1. Richness of Orchid Taxa and Taxonomic Analysis
The floristic composition of the orchid flora of wetland vegetation in the Central Balkans includes 33 species and subspecies classified in 14 genera. Eight taxa occur exclusively in these vegetation types, while six taxa grow mainly in wetland vegetation types and rarely occur in other vegetation types. On the other hand, there are 11 taxa that grow in wetland vegetation types but occur more frequently and with greater abundance in other vegetation types, while eight taxa very rarely occur in wetland vegetation types. The most taxon-rich genera are Dactylorhiza (ten taxa), Anacamptis (six taxa) and Gymnadenia (four taxa). The genera Orchis and Platanthera are represented by two taxa, while nine genera (Coeloglossum, Epipactis, Herminium, Neotinea, Neottia, Nigritella, Pseudorchis, Spiranthes and Traunsteinera) are represented by a single taxon.
The genus
Dactylorhiza has the highest number of taxa within the total orchid flora of wetland vegetation in the Central Balkans (
Figure 1), which is not surprising considering that wet habitats (fens, bogs, marshes and wet meadows) are typical habitats for representatives of this genus
[1][2]. The occurrence of a large number of
Dactylorhiza taxa can also be explained by the significant presence of silicate geological substrates in the study area, known for their high water-storage capacity, which is favorable for the growth and survival of numerous representatives of this genus
[3]. The presence of the two Balkan endemics (
D. cordigera subsp.
bosniaca and
D. kalopissi subsp.
macedonica) and two Carpathian-Balkan subendemics (
D. cordigera subsp.
cordigera and
D. maculata subsp.
transsilvanica) is particularly important (
Figure 1).
Dactylorhiza × serbica (H.Fleischm.) Soó, a natural hybrid between
D. incarnata subsp.
incarnata and
D. saccifera subsp.
saccifera, which also inhabits the wetlands, was described in Serbia
[4]. In addition,
D. maculata subsp.
maculata,
D. maculata subsp.
trassilvanica,
D. cordigera subsp.
bosniaca and
D. majalis have a southern limit of their distribution in the Central Balkans
[4], making their habitats of high conservation value in this region. The area of the Central Balkans is also a contact zone where
D. fuchsii and
D. maculata subsp.
maculata from the west, north and northwest and
D. saccifera from the south and southeast meet
[5][6], so there is potential for their future taxonomic and phylogeographic research. Due to their complicated taxonomy, the
D. maculata and
D. majalis groups require detailed taxonomic and phylogeographic research in the Central Balkans.
Figure 1. Some representatives of the genus Dactylorhiza of wetland vegetation in the Central Balkans: (a) Dactylorhiza cordigera subsp. cordigera, (b) Dactylorhiza maculata subsp. maculata, (c) Dactylorhiza maculata subsp. transsilvanica, (d) Dactylorhiza incarnata subsp. incarnata, (e) Dactylorhiza saccifera subsp. saccifera, (f) Dactylorhiza sambucina (photos V. Djordjević).
Three
Anacamptis taxa that occur exclusively in wetland vegetation types are
Anacamptis laxiflora,
A. palustris subsp.
palustris and
A. palustris subsp.
elegans (
Figure 2). However, knowledge on the distribution of these taxa in the Central Balkans is insufficient, as these taxa are usually presented in the literature under their species rank for the flora of Serbia, i.e., as
Orchis laxiflora Lam.
[4]. Therefore, published data on the distribution and habitat preferences of these taxa in Serbia cannot be considered with any degree of certainty. Recent studies have shown that
A. palustris subsp.
elegans is the most widespread taxon, while
A. palustris subsp.
palustris is a rarer taxon, distributed mainly in the southern part of the Pannonian plain and very rarely in other parts of the Central Balkans
[4][7][8][9]. Although
A. coriophora subsp.
coriophora,
A. morio subsp.
morio and
A. pyramidalis occur in wet habitats, they are more common in other habitat types (mesophilous and xerophilous grasslands)
[10].
Figure 2. Some representatives of the genus Anacamptis of wetland vegetation in the Central Balkans: (a) Anacamptis palustris subsp. palustris, (b) Anacamptis palustris subsp. elegans, (c) Anacamptis laxiflora ((a) photo I. Stevanoski; (b,c) photo S. Tsiftsis).
Among the
Gymnadenia taxa, the Carpathian-Balkan subendemic
Gymnadenia frivaldii (
Figure 3a), which occurs exclusively in wetland vegetation types, should be emphasized. The center of its distribution is on the mountains of the southeastern Dinaric Alps and on the mountains of the Scardo-Pindhian province, with a disjunction in the southeastern Carpathians
[11][12][13]. This species has the southern and western limits of its distribution in the Central Balkans. Although
G. conopsea is very common in wetland vegetation types in the Central Balkans, where it is often very abundant, this species also grows in other habitats such as mesophilous and xero-mesophilous meadows, as well as open woodlands
[10].
Figure 3. Some representatives of orchids of wetland vegetation in the Central Balkans: (a) Gymnadenia frivaldii, (b) Epipactis palustris, (c) Orchis mascula subsp. speciosa, (d) Platanthera bifolia, (e) Pseudorchis albida, (f) Traunsteinera globosa (photos V. Djordjević).
The orchid flora of the wetland vegetation of the Central Balkans includes a small number of representatives of the genera
Epipactis and
Neottia, known for their typical forest representatives. The species
Epipactis palustris (
Figure 3b), which occurs exclusively in wetland vegetation, is widespread throughout the Central Balkans
[12].
Neottia ovata, on the other hand, is an ecologically very plastic species that grows in wetland vegetation as well as in other vegetation types, including forests
[9][14][15]. The genus
Orchis is also represented by only a few representatives, which is not surprising knowing that the species of this genus tend to grow in xerophilous and mesophilous habitats and often in forest ecosystems. Among the representatives of orchids,
Pseudorchis albida (
Figure 3e) and
Traunsteinera globosa (
Figure 3f) are of great importance, because these species have the southern limit of their distribution in the Central Balkans.
2. Phytogeographical Analysis
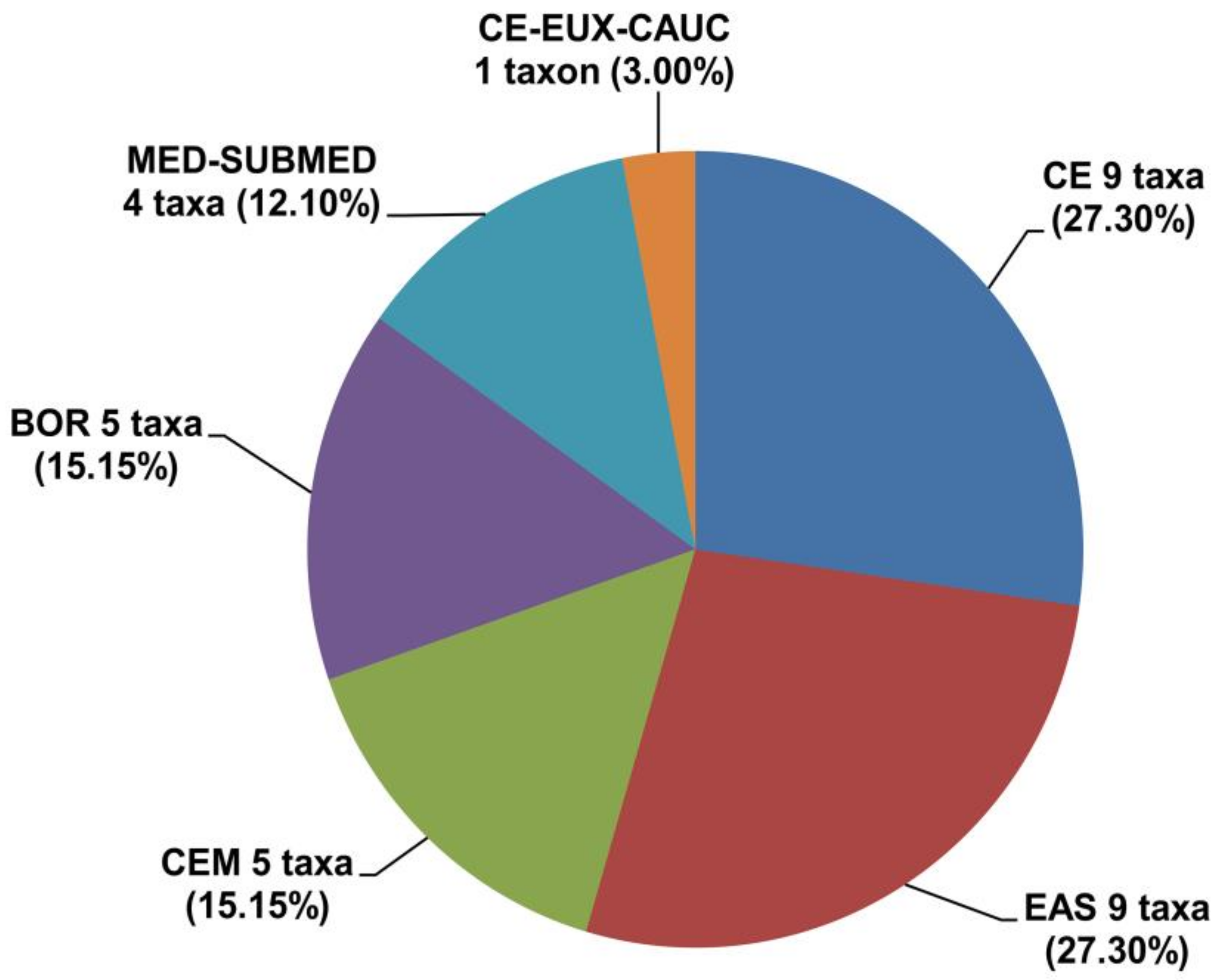
Chorological analysis of the orchid flora of wetland vegetation in the Central Balkans revealed the presence of six chorological groups (
Figure 4). The chorological types for phytogeographical analysis were determined according to the principles defined by Meusel et al.
[16][17], Meusel and Jäger
[18], Stevanović
[19] and Djordjević et al.
[4]. The occurrence of orchids from different chorological groups can be explained by the fact that the Central Balkans is located in an area influenced by different floristic-vegetation regions due to historical, geological, geomorphological and climatic reasons and the considerable altitude differences in the study area.
Figure 4. Spectrum of basic chorological groups of the orchid flora of wetland vegetation in the Central Balkans. CE—Central European; EAS—Eurasian; CEM—Central European mountainous; BOR—Boreal; MED-SUBMED—Mediterranean-Submediterranean; CE-EUX-CAUC—Central European-Euxine-Caucasian.
The chorological analysis of the orchid flora of wetland vegetation in Serbia indicates a pronounced dominance of orchids belonging to the Central European and Eurasian chorological groups (Figure 4). The Central European chorological group includes nine taxa from six genera (Anacamptis, Dactylorhiza, Gymnadenia, Neotinea, Platanthera and Spiranthes). The significant representation of Central European orchids is not surprising, considering that a large part of the Central Balkans has a temperate-continental climate and many different types of habitats where the majority of orchids of this chorological group occur. The Central European mountainous group is represented by five taxa from four genera (Dactylorhiza, Gymnadenia, Nigritella and Traunsteinera). Considering the numerous high-mountain areas in the Central Balkans, orchid representatives from this chorological group are expected.
The Eurasian chorological group includes nine taxa from seven genera (
Anacamptis,
Epipactis,
Gymnadenia,
Herminium,
Neottia,
Orchis and
Platanthera) (
Table 1). Many orchids of this chorological group are characterized by great ecological plasticity, which allows them to grow and survive in different habitats.
Gymnadenia conopsea,
Neottia ovata and
Platanthera bifolia are among the least specialized and most widespread species
[10][14][20].
The boreal chorological group has a significant presence—five orchid taxa from three genera (
Coeloglossum,
Dactylorhiza and
Pseudorchis). Their occurrence in the Central Balkans can be explained not only by historical factors but also by favorable climatic conditions, adequate habitats as well as by the widespread presence of siliceous geological substrates. The fact is that most siliceous substrates, especially acidic and intermediate igneous rocks, have a high water-holding capacity, and siliceous substrates occupy large areas at higher altitudes suitable for many representatives of boreal orchids
[3].
The Mediterranean-Submediterranean chorological group includes four taxa from two genera (
Anacamptis and
Dactylorhiza). While
A. laxiflora is distributed mainly in the Mediterranean and Submediterranean regions and is less common in continental areas,
A. pyramidalis and
D. saccifera are species widely distributed throughout the Central Balkans
[9]. However, the localities of
D. kalopissi subsp.
macedonica and
D. saccifera in the Central Balkans represent their northern distribution limits in this part of Europe. In addition,
D. kalopissi subsp.
macedonica occurs in the Central Balkans only in North Macedonia but is also distributed in Albania, Greece and Bulgaria
[21].
Orchis mascula subsp.
speciosa is the only taxon belonging to the Central European-Euxine-Caucasian chorological group.
3. Life Forms
The orchid representatives of wetland vegetation in the Central Balkans are terrestrial orchids that display characteristics of the geophyte life form
[22][23]. However, orchids were classified according to the concept presented by Tsiftsis et al.
[24], Averyanov
[1], Štípková et al.
[25] and Djordjević et al.
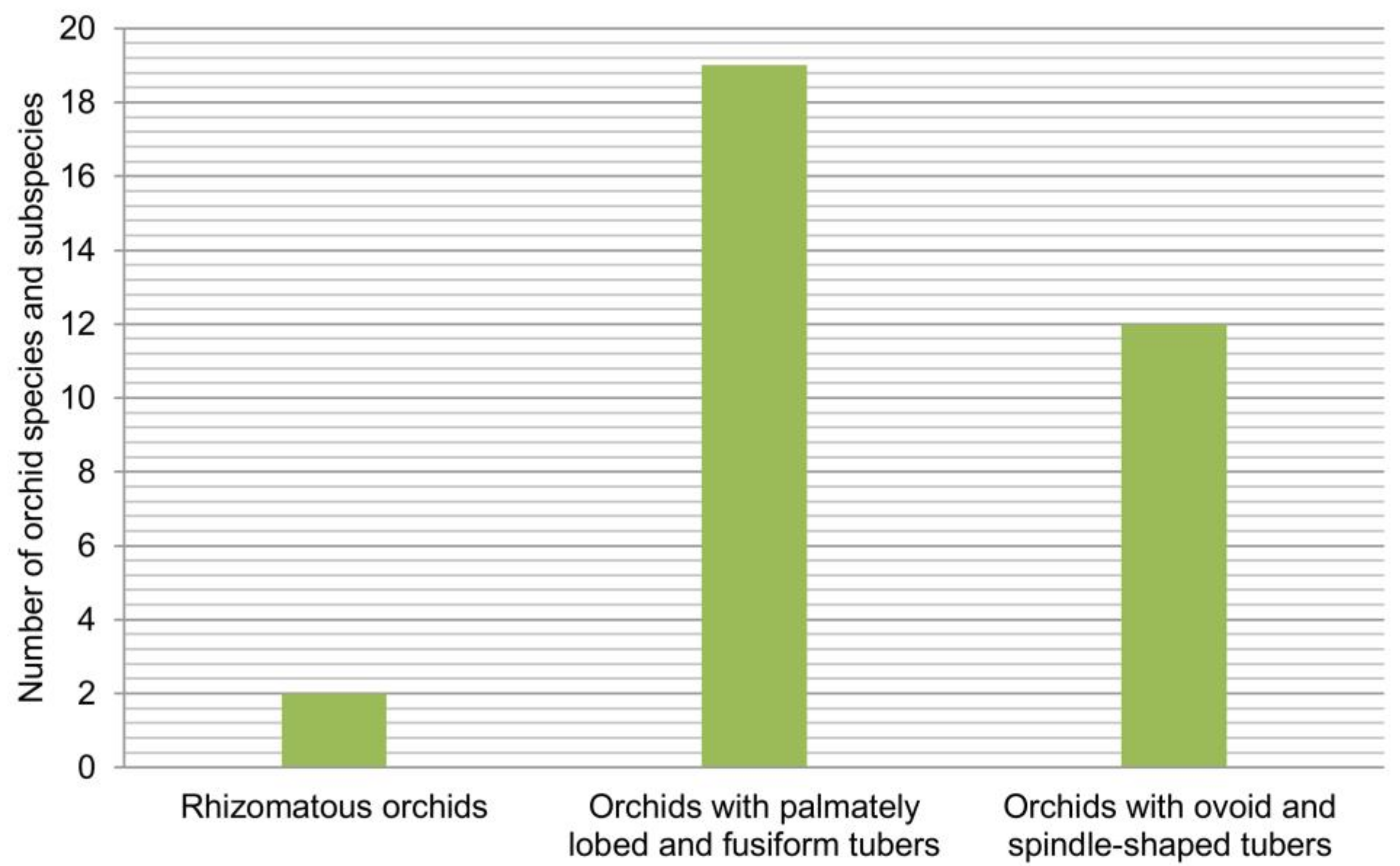
[26]: (1) rhizomatous orchids; (2) “intermediate orchids” (intermediate in evolutionary history between rhizomatous orchids and orchids with spheroid tubers), i.e., orchids with palmate, fusiform, or stoloniferous tubers; and (3) tuberous orchids, i.e., orchids with spheroid tubers. The structure of life forms of orchids of wetland vegetation in the Central Balkans is shown in
Figure 5.
Figure 5. Structure of life forms of orchids of wetland vegetation in the Central Balkans.
The orchid flora of wetland vegetation in the Central Balkans is dominated by "intermediate orchids" (orchids with palmately lobed and fusiform tubers) (
Figure 5). This group includes 19 orchid taxa from six genera (
Coeloglossum,
Dactylorhiza,
Gymnadenia,
Nigritella,
Platanthera and
Pseudorchis). Among these orchids, taxa of the genera
Coeloglossum,
Dactylorhiza,
Nigritella and
Gymnadenia have palmately lobed (finger-like) tubers, whereas species of the genus
Platanthera are characterized by fusiform tubers. The significant presence of these orchids in the wetland vegetation of the Central Balkans is not surprising considering the origin and evolutionary development of orchids of this life form. The first occurrences of “intermediate orchids” have been associated with Alpine orogeny, i.e., the emergence of lower-temperature mountain habitats
[1]. These orchids significantly expanded their range as a result of cooling at the end of the Neogene and in the Pleistocene and were able to colonize areas with plains where the degradation of the Tertiary thermophilic flora took place
[1]. Therefore, they can be considered to have well-developed adaptations to the cold and wet conditions of the habitats.
The group having ovoid and spindle-shaped tubers includes 12 orchid species and subspecies classified into six genera (
Anacamptis,
Herminium,
Neotinea,
Orchis,
Spiranthes and
Traunsteinera). A lower proportion of orchids with spherical tubers in wetland vegetation is to be expected since these orchids usually inhabit dry and semi-dry habitats. Their tubers represent the final stage in the development of the underground organs of orchids, which enable many representatives to survive in habitats with dry and warm conditions
[24]. However,
A. laxiflora,
A. palustris subsp.
palustris and
A. palustris subsp.
elegans are taxa that represent exceptions to the rule and grow exclusively in wetland vegetation types. There are only two orchids with rhizomes (
Epipactis palustris and
Neottia ovata). The smaller number of representatives of rhizomatous orchids is understandable because it is known that these orchids occur mainly in forest ecosystems
[10][14].
4. Wetland Vegetation
Terrestrial orchids are widely represented in various types of wetland vegetation, including wet meadows, as well as bogs, fens and marshes
[10][20][27][28]. In this section, an overview of the main wetland vegetation types with terrestrial orchids and literature sources is presented. A total of 33 orchid species and subspecies were recorded in plant communities from five classes, 10 orders and 17 alliances. The syntaxonomic nomenclature follows Mucina et al.
[29] and Peterka et al.
[30].
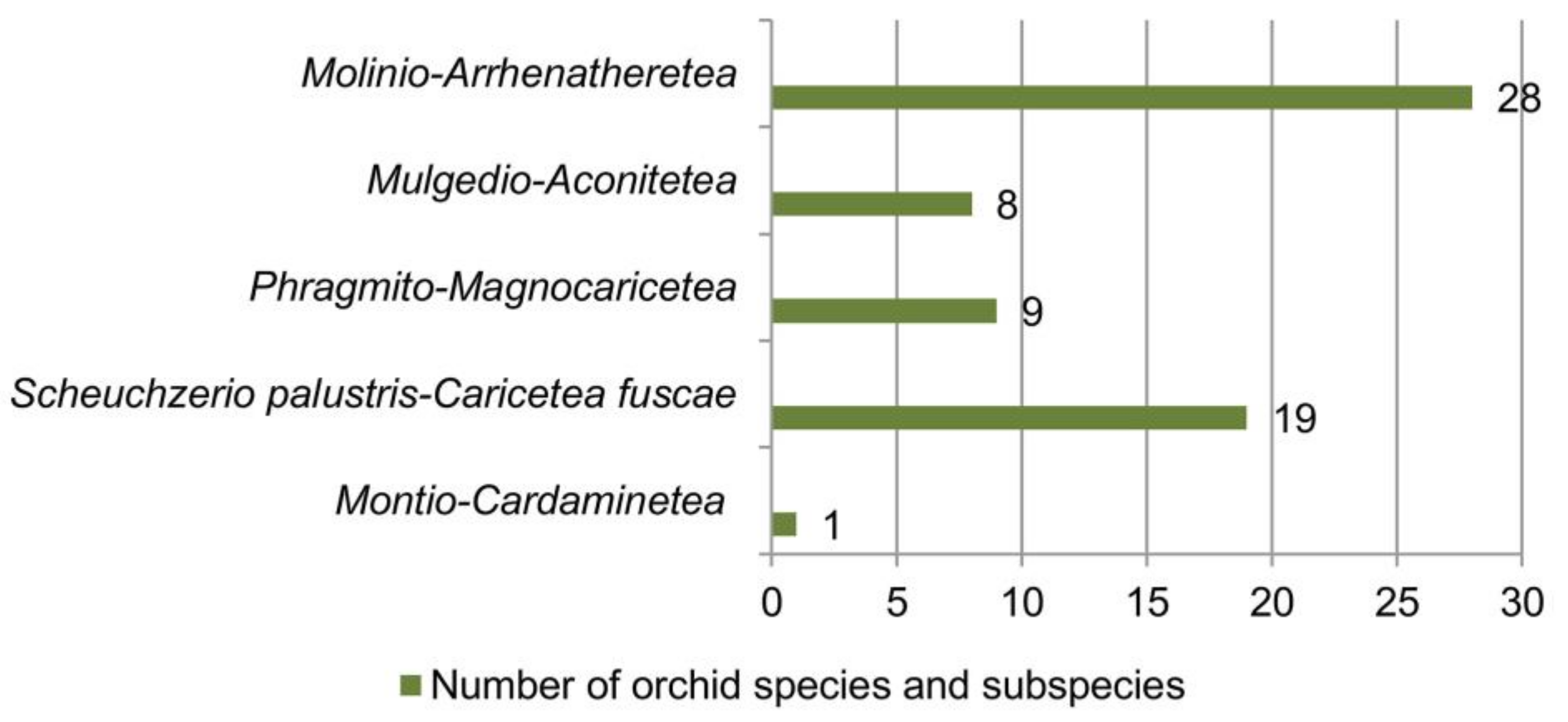
Orchid richness in relation to vegetation classes, orders and alliances in the Central Balkans is presented in Figure 6, Figure 7 and Figure 8. The greatest number of orchids was recorded in the class Molinio-Arrhenatheretea (28 taxa or 84.9% of the total analyzed orchid flora), followed by Scheuchzerio palustris-Caricetea fuscae (19 taxa), Phragmito-Magnocaricetea (nine taxa), Mulgedio-Aconitetea (eight taxa) and Montio-Cardaminetea (one taxon) (Figure 6).
Figure 6. Richness of orchid species and subspecies in relation to vegetation classes.
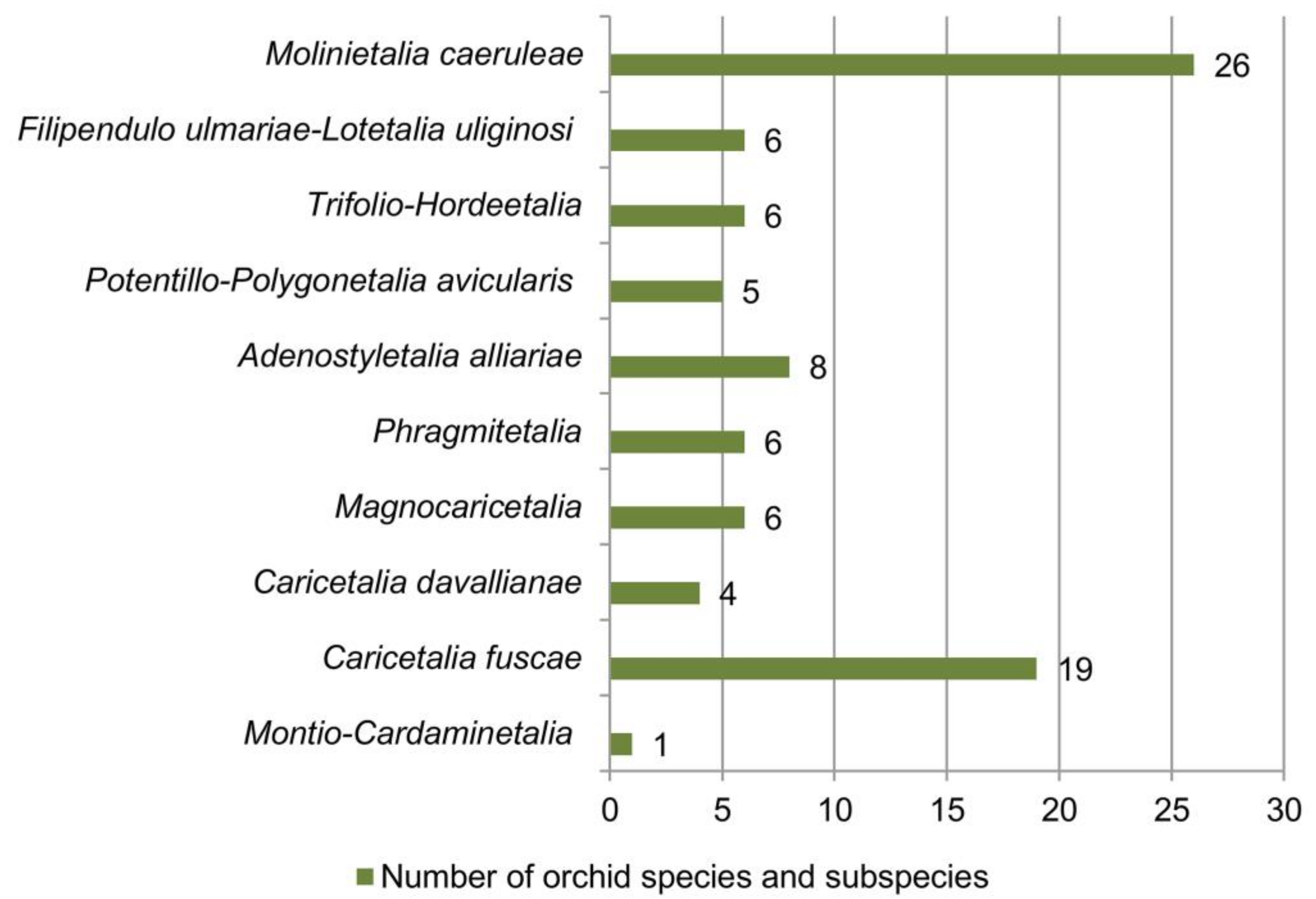
Figure 7. Richness of orchid species and subspecies in relation to vegetation orders.
Figure 8. Richness of orchid species and subspecies in relation to vegetation alliances.
Concerning vegetation orders, the greatest number of orchids was recorded in the Molinietalia caeruleae (26 taxa or 78.8% of the total analyzed orchid flora), followed by Caricetalia fuscae (19 taxa), Adenostyletalia alliariae (eight taxa), Filipendulo ulmariae-Lotetalia uliginosi, Trifolio-Hordeetalia, Phragmitetalia, Magnocaricetalia (six taxa each) and Potentillo-Polygonetalia avicularis (five taxa) (Figure 7). The smallest number of orchid taxa was found in the orders Caricetalia davallianae and Montio-Cardaminetalia (Figure 7).
Regarding the affiliation to vegetation alliances, the greatest number of orchid taxa was recorded in the Molinion caeruleae (24 taxa or 72.7%), followed by Caricion fuscae (19 taxa), Deschampsion cespitosae (15 taxa), Calthion palustris (13 taxa), Cirsion appendiculati (eight taxa), Mentho longifoliae-Juncion inflexi, Phragmition communis (six taxa each), and Trifolion resupinati, Potentillion anserinae, Magnocaricion elatae, Narthecion scardici and Sphagno-Caricion canescentis (five taxa each). The smallest number of orchid taxa was found in the alliances Trifolion pallidi, Molinio-Hordeion secalini, Magnocaricion gracilis, Caricion davallianae and Cardamino-Montion (Figure 8).
4.1. Wet Meadows
Orchids of the Central Balkans, which require hygrophilous and hygro-mesophilous habitat conditions, are particularly abundant in communities of the vegetation class Molinio-Arrhenatheretea. Many orchid taxa, including numerous taxa of the genus Dactylorhiza, were found in communities of the order Molinietalia caeruleae (mown meadows on mineral and peaty soils), especially in the alliances Molinion caeruleae, Deschampsion cespitosae and Calthion palustris.
Recent studies from Serbia have shown that in the case of the alliance
Molinion caeruleae, orchids are most abundant in stands of the communities
Molinietum caeruleae W. Koch 1926 (
Figure 9),
Molinio caeruleae-Deschampsietum cespitosae Pavlović 1951 and
Lathyro pannonici-Molinietum caeruleae Tatić et al. ex Aćić et al. 2013
[9]. Moreover, it has been shown that the composition of orchids in this vegetation type largely depends on the bedrock types. For example,
Molinion caeruleae communities in Serbia on Quaternary sediments and carbonate clastites are particularly suitable for
Dactylorhiza incarnata and
Anacamptis palustris subsp.
elegans, while
Molinion caeruleae communities on serpentine support significant populations of
Platanthera bifolia and
Dactylorhiza maculata subsp.
transsilvanica [20][31]. The importance of the order
Molinietalia caeruleae as an important vegetation type for orchids has also been recognized in other European regions. The following orchid taxa have been recorded in
Molinion caerulae communities in Europe:
Epipactis palustris,
Dactylorhiza majalis,
D. maculata subsp.
maculata,
Neotinea ustulata,
Gymnadenia conopsea,
G. densiflora and
Neottia ovata [15][32][33][34].
Figure 9. The association Molinietum caeruleae W. Koch 1926 (Serbia, photos V. Djordjević).
In the Central Balkans, a significant occurrence of orchids has been noted within the alliance
Calthion palustris, which represents wet grasslands and tall herb communities that are often unmanaged and found on flat lands along streams or on saturated soils near headwaters. Orchids are most frequently recorded in communities of
Equiseto palustris-Eriophoretum latifolii Petković ex Aćić et al. 2013,
Scirpetum sylvatici Ralski 1931 (
Figure 10),
Calthaetum palustris s.l. and
Cirsietum rivularis Nowiński 1927 (
Figure 11). Previous studies in Europe have shown that
Dactylorhiza incarnata,
D. maculata,
D. majalis,
D. praetermissa,
D. saccifera,
D. cordigera,
Epipactis palustris,
Gymnadenia conopsea and
Neottia ovata have significant populations in
Calthion palustris communities
[15][32][33][35][36][37][38].
Figure 10. The association Scirpetum sylvatici Ralski 1931 (Serbia, photo V. Djordjević).
Figure 11. The association Cirsietum rivularis Nowiński 1927 (Serbia, photo V. Djordjević).
In addition, many orchids occur in the Central Balkans in communities of the alliance
Deschampsion cespitosae, which are mown temporarily wet meadows on heavy soils on floodplains in the forest and forest-steppe zones of (sub)continental Central and Eastern Europe. The orchids were most frequently recorded in the Central Balkans within the following communities:
Deschampsietum cespitosae Horvatić 1930,
Agrostio stoloniferae-Juncetum effusi Cincović 1959,
Junco articulati-Deschampsietum cespitosae Petković ex Aćić et al. 2013 and
Rhinantho borbasii-Festucetum pratensis Gajić ex Aćić et al. 2013 (
Table 2). According to earlier published data from Europe,
Dactylorhiza incarnata, D. saccifera,
Epipactis palustris,
Gymnadenia conopsea and
Platanthera bifolia are orchids commonly found in communities of this alliance
[32][33][36].
Some orchids in the Central Balkans have significant representation within the vegetation order
Filipendulo ulmariae-Lotetalia uliginosi (tall-herb wet meadow fringe vegetation on mineral soils). Within this order,
Epipactis palustris and
Dactylorhiza incarnata are among the most common species, especially abundant in
Mentho longifoliae-Juncion inflexi communities
[9][20].
Orchids are less prevalent in communities of the vegetation order
Trifolio-Hordeetalia . This vegetation type represents the wet meadows of the humid continental regions of the north-central Balkans, occurring on clayey, mesotrophic to eutrophic soils on riverside terraces and gentle slopes along the rivers
[29][39][40].
Anacamptis palustris subsp.
elegans is one of the most common taxa that have been recorded both in communities of the alliance
Trifolion resupinati (vegetation of wet meadows of the subarid continental regions of the Southern Balkans) and in communities of the alliance
Trifolion pallidi (vegetation of wet meadows of the humid continental regions of the north-central Balkans).
Anacamptis laxiflora is especially common in communities of the alliances
Trifolion resupinati (
Cynosuro-Caricetum hirtae K. Micevski 1957,
Hordeo-Caricetum distantis K. Micevski 1957 and
Trifolietum nigrescentis-subterranei K. Micevski 1957) and
Molinio-Hordeion secalini [41].
Some orchid taxa in the Central Balkans have been recorded in communities of the order
Potentillo-Polygonetalia avicularis Tx. 1947 and the alliance
Potentillion anserinae Tx. 1947. These are temporarily flooded and heavily grazed nutrient-rich pastures experiencing variable wet-dry or brackish-freshwater alternating conditions of temperate Europe
[29][40][42].
4.2. Tall-Herb Vegetation along Mountain Streams and Springs
Representatives of the family Orchidaceae are less abundant in communities of the vegetation class
Mulgedio-Aconitetea in the Central Balkans. This vegetation represents tall-herb vegetation in nutrient-rich habitats moistened and fertilized by percolating water at high altitudes in Europe, Siberia and Greenland
[29]. Within this vegetation class, certain orchid species were recorded in communities of the order
Adenostyletalia alliariae (tall-herb vegetation on fertile soils at high altitudes of temperate and Mediterranean Europe) and the alliance
Cirsion appendiculati (tall-herb vegetation on acidic soils along mountain streams and springs at high altitudes of the Eastern and Central Balkans).
4.3. Marshland Vegetation
In the Central Balkans, orchids also inhabit marsh communities of the class
Phragmito-Magnocaricetea (reed, sedge bed and herb-land vegetation of freshwater or brackish water bodies and streams of Eurasia). Based on recent studies in the Central Balkans, it can be stated that especially
Dactylorhiza incarnata and
Epipactis palustris are significantly represented in the communities of
Magnocaricion elatae (
Magnocaricetalia) and
Phragmition communis (
Phragmitetalia) (
Figure 12), whereas
Anacamptis palustris subsp.
elegans is recorded in the community of
Magnocaricion gracilis (
Magnocaricetalia). In Germany,
E. palustris has also been recorded in communities of
Magnocaricion elatae (marsh vegetation on oligotrophic to mesotrophic organic sediments of temperate Europe)
[36]. In addition,
Dactylorhiza incarnata,
D. majalis,
Epipactis palustris,
Hammarbya paludosa and
Liparis loeselii were found in the Czech Republic, Hungary and Germany in communities with
Phragmites australis as a strongly represented species
[38][43].
Figure 12. The association Phragmitetum australis Savič 1926 (Serbia, photo V. Djordjević).
4.4. Vegetation of Bogs and Fens
The vegetation class
Scheuchzerio palustris-Caricetea fuscae (fens, transitional mires and bog hollows in the temperate, boreal and Arctic zones of the Northern Hemisphere) represents important vegetation types for many moisture-demanding orchid taxa in the Central Balkans. This vegetation type has been estimated to occupy less than 0.001% of the total Serbian territory
[20], so the existence of 19 orchid taxa in these wetland communities in the Central Balkans indicates its great conservation value. Moreover, recent studies in western Serbia indicated that four orchids (
Dactylorhiza cordigera subsp.
cordigera,
D. maculata subsp.
maculata,
D. saccifera and
Gymnadenia frivaldii) were significantly correlated with this vegetation class
[20]. Orchids in the Central Balkans were recorded in communities of the order
Caricetalia fuscae (sedge-moss vegetation of acidic fens in the boreal and temperate zones and in the supra-Mediterranean belt of mountains in Southern Europe) (
Table 2). Within the order
Caricetalia fuscae, orchids are significantly represented in the following communities:
Carici-Sphagno-Eriophoretum R. Jovanović 1978,
Eriophoro-Caricetum paniculatae R. Jov. 1983 (
Figure 13),
Eriophoro-Caricetum echinatae V. Randjelović 1998 (within the alliance
Caricion fuscae), and
Sphagno-Caricetum nigrae P. Lazarević 2016,
Molinio-Sphagnetum fusci P. Lazarević 2016,
Sphagno-Caricetum rostratae P. Lazarević 2016 (within the alliance
Sphagno-Caricion canescentis).
Figure 13. The association Eriophoro-Caricetum paniculatae R. Jov. 1983 (Serbia, photo V. Djordjević).
Orchids belonging to the alliance
Carici-Nardion V. Ranđelović 1998 at the national level are assigned to the alliance
Caricion fuscae [29][30]. These are wet communities dominated by
Nardus stricta, which are not well defined and for which research is still needed, not only in ecological terms but also in terms of nomenclature and classification. Some of the typical orchid taxa in these communities are
Anacamptis coriophora subsp.
coriophora,
Dactylorhiza sambucina,
D. maculata subsp.
maculata,
D. maculata subsp.
transsilvanica,
D. cordigera subsp.
bosniaca,
D. cordigera subsp.
cordigera,
Gymnadenia conopsea,
Platanthera bifolia and
Traunsteinera globosa. In addition, it should be noted that the separation of the alliances
Sphagno-Caricion canescentis and
Caricion fuscae in the area of the Central Balkans requires additional studies.
The specificity of the Central Balkans is the presence of orchids in the alliance
Narthecion scardici, which represents relic oro-Mediterranean moderately-rich fens of the Balkans. Within this alliance, orchids are significantly represented in the community
Carici-Narthecietum scardici Ht. 1953
[44]. The communities of this alliance have great conservation value, hosting significant populations of
Gymnadenia frivaldii,
Dactylorhiza cordigera subsp.
cordigera,
D. cordigera subsp.
bosniaca and
Pseudorchis albida. Orchids in the Central Balkans are less prevalent in communities of the vegetation alliance
Caricion davallianae (sedge-moss calcareous mineral-rich fen vegetation of Europe and Western Asia) within the order
Caricetalia davallianae.
Communities of the class
Scheuchzerio palustris-Caricetea fuscae are considered important for the growth and survival of numerous orchids in Europe. The following orchids have significant representation within this vegetation class in other European countries:
Anacamptis palustris subsp.
palustris,
Liparis loeselii,
Dactylorhiza cordigera,
D. maculata,
D. majalis,
D. fuchsii,
D. incarnata,
D. lapponica,
D. russowii,
D. traunsteineri,
Epipactis palustris,
Gymnadenia densiflora,
G. frivaldii,
G. conopsea,
Malaxis monophyllos,
Herminium monorchis,
Hammarbya paludosa,
Neottia ovata,
Platanthera bifolia,
Pseudorchis albida,
Spiranthes aestivalis and
S. sinensis [15][29][32][33][35][43][45][46][47][48][49][50].
4.5. Vegetation of Springs
In the Central Balkans, orchids are less common in communities of the vegetation class Montio-Cardaminetea (vegetation of springs of Europe, the European Arctic archipelagos and Greenland). Within this vegetation class, only Dactylorhiza cordigera subsp. cordigera was found in communities of the order Montio-Cardaminetalia and the alliance Cardamino-Montion (vegetation of springs with cold and nutrient-poor water in the subalpine and alpine belts of mountains of central and southwestern Europe).
 +1 credit
+1 credit