptomyces species, have been identified as biocontrol agents [8]. Bacillus species have emerged as an important biological control agent because of their ability to produce antibiotics and tough and resistant endospores to control a range of phytopathogens [9].
Plant growth-promoting attributes have been reported in a variety of Bacillus spp., including B. velezensis, B. subtilis, B. macerans, B. circulans, B. azotofixans, B. coagulans, and others [10]. Phosphate solubilization, nitrogen fixation, production of siderophores, phytohormones, production of antimicrobial compounds, and systemically induced disease resistance are a few of the direct and indirect ways through which Bacillus spp. promote plant growth [11]. Antagonistically important species of the genus Bacillus are growing quickly. Abiotic stress resistance, rapid replication, and a broad spectrum of biocontrol capabilities are all characteristics of Bacillus spp. Volatile organic chemicals produced by B. subtilis are required for stimulating plant development and activating defense mechanisms in plants by boosting the induced systemic resistance (ISR) in plants [12]. Various crops, including tomato, potato, cucumber, maize, common [11] bean, soybean, sunflower, wheat, pepper, and many others, have shown positive impacts of Bacillus spp. on growth and crop yield [13].
2. Microbes as Sources of Biopesticides in Sustainable Agriculture (Biopesticides and Sustainable Agriculture)
Biopesticides are obtained from microorganisms or natural sources [14]. They have been classified as follows; (a) botanical-biopesticides, (b) microbial-biopesticides, and (c) plant-incorporated protectants (PIPs). Sustainable agriculture aims to reduce pests and disease incidence to such an extent that it does not sternly damage crops without disturbing nature’s balance [15]. Although chemical pesticides expeditiously kill a range of agricultural pests, over-dependence on these pesticides has given rise to several problems involving safety risks, environmental pollution, secondary pests breakout, a decrease in biodiversity, and insecticide resistance [16]. Contrary to this, biopesticides are renewable and a significant alternative to conventional pesticides. The sources of microbial-based biopesticides are displayed in Figure 1.
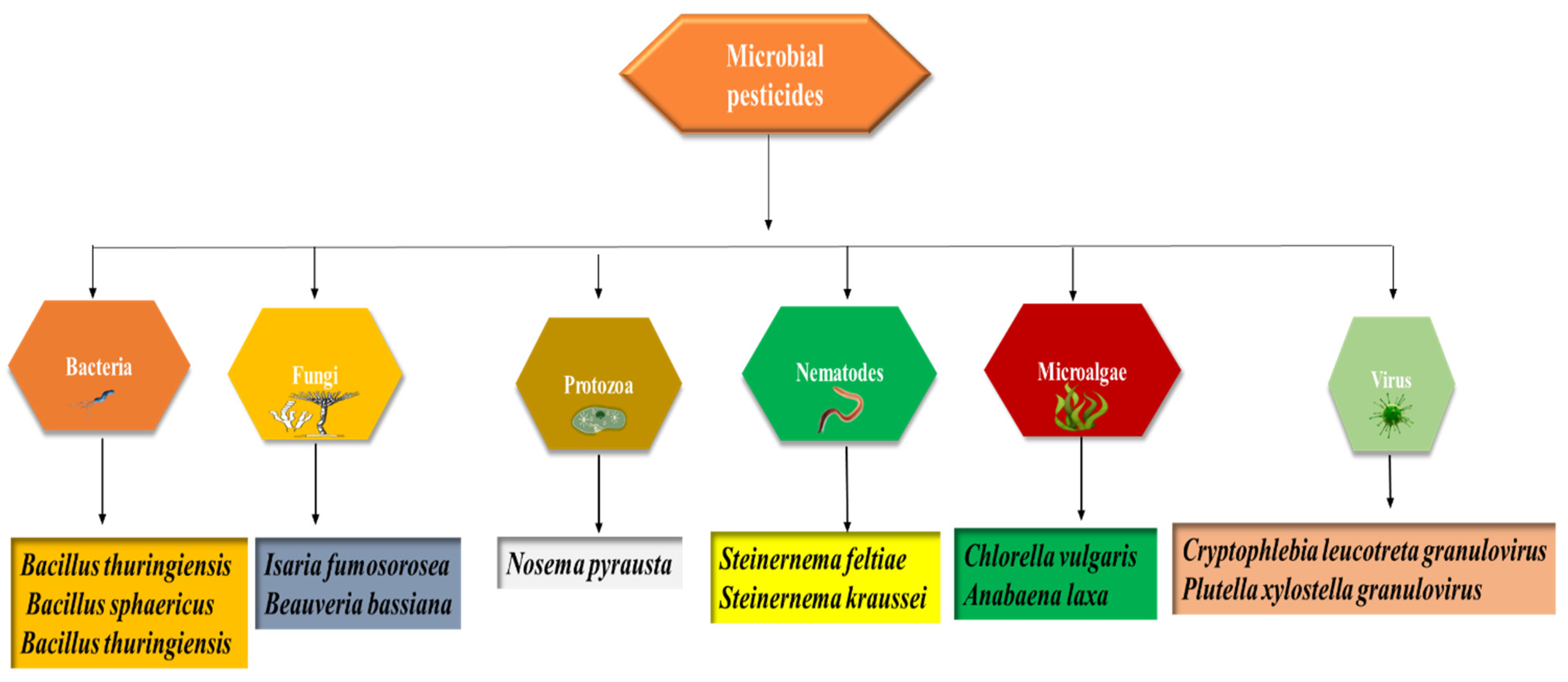
Figure 1. Sources of microbial-based biopesticides.
They are helpful due to their less toxicity, eco-safety, specificity, no development of resistance to pests, and improved crop quality and production [17]. For example, Bt (Bacillus thuringiensis) is one of the biopesticides that is used globally as a tool for insect and pest control. It is also used in the management of phytopathogenic fungi via chitinolytic activity [18]. Kamarulzaman, et al. [19] carried out a comparative study to explore the effectiveness of biopesticides and conventional pesticides in paddy fields. In this study, a neem-based biopesticide (Azadirachta indica), its bioactive secondary metabolite (azadirachtin) bearing insecticidal properties, and conventional pesticides (niclosamideas) were tested. The productivity of rice (Oryza sativa) was increased by the application of biopesticides as compared to the conventional pesticide that offers an alternative for healthier rice cultivation. The primary aim of promoting biopesticides for sustainability is to establish a connection between socially admissible (health, culture, food security), economic growth (farming, marketing, income), and environmental stewardship (water, soil, climate, biodiversity). Along this configuration, the agriculture sector can attain feasibility, community welfare, and eco-safety [20].
3. Diversity of Species of the Genus Bacillus Existing in Agriculture Soil
In the detention of soil, the enormous diversity of microbes, inclusive of bacteria species, archaea, and fungi, are precariously intricate with each other and involved in ecosystem functioning. According to an estimate, 1 g of soil may consist of 1010 to 1011 bacteria, 6000–50,000 bacterial species, and up to 200 m of myco-fungal hyphae, and most of them are propitious for plants and soils [21]. In recent days, intensive farming (by utilizing synthetic fertilizers and chemicals) has been the primary source of food for a growing population. These industrially composed chemical products cause eco-pollution and reduce the microbial population in the soil [22]. To secure biosafety, researchers are involved in the production of nutritious food under sustainable agriculture [23]. The interconnection between the plant, soil, and microbiomes is presented in Figure 2.
Figure 2. Interconnection between the plant, soil, and microbes. Plant roots secrete a prodigious diversity of organic nutrients and signals that attract microbes. Consequently, microbes breakdown the complex nutrients from complex to simple available forms to plants.
Various researches have reported the utilization of biofertilizers instead of hazardous chemicals. The microbes grown on synthetic culture are known as microbial inoculants or biofertilizers. These effective biofertilizers originated from beneficial bioagents that can improve soil fecundity as well as crop yield [23]. Extensive research has reported some probiotic bacteria such as Bacillus, Pseudomonas, Enterobacter, Azotobacter, Serratia, Serratia, Arthrobacter, Erwinia, Microbacterium, Serratia, Azospirillum, Flavobacterium, and Caulobacter [24]. The Bacillus species is one of the significant rhizobacteria species, such as Bacillus subtilis, B. thuringiensis, B. cereus, B. pumilus, etc., that promote plant growth and development and inhibit phytopathogens by the secretion of different exudates such as chitinase and beta-1,3 glucanase, etc. [25].
4. Bacillus spp. as PGPR (Plant Growth Promoting Rhizobacteria)
Bacteria that establish colonies in the rhizosphere (plant root zone) and boost plant growth are referred to as plant growth-promoting rhizobacteria—PGPR [26]. The bacterial strains of the genus Bacillus are among the most well-known PGPR [12]. Bacillus spp. is noted by rhizosphere residents and usually shows growth-promoting activities [27]. Certain Bacillus spp. enhances plant growth either by increasing the absorption of nutrients or by the activation of the host’s defensive mechanism against phytopathogens; in addition, other species can repress the population of pathogenic microbes [28]. These growth-promoting abilities of Bacillus can make it a competent PGPR and beneficent in sustainable agriculture [29]. A research study of Sansinenea [30] reported the inoculation of PGPR induced stress resistance as well as enhanced the yield in numerous crop species like tomato, lettuce, wheat, rice, soybean, groundnut, broad bean, maize, chickpea, etc. Various species, including Bacillus azotofixans, B. subtilis, B. circulans, B. velezensis, B. coagulans, B. macerans, etc., are reported as PGPR [31].
Presently, a change in the world’s climate and deleterious environmental conditions are playing an important role in the reduction of crop development, growth, and yield [32]. The manifestation of new varieties of crops (against stress) by implying genetic engineering and molecular breeding is costly and labor-intensive. So recently, the usage of growth-promoting rhizobacteria has been gaining huge popularity as an alternative approach for amelioration of stress in different crops [33].
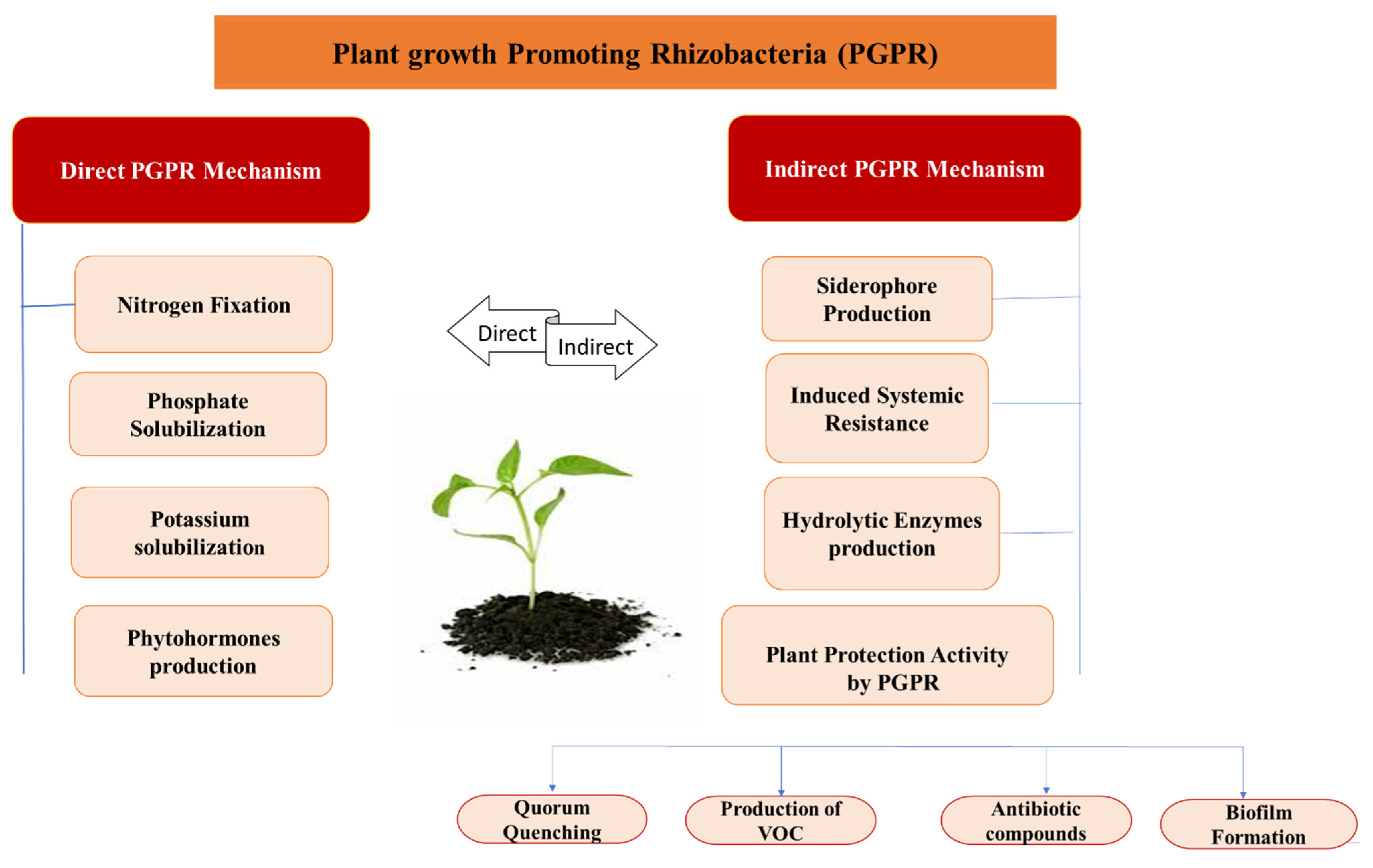
5. Mechanisms of PGPR
Plant growth-promoting bacteria, through various direct and indirect mechanisms, suppress the phytopathogens and promote plant growth, as presented in Figure 3.
Figure 3. Direct and indirect mechanisms of plant growth promoting activities of PGPR.
5.1. Direct Mechanism of PGPR
5.1.1. Nitrogen Fixation
The capability of Bacillus spp. to produce different types of metabolites has a direct impact on plant development and agricultural yield by boosting the nutrients that are available to the plants. Most of the required nutrients are applied to plants by fertilization. This technique not only causes greater economic loss but also produces a harmful impact on the surroundings. Biofertilizers containing phosphorous solubilizing and nitrogen-fixing Bacillus spp. are an acceptable way to minimize dependency on chemical fertilizers while maintaining food safety [34]. Despite being primarily (more than 80%) inaccessible in its atmospheric form, nitrogen (N) is essential for the optimal development of plants [35]. Biological nitrogen fixation (BNF) is a phenomenon in which microorganisms absorb elemental nitrogen from the environment and convert it into a compound that is used by plants as nutrients [36]. Nitrogenase, an enzyme produced by nitrogen-fixing bacteria, catalyzes molecular dinitrogen (N2) to ammonia (NH3) and then is absorbed by the roots of plants. BNF produces approximately 200 million tons of nitrogen annually in the ecosystems of the earth. The microorganisms that are involved in nitrogen fixation may be free-living or symbiotic in nature. By engaging in a-symbiotic nitrogen fixation, several Bacillus spp. can reduce the dependence on nitrogen-based synthetic fertilizers while promoting plant growth along with crop productivity. In crops, 12–70% of total nitrogen uptake is possible due to BNF [35].
5.1.2. Phosphate Solubilization
Phosphorus (P) is a primary macronutrient that is crucial for the development and growth of plants [37]. P-solubilization is defined as the mobility of bound inorganic P (Pi) through the desorption of P and dissolution of P-containing minerals, such as apatite (the group of phosphate minerals) [38]. Alori, Glick and Babalola [37] reported the excessive utilization of synthetic P fertilizers to uplift agricultural production to fulfill ever-increasing global food demand has the potential to pollute surface and groundwater, eutrophication of waterways, and deplete soil fertility. A variety of soil microbes are capable of solubilizing organic P into Pi, which can then be used by plants. These microbes enhance the growth as well as production of a broad range of crops.
5.1.3. Potassium Solubilization
Potassium is the third fundamental plant macronutrient, following nitrogen and phosphorus [39]. This element takes part in several physiological and metabolic processes of the plants, including photosynthesis, stomata regulation, proper seed development, and promoting crop growth and yield [40]. In soils, K-containing minerals that discharge K through weathering are feldspar, muscovite, biotite, alkali, and illite [41]. However, a major dilemma is the unavailability of these minerals (for plants), but with the assistance of K solubilizing microorganisms inclusive of bacteria, actinomycetes, and molds, improved potassium dissolution can be achieved.
5.1.4. Phytohormones Production
Chemical messengers that are mediated in biochemical and physiological processes of higher plants that are active at very low concentrations refer as phytohormones [42]. Charles Darwin was the first person who suggested that certain chemical compounds capable of stimulating growth in crops are latterly known as phytohormones. Microorganisms can stimulate growth and enhance the resistance of plants by synthesizing phytohormones [43]. Plant roots are heavily surrounded by microbes because of root exudates that are rich in nutrient components [44]. Classical bacterial-phytohormones are ethylene, cytokinins, auxins, abscisic acid, and gibberellins [45].
5.2. Indirect Mechanism of PGPR
5.2.1. Siderophore Production by Bacillus spp.
Siderophores are low molecular weight, metal-chelating compounds that are produced under iron-limited conditions by some microbes and plants [46]. Iron (Fe) acts as a key element in various kinds of biological processes, e.g., metabolism of oxygen, synthesis of DNA and RNA, transfer of electrons, and enzymatic processes. Siderophores have the capability to lessen the accessibility of Fe for pathogens [47]. By functioning as biocontrol agents, microbes that create siderophores can restrict the spread of diseases and promote the growth of the plant [11]. Out of various Bacillus spp., B. licheniformis, B. anthracis, B. velezensis, B. thuringiensis, B. cereus, B. halodenitrificans, B. atrophaeus, B. mojavensis, B. pumilus, and B. subtilis are the well-known for siderophore production [48]. Siderophore is induced by numerous species of Bacillus, and these species actively participate in the reduction of different plant diseases. For instance, B. subtilis produced a siderophore, which was involved in the reduction of Fusarium wilt and increased the pepper yield [49].
5.2.2. Induced Systemic Resistance—ISR
Non-pathogenic rhizobacteria have the capacity to lessen diseases in plants by mediating a plant defense process called “Induced Systemic Resistance” (ISR) [47]. It takes a combination of biotic and abiotic stimuli for plants to start developing the ISR (mechanism of resistance). Non-pathogenic rhizobacteria participate in mediating ISR and usually rely on the ethylene (ET) or jasmonate (JA) signaling pathways [50], while Systemic Acquired Resistance (SAR) is promoted through the help of salicylic acid (SA). SAR is responsible for the stimulation of a particular group of defense-related genes, while ISR is not involved in the triggering of any certain kinds of defense-related genes [51]. PGPR induces ISR in plants by releasing various metabolites, e.g., antibiotics, siderophores, volatile organic compounds (VOCs), etc. Through the release of these compounds, PGPR can trigger the mechanism of ISR in plants. Bacillus spp. can initiate ISR by the production of antioxidant defense enzymes. Different defense-related enzymes, e.g., polyphenol oxidase (PPO), superoxide dismutase (SOD), peroxidase (POX), and phenylalanine ammonia-lyase (PAL), are induced by B. subtilis. In tomato seedlings, the prolonged formation of antioxidant defense enzymes induces the mechanism of ISR against early and late blight diseases [52].
5.2.3. Production of Lytic Enzymes
Lytic enzyme production is an intrinsic characteristic of biocontrol agents in the prevention of disease-causing microbes [53]. The activity of lytic enzymes disrupts the cell walls of targeted pathogens by changing the structural stability and integrity [54]. Chitin is a major constituent of the cell walls of fungi, among other composition molecules [55]. Some bacterial strains (PGPR) can degrade fungal cell walls by producing hydrolytic enzymes, including chitinases, dehydrogenases, exo- and endo-polygalacturonases, lipases, phosphatases, proteases, β-glucanases, hydrolases, pectinolyases, and cellulases. Another study reported that the synthesis of lytic enzymes might also be helpful for bacteria to penetrate plant tissues and grow as endophytes [56].
6. Plant Protection Activity Stimulated by Bacillus spp.
Strains of the Bacillus spp. are used as biological control agents (BCAs) to protect plants from pathogenic diseases. Chemical pesticides are being replaced by BCAs, which is a viable option. As a result, various researchers are focusing on exploring their interactions with pests, plants, and pathogenic and beneficial microbes, as well as their environmental impact and human implications. Important characteristics, including efficacy, formulation, stability, and viability, were all thoroughly investigated in many studies.
6.1. Quorum Quenching
Communication inside the bacterial population is feasible with the help of quorum sensing molecules, N-acyl homoserine lactone (AHL). Such indicating molecules are the main reason for boosting the infectious diseases in the pathogenic microbes. Those microorganisms which release AHL lactonase enzyme behave as a biocontrol agent. AHL lactonase is an enzyme that hinders bacterial communication systems by breaking down the quorum-sensing signaling molecule. Quorum quenching was noticed in different Bacillus spp., including B. cereus, B. thuringiensis, and B. licheniformis [57].
6.2. Production of Volatile Organic Compounds (VOCs)
Lower molecular weight lipophilic compounds with high vapor pressure and low boiling point are released by microbial metabolic processes. VOCs function as signal molecules both over short and long distances in the rhizosphere [58]. Additionally, 2,3-butanediol is a volatile organic compound produced by B. subtilis engaged in the mechanisms of plant defense. Phytopathogens were challenged by using the root exudates from peppers inoculated with B. subtilis.
6.3. Antibiotic Compounds
Antibiotic production by beneficial microorganisms [59] is the most effective biological control method for controlling plant diseases. Such chemicals are secreted by Bacillus spp. during sporulation and the stationary development stages [59]. Bacitracin, Kanosamine, fengycin or plipastatin, surfactins, zwittermicin A, kurstakin, gramicidin, and iturins are important antibiotic compounds produced by Bacillus spp. Bacitracin is another kind of antibiotic compound that has strong bactericidal activity. Different Bacillus spp., including B. subtilis and B. licheniformis, have been found to synthesize bacitracin [57].
6.4. Biofilm Formation by Bacillus spp.
In the past, induction of systemic resistance and synthesis of antimicrobial compounds were two reported methods that biocontrol agents utilize to combat phytopathogens. However, current research in the field of biocontrol has focused on biofilm formation and root colonization as defense mechanisms against biocontrol activity. Several Bacillus spp. including B. velezensis, B. atrophaeus, and B. subtilis have been reported to colonize roots and create biofilms as a biocontrol strategy. In many Bacillus species, plant root exudates and various lipopeptides, including bacillomycin and surfactin, play a vital role in the formation of biofilm [60].
7. Multifaceted Role of Bacillus thuringiensis as a Biocontrol Agent
Bacillus thuringiensis (Bt) is an entomopathogenic bacteria that create parasporal crystal proteins (δ-endotoxins). These δ-endotoxins are poisonous to Lepidoptera, Coleoptera, and Diptera, among other insect pests [61]. Throughout the previous century, Bt has been regarded as the most effective bioinsecticide [62]. Because Bt is a rapid-acting and host-specific bioinsecticide, it has few side effects on non-target organisms. Furthermore, its production and use are simple and inexpensive [63]. To generate transgenic crops that are resistant to pests, plant genetic engineering has successfully used Bt as a source of Cry genes [64].
The production of bacteriocins is the main antimicrobial activity of the Bt strain [62]. To strengthen the defense against different microorganisms, prokaryotes frequently produce a variety of antimicrobial peptides. Bacteriocins are tiny, thermotolerant antimicrobial peptides produced by ribosome synthesis in the stationary phase, with molecular weights ranging from 3 to 12 kDa.
Various Bt strains can compete with plant pathogenic bacteria through the production of various bacteriocins and AHL-degrading enzymes. AHL-degrading enzyme (AiiA), released by some Bt strains, can reduce the virulence of pathogenic bacteria like Erwinia carotovora, which causes soft rot in the roots of Capsicum annuum [65]. Furthermore, the inclusion of vegetative cells of Bt in combination with other bacterial (Streptomyces avermitilis and Citrobacter farmeri) and fungal (T. viride, T. parareesei, and Paecilomyces variotii) antagonists significantly increased their effectiveness to suppress Ralstonia solanacearum in Capsicum chinense [66] and S. lycopersicum [67].
Bacillus thuringiensis (Bt) produces crystal proteins (Cry), also called δ-endotoxins. Bacillus produces the most prominent group of insecticidal proteins, which are known as cry toxins. According to the nomenclature committee of Bt toxin, 78 distinct Cry toxins have been identified to date, with Cry1 being the most common [68]. A wide range of B. thuringiensis subspecies produces a variety of Cry toxins. B. thuringiensis var kurstaki produces 31 distinct forms of Cry proteins, the most common of which are Cry1Aa and Cry1Ac. B. thuringiensis israelensis is the main producer of Cry4, Cry10, and Cry11 toxins. Cry1 toxins are mostly active against Dipterans, Lepidopterans, and Coleopterans, whereas Cry2 toxins are mostly poisonous to Dipterans, Lepidopterans, and Hemipterans.
8. Conclusions
Pesticides have been proven to be a promising agent to fulfill the food demand of the growing population. However, these hazardous pesticides have caused human health problems, development of pest resistance, narrowing of biodiversity, and environmental challenges, raising concerns about the pesticides’ safety. Thus, the need to reduce reliance on these synthetic pesticides is pertinent. The application of PGPR is an auspicious solution for eco-friendly agriculture. Bacillus spp. have been elucidated as growth promoters in sustainable agriculture through both direct and indirect mechanisms. The N2-fixation, P and K Solubilization, phytohormones production by Bacillus strains, moreover synthesis of antibiotics, production of lytic enzymes, and ISR are direct and indirect mechanisms, respectively, and all these action mechanisms of Bacillus are supportive in the growth promotion of plants, pest resistance, and circumventing of disease. Some of the Bacillus spp. have been documented as promising biocontrol agents. Food production and its accessibility always are an overwhelming priority to feed the world’s population. So, the best route is to be cautious about chemical-based pesticides. Biopesticides have long been attracting global attraction due to their safer strategy than conventional pesticides. Considering the importance of sustainable agriculture [69][70][71][72], Bacillus spp.-based bioproducts could be a promising addition to sustainable agriculture as there is a limited product range available. There is a dire need to explore the potential of Bacillus spp. in combination with other compatible microbial agents to increase PGP activity and quality food production.
References
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant Pathol. 2011, 60, 2–14.
- Savary, S.; Ficke, A.; Aubertot, J.-N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537.
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111.
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12.
- Souza, R.d.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419.
- Lazarovits, G.; Turnbull, A.; Johnston-Monje, D. Plant Health Management: Biological Control of Plant Pathogens. In Encyclopedia of Agriculture and Food Systems; Elsevier: Amsterdam, The Netherlands, 2014; pp. 388–399.
- Mnif, I.; Ghribi, D. Potential of bacterial derived biopesticides in pest management. Crop Prot. 2015, 77, 52–64.
- Nemutanzhela, M.E.; Roets, Y.; Gardiner, N.; Lalloo, R. The use and benefits of Bacillus based biological agents in aquaculture. Sustain. Aquac. Tech. 2014, 19, 1–34.
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459.
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491.
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500.
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959.
- Aloo, B.N.; Makumba, B.A.; Mbega, E.R. The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 2019, 219, 26–39.
- Kumar, A.; Patel, J.S.; Meena, V.S. Rhizospheric Microbes for Sustainable Agriculture: An Overview. In Role of Rhizospheric Microbes in Soil; Springer: Singapore, 2018; pp. 1–31.
- Kogan, M. Integrated pest management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270.
- Yadouleton, A.W.; Padonou, G.; Asidi, A.; Moiroux, N.; Bio-Banganna, S.; Corbel, V.; N’Guessan, R.; Gbenou, D.; Yacoubou, I.; Gazard, K.; et al. Insecticide resistance status in Anopheles gambiae in southern Benin. Malar J. 2010, 9, 83.
- Dhakal, R. Biopesticides: A Key to Sustainable Agriculture. Int. J. Pure Appl. Biosci. 2019, 7, 391–396.
- Azizoglu, U. Bacillus thuringiensis as a Biofertilizer and Biostimulator: A Mini-Review of the Little-Known Plant Growth-Promoting Properties of Bt. Curr. Microbiol. 2019, 76, 1379–1385.
- Kamarulzaman, P.S.D.; Yusup, S.; Osman, N.; , N.H.; Kueh, B.W.B.; Talib, R. Effectiveness of neem based biopesticide to enhance rice (Oryza sativa) productivity. Sustain. Chem. Pharm. 2018, 7, 36–40.
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Biopesticides in Sustainable Agriculture: Current Status and Future Prospects. In New and Future Development in Biopesticide Research: Biotechnological Exploration; Springer Nature: Singapore, 2022; pp. 1–53.
- Mishra, J.; Prakash, J.; Arora, N.K. Role of Beneficial Soil Microbes in Sustainable Agriculture and Environmental Management. Clim. Chang. Environ. Sustain. 2016, 4, 137.
- Youssef, M.M.A.; Eissa, M.F.M. Biofertilizers and their role in management of plant parasitic nematodes. J. Biotechnol. Pharm. Res. 2014, 5, 1–6.
- Kumar, V.V. Biofertilizers and Biopesticides in Sustainable Agriculture. In Role of Rhizospheric Microbes in Soil; Springer: Singapore, 2018; pp. 377–398.
- Meena, R.K.; Singh, R.K.; Singh, N.P.; Meena, S.K.; Meena, V.S. Isolation of low temperature surviving plant growth–promoting rhizobacteria (PGPR) from pea (Pisum sativum L.) and documentation of their plant growth promoting traits. Biocatal. Agric. Biotechnol. 2015, 4, 806–811.
- Meena, V.S.; Maurya, B.R.; Meena, S.K.; Meena, R.K.; Kumar, A.; Verma, J.P.; Singh, N.P. Can Bacillus Species Enhance Nutrient Availability in Agricultural Soils? In Bacilli and Agrobiotechnology; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 367–395.
- Gray, E.J.; Smith, D.L. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant–bacterium signaling processes. Soil Biol. Biochem. 2005, 37, 395–412.
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586.
- Kumar, A.; Prakash, A.; Johri, B.N. Bacillus as PGPR in Crop Ecosystem. In Bacteria in Agrobiology: Crop Ecosystems; Springer: Berlin/Heidelberg, Germany, 2011; pp. 37–59.
- Govindasamy, V.; Senthilkumar, M.; Magheshwaran, V.; Kumar, U.; Bose, P.; Sharma, V.; Annapurna, K. Bacillus and PaeniBacillus spp.: Potential PGPR for Sustainable Agriculture. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2010; pp. 333–364.
- Sansinenea, E. Bacillus spp.: As plant growth-promoting bacteria. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Springer International Publishing: Midtown Manhattan, NY, USA, 2019; pp. 225–237.
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2019, 128, 1583–1594.
- Lata, C.; Muthamilarasan, M.; Prasad, M. Drought Stress Responses and Signal Transduction in Plants. In Elucidation of Abiotic Stress Signaling in Plants; Springer: New York, NY, USA, 2015; pp. 195–225.
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117.
- Bhattacharyya, P.; Goswami, M.; Bhattacharyya, L. Perspective of beneficial microbes in agriculture under changing climatic scenario: A review. J. Phytol. 2016, 8, 26–41.
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037.
- Mus, F.; Crook, M.B.; Garcia, K.; Garcia Costas, A.; Geddes, B.A.; Kouri, E.D.; Paramasivan, P.; Ryu, M.H.; Oldroyd, G.E.D.; Poole, P.S.; et al. Symbiotic Nitrogen Fixation and the Challenges to Its Extension to Nonlegumes. Appl. Environ. Microbiol. 2016, 82, 3698–3710.
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971.
- Spohn, M.; Zeißig, I.; Brucker, E.; Widdig, M.; Lacher, U.; Aburto, F. Phosphorus solubilization in the rhizosphere in two saprolites with contrasting phosphorus fractions. Geoderma 2020, 366, 114245.
- Sharma, A.; Shankhdhar, D.; Shankhdhar, S.C. Potassium-Solubilizing Microorganisms: Mechanism and Their Role in Potassium Solubilization and Uptake. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 203–219.
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2019, 133, 146–159.
- Raghavendra, M.P.; Chandra Nayaka, S.; Nuthan, B.R. Role of Rhizosphere Microflora in Potassium Solubilization. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 43–59.
- Han, X.; Zeng, H.; Bartocci, P.; Fantozzi, F.; Yan, Y. Phytohormones and Effects on Growth and Metabolites of Microalgae: A Review. Fermentation 2018, 4, 25.
- Sokolova, M.G.; Akimova, G.P.; Vaishlia, O.B. Effect of phytohormones synthesized by rhizosphere bacteria on plants. Prikl. Biokhim. Mikrobiol. 2011, 47, 302–307.
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd Allah, E.F.; Hashem, A. Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress and Fitness. Front. Microbiol. 2017, 8, 2104.
- Patel, K.; Goswami, D.; Dhandhukia, P.; Thakker, J. Techniques to Study Microbial Phytohormones. In Bacterial Metabolites in Sustainable Agroecosystem; Springer International Publishing: Midtown Manhattan, NY, USA, 2015; pp. 1–27.
- Khan, N.; Bano, A.; Zandi, P. Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J. Plant Interact. 2018, 13, 239–247.
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051.
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. Springerplus 2013, 2, 6.
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145.
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants—With special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513.
- Niu, D.; Wang, X.; Wang, Y.; Song, X.; Wang, J.; Guo, J.; Zhao, H. Bacillus cereus AR156 activates PAMP-triggered immunity and induces a systemic acquired resistance through a NPR1 -and SA-dependent signaling pathway. Biochem. Biophys. Res. Commun. 2016, 469, 120–125.
- Chowdappa, P.; Mohan Kumar, S.P.; Jyothi Lakshmi, M.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control 2013, 65, 109–117.
- Xie, J.; Shi, H.; Du, Z.; Wang, T.; Liu, X.; Chen, S. Comparative genomic and functional analysis reveal conservation of plant growth promoting traits in PaeniBacillus polymyxa and its closely related species. Sci. Rep. 2016, 6, 21329.
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering with Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157.
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Rhizosphere Colonization Determinants by Plant Growth-Promoting Rhizobacteria (PGPR). Biology 2021, 10, 475.
- Bodhankar, S.; Grover, M.; Hemanth, S.; Reddy, G.; Rasul, S.; Yadav, S.K.; Desai, S.; Mallappa, M.; Mandapaka, M.; Srinivasarao, C. Maize seed endophytic bacteria: Dominance of antagonistic, lytic enzyme-producing Bacillus spp. 3 Biotech 2017, 7, 232.
- Zamani, M.; Behboudi, K.; Ahmadzadeh, M. Quorum quenching by Bacillus cereus U92: A double-edged sword in biological control of plant diseases. Biocontrol Sci. Technol. 2013, 23, 555–573.
- Fincheira, P.; Quiroz, A. Microbial volatiles as plant growth inducers. Microbiol. Res. 2018, 208, 63–75.
- Alina, S.O.; Constantinscu, F.; Petruţa, C.C. Biodiversity of Bacillus subtilis group and beneficial traits of Bacillus species useful in plant protection. Rom. Biotechnol. Lett. 2015, 20, 10737–10750.
- Pandin, C.; Le Coq, D.; Canette, A.; Aymerich, S.; Briandet, R. Should the biofilm mode of life be taken into consideration for microbial biocontrol agents? Microb. Biotechnol. 2017, 10, 719–734.
- Salehi Jouzani, G.; Pourjan Abad, A.; Seifinejad, A.; Marzban, R.; Kariman, K.; Maleki, B. Distribution and diversity of Dipteran-specific cry and cyt genes in native Bacillus thuringiensis strains obtained from different ecosystems of Iran. J. Ind. Microbiol. Biotechnol. 2007, 35, 83–94.
- Jouzani, G.S.; Valijanian, E.; Sharafi, R. Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711.
- Jain, S.; Vaishnav, A.; Kumari, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. Chitinolytic Bacillus-Mediated Induction of Jasmonic Acid and Defense-Related Proteins in Soybean (Glycine max L. Merrill) Plant Against Rhizoctonia solani and Fusarium oxysporum. J. Plant Growth Regul. 2016, 36, 200–214.
- Melo, A.L.d.A.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of action, resistance, and new applications: A review. Crit. Rev. Biotechnol. 2014, 36, 317–326.
- Park, S.-J.; Park, S.-Y.; Ryu, C.-M.; Park, S.-H.; Lee, J.-K. The role of AiiA, a quorum-quenching enzyme from Bacillus thuringiensis, on the rhizosphere competence. J. Microbiol. Biotechnol. 2008, 18, 1518–1521.
- Bora, L.C.; Kataki, L.; Talukdar, K.; Nath, B.C.; Sarkar, R. Molecular characterizations of microbial antagonists and development of bioformulations for management of bacterial wilt of Naga Chilli (Capsicum chinens Jacq.) in Assam. J. Exp. Biol. Agric. Sci. 2015, 3, 109–122.
- Elsharkawy, M.M.; Nakatani, M.; Nishimura, M.; Arakawa, T.; Shimizu, M.; Hyakumachi, M. Control of tomato bacterial wilt and root-knot diseases by Bacillus thuringiensis CR-371 and Streptomyces avermectinius NBRC14893. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2015, 65, 575–580.
- Crickmore, N. Bacillus thuringiensis Toxin Nomenclature. 2016. Available online: http://www.btnomenclature.Info (accessed on 1 November 2022).
- Jamil, N.; Hyder, S.; Valipour, M.; Yasir, M.; Iqbal, R.; Roy, R.; Zafar, M.U.; Ahmed, A. Evaluation of the Bioremediation Potential of Staphlococcus lentus Inoculations of Plants as a Promising Strategy Used to Attenuate Chromium Toxicity. Sustainability 2022, 14, 13056.
- Fayaz, S.; Kanth, R.H.; Bhat, T.A.; Valipour, M.; Iqbal, R.; Munir, A.; Nazir, A.; Mir, M.S.; Ahanger, S.A.; Al-Ashkar, I.; et al. Leaf Color Chart (LCC)-Based Precision Nitrogen Management for Assessing Phenology, Agrometeorological In-dices and Sustainable Yield of Hybrid Maize Genotypes under Temperate Climate. Agronomy 2022, 12, 2981.
- Mazlan, N.A.; Zaki, N.A.M.; Narashid, R.H.; Talib, N.; Manokaran, J.; Arshad, F.C.; Fauzi, S.S.M.; Dom, N.C.; Valipour, M.; Dambul, R.; et al. COVID-19 Restriction Movement Control Order (MCO) Impacted Emissions of Peninsular Malaysia Using Sentinel-2a and Sentinel-5p Satellite. Earth Syst. Environ. 2022, 1–12.
- Haider, M.W.; Nafees, M.; Valipour, M.; Iqbal, R.; Ali, S.; Asad, H.U.; Faried, H.N.; Aslam, M.; Iqbal, J.; Shahzad, M.A. Postharvest Eucalyptus Leaf Extract Application Extends the Sustainable Supply of Strawberries by Retaining Physicochemical Quality during Cold Storage. Sustainability 2022, 14, 14822.
More
Information
Subjects:
Plant Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.8K
Revisions:
2 times
(View History)
Update Date:
01 Feb 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No