Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Adnan Iqbal | -- | 3023 | 2022-12-22 12:04:06 | | | |
| 2 | Amina Yu | Meta information modification | 3023 | 2022-12-23 02:18:07 | | | | |
| 3 | Anna Nadolska-Orczyk | Meta information modification | 3023 | 2022-12-27 11:30:40 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Iqbal, A.; Bocian, J.; Hameed, A.; Orczyk, W.; Nadolska-Orczyk, A. TaNACs in Wheat Crop Improvement. Encyclopedia. Available online: https://encyclopedia.pub/entry/39086 (accessed on 26 July 2026).
Iqbal A, Bocian J, Hameed A, Orczyk W, Nadolska-Orczyk A. TaNACs in Wheat Crop Improvement. Encyclopedia. Available at: https://encyclopedia.pub/entry/39086. Accessed July 26, 2026.
Iqbal, Adnan, Joanna Bocian, Amir Hameed, Waclaw Orczyk, Anna Nadolska-Orczyk. "TaNACs in Wheat Crop Improvement" Encyclopedia, https://encyclopedia.pub/entry/39086 (accessed July 26, 2026).
Iqbal, A., Bocian, J., Hameed, A., Orczyk, W., & Nadolska-Orczyk, A. (2022, December 22). TaNACs in Wheat Crop Improvement. In Encyclopedia. https://encyclopedia.pub/entry/39086
Iqbal, Adnan, et al. "TaNACs in Wheat Crop Improvement." Encyclopedia. Web. 22 December, 2022.
Copy Citation
Wheat (Triticum aestivum) is one of the main staple food crops cultivated around the world. Although it is a very important cereal crop, it is underexplored compared to rice, barley, and maize. The main reason is its large and complex hexaploid genome, AABBDD (2n = 6x = 42), which has three homoeologous sets of genes that make research more demanding. The genome of rice is diploid, 33 times smaller than that of wheat; therefore, this species is very well explored and treated as a model among cereals. Barley and maize are diploid species. However, the first one is more closely related to wheat, thus reports on the cis-regulation of genes in these two species, rice, and barley, by NAC TFs are also discussed and compared with wheat.
wheat
TaNAC
yield-related traits
biotic and abiotic stresses
1. Seed-Associated Traits Regulated by Wheat NACs
Several seed traits, such as seed germination, seed vigor, seed protein content, and endosperm starch content, are associated with wheat yield and quality. Most seed-related traits are regulated by transcription factors. However, considering seed storage proteins (SSPs), very few TFs have been reported. The SSPs in seeds impart elasticity to wheat flour and, due to the different values of this parameter, flour is used for diverse types of food [1][2][3]. The genome-wide study of transcriptional factors that regulate SSP proteins revealed that Triticum urartu TuNAC74 binds to the cis-elements on the promoters of SSP-related genes and increases their activity, while the knockdown of TaNAC74 in wheat reduces SSPs in seeds by 24%. The overexpressed TaNAC74 plants also showed a higher germination rate [2]. T. urartu is an ‘A’ genome donor for bread wheat and having a simple genome is a model for wheat. TuSPR is another gene coding NAC TF from T. urartu. Overexpression of this gene reduced the total content of SSP by 15.97%, while knockdown of its homologue (TaSPR) in bread wheat increased the total content of SSP from 7.07 to 20.34%. Analysis revealed that TuSPR cis-regulated the SSP genes by binding to the 5′-CANNTG-3′ sites distributed in the promoter region [4].
Starch is found in abundance in cereal grains, making it one of the most significant constituents of our diet. The starch content determines the seed yield, quality, and endosperm development. Therefore, it is important to understand the mechanisms of starch content regulation to improve seed yield and quality [5][6]. The quality of wheat flour is also determined by high molecular weight glutenin subunits (HMW-GS), which are important components of SSPs. TaNAC100 overexpression caused a reduced content of HMW-GS, and as a result, total SSPs were also reduced. However, overexpression of this gene increased the expression of two starch synthesis genes, TaGBSS1 and TaSUS2, resulting in a significantly higher seed starch content. Other phenotypic characteristics improved by TaNAC100 overexpression included seed size and thousand seed weight [1].
In the endosperm of cereal grains, starch synthesis starts after sucrose transport. Many enzymes such as sucrose-synthase, ADP-glucose-pyrophosphorylase, phosphoglucose isomerase, and phosphoglucomutase play their role in converting sucrose to ADP-glucose and their transport to amyloplast with the help of the BRITTLE-1 (BT-1) protein, which acts as an ADP-glucose transporter [6][7]. Numerous TFs that regulate the mechanism of starch synthesis have been reported in cereals, including rice bZIP58, NF-YB1, and NF-YC12 [8][9][10]. Furthermore, ZmaNAC36 also known as ZmaNAC130 and ZmaNAC128 from maize has also been reported to play a role in starch biosynthesis [11][12]. Similarly, TaNAC19-A1 in wheat negatively regulated starch biosynthesis in grain endosperm. Overexpression of this gene reduced starch content by binding to 5′-ACGCAG-3′cis-elements of the promoters of TaAGPS1-A1 and TaAGPS1-B1, which were predominantly involved in starch synthesis [13]. The conserved domains of common grain-related NAC TFs may have common cis-elements or binding sites in the promoter regions. However, the difference in negative or positive effects may be due to differences in the C-terminal domain of transcription factors. For example, in wheat, TaNAC019-A1 binds to 5′-ACGCAG/A-3′. A similar cis-element is also present in maize, compatible with two other NAC proteins ZmNAC128 and ZmNAC130, and conversely, in wheat, they positively regulate starch biosynthesis [12][13].
Seed vigor is also a critical trait for crop yield. This trait depends on the timing and uniformity of seed germination. TaNAC2 is a well-established wheat TF that regulates vigor in seeds by binding to the cis-element at the promoter of the TaNRT2.5-3B gene. This gene participates in the acquisition of nitrates from the soil [14]. Two NAC rice genes, namely OsNAC-20 and OsNAC-26, have been reported to regulate seed storage protein and starch [15]. It will be interesting to find the homolog of NAC TFs, already reported from well-established cereal crops, in wheat and perform its functional characterization for crop improvement.
2. NAC-Dependent Root Modulation in Wheat
Root organs are important for the uptake of water and nutrients from the soil, and the importance of the root system increases when the soil is deficient in water. Like any other crop, in wheat, a well-established root system ensures crop yield and quality [16]. Many TFs have been reported to have a significant effect on the plant root system. TaRNAC1 is preferably expressed in wheat roots [17]. Overexpression of this gene resulted in an improvement in root length and aboveground biomass. Furthermore, the transgenic plants showed better tolerance to drought under the applied conditions of PEG treatment. TaRNAC1 has also been reported to interact with GA3-ox2 and enhance its expression in roots. GA3-ox2 encodes an enzyme that converts the inactive form of gibberellin to the active one [17]. Furthermore, overexpression of TaSNAC8-6A led to the activation of various drought-responsive and auxin-signaling genes and subsequently helped to develop lateral roots [18]. Previously, it has been proven that tae-miR-164 modulates many NAC TFs in wheat; however, in a recent study, tae-miR-164 has been reported to target and down-regulate the TaNAC14 gene. Experiments with overexpressed tae-miR-164 and TaNAC14 demonstrated inhibition of root development and reduced tolerance to drought and stress. Therefore, TaNAC14 negatively regulates root development in wheat crops [19].
As a result of global warming, water evaporation poses a serious threat to water reservoirs and causes droughts [20]. Therefore, the productivity of cereal crops will mainly depend on the ability of the roots to extract deep water from the soil. Several recent studies on the plant root system have shown that longer and deeper roots with increased diameter have increased crop yield and quality under reduced water availability conditions [17][21]. However, there are very limited gene and transcription factor data available that indicate the modulation of root systems in cereal crops, including wheat. Therefore, the identification and characterization of more root-regulating TaNACs could be beneficial for wheat improvement. Figure 1 provides a pictorial overview of organ-specific TaNACs.
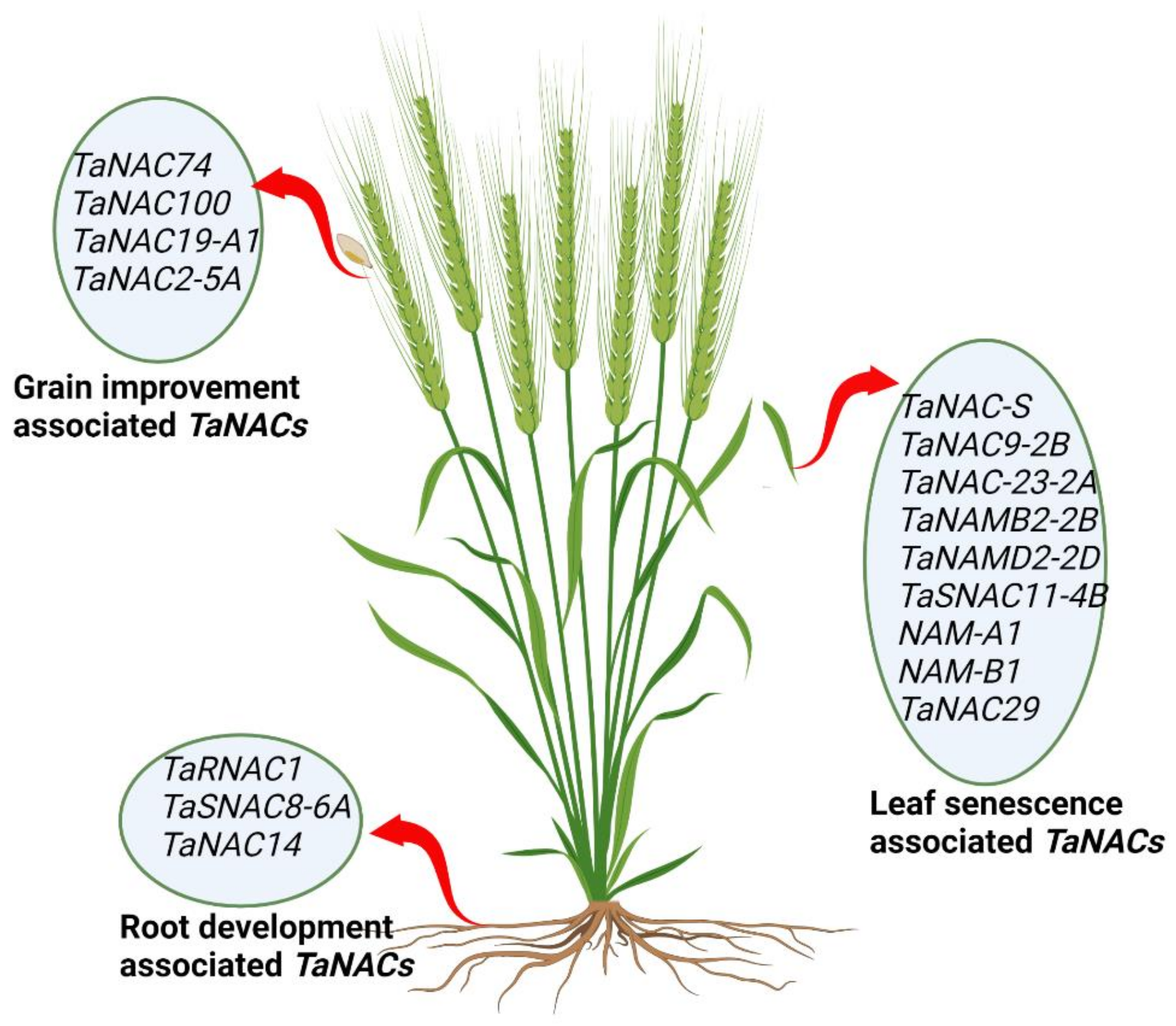
Figure 1. Pictorial representation of TaNACs associated with root development, grain improvement, and leaf senescence in the wheat plant.
3. Role of Wheat NACs in Abiotic Stresses
Biotic and abiotic stresses trigger various changes in plants, from transcription to the level of metabolism, which affect plant growth, development, and yield. TFs interact with functional cis-elements in the promoter regions of stress-related genes and regulate them to achieve better whole-plant tolerance to various biotic and abiotic stresses by either overexpression or underexpression of the genes. Plant NAC proteins are most widely identified as stress-related; however, initially plant NAC proteins were associated with plant development [22][23][24]. Like other crops, wheat also shows a wide range of NAC genes involved in the regulation of biotic and abiotic stress. TaNAC2 is associated with multiple abiotic stresses such as drought, salt, and freezing [25]. Transgenic Arabidopsis was found to be stress tolerant when wheat TaNAC2 was overexpressed [25]. Furthermore, the regulation of TaNAC2L has been reported to be heat-dependent and the expression of this gene has increased significantly in response to higher temperatures. Arabidopsis overexpressing TaNAC2L showed enhanced tolerance to a higher temperature, allowing transgenic plants to be more thermotolerant [26]. Furthermore, TaNAC29 overexpression in Arabidopsis allowed transgenic plants to be more tolerant to drought and salt stresses and also showed a hypersensitive response to ABA [27].
The concentration of hydrogen peroxide (H2O2) is an important indicator of abiotic stress conditions in plants. During abiotic stress, plants have a higher oxidation rate, especially in membrane lipids [28][29]. Treatment with H2O2 in Arabidopsis increased salt and drought-related transcripts of the JUB1 and ATAF1 genes [30][31]. Activities of antioxidant enzymes such as superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) have also increased during salt and drought, and prolonged stress gradually decreased their activities [28][29]. Transgenic plants overexpressing TaNAC29 also showed less accumulation of H2O2 and increased CAT and SOD, leading to increased salt and drought stress. Furthermore, the coexpression analysis indicated that the transcript level of two senescence-associated genes (SAG) SAG13 and SAG113 also decreased [27]. TaNAC47, another wheat NAC gene, is differentially expressed in different tissues of wheat plants in response to the various abiotic stresses. The results suggested that TaNAC47 activated multiple downstream gene expressions. They also uncovered its function as a transcription factor, as it bound to the ABRE cis-element in the yeast one-hybrid assay. In Arabidopsis, TaNAC47 overexpression increased the tolerance of transgenic plants to abiotic stresses such as cold, salt, polyethylene glycol (PEG), and ABA [32].
Drought is one of the devastating abiotic stresses to cereal crops that causes almost 14% of the average losses worldwide. Wheat is also strongly affected by water scarcity; therefore, drought-tolerant germplasm is urgently needed, and this can be achieved with new wheat breeding techniques [18][33]. Many functions of NAC proteins have been validated by overexpression or knockdown experiments to confirm their role in various stresses, including drought stress. An example is TaSNAC8-6A, which belongs to the wheat NAC subfamily. Overexpression of this gene in wheat and Arabidopsis increased tolerance to drought in both transgenic plant lines in an auxin-induced response [18]. Overexpression of rice SNAC1 under the maize ubiquitin promoter leads to improved tolerance to drought and yield in wheat without losing phenotypical traits [18][34][35]. Similarly, transgenic wheat lines overexpressing TaRNAC1 showed tolerance to drought, increased grain weight, and overall biomass under PEG treatment [17]. TaNAC69 is another well-characterized transcription factor for tolerance to drought. Transgenic wheat plants that overexpress TaNAC69 under the drought-specific promoter HvDhn4 exhibited greater tolerance to drought stress under PEG-induced dehydration conditions [36]. Two consensus sequences, expanding to 23–24 bp, were identified for the DNA binding of TaNAC69. The TaNAC69 ortholog was also identified in Arabidopsis [37]. Another NAC gene, TaNAC67, has been associated with various abiotic stresses such as drought, salinity, and freezing. Like many other NACs, TaNAC67 is also ABA-responsive and activates multiple stress-related genes, namely DREB2A, COR15, ABI1, and ABI2. DREB2A. In addition to tolerance to abiotic stresses, transgenic plants also showed a higher chlorophyll content and improved cell membrane strength [38].
Ion flux measurement is a technique that allows for the measurement of salt stress tolerance in plants. NaCl-induced measurement of K+ efflux is a recommended technique to measure salt tolerance in wheat, barley, and Arabidopsis, as represented in Figure 2 [39][40][41]. Transgenic plants overexpressing TaNAC67 showed higher K+ and Na+ efflux rates while there was no significant effect on the H+ ion flux rate [38]. Structural analysis of TaNAC67 indicated that its DNA binding domain had a high similarity index to other NACs such as rice OSNAC1, TaNAC2, TaNAC2A, TaGRAB1 TaNAC4, and TaNAC69, but differs in its C-terminal domain related to transcriptional activity [38]. Stress-related NAC TFs are denoted as SNAC and there are at least 41 stress-related SNAC TFs that fall into 14 different groups by phylogenic analysis. TaSNAC4-3D encodes one of those drought stress-related NAC TFs that negatively regulates drought stress in wheat when induced by ABA, leading to oxidative damage to plant cells [42]. TaSNAC4-3A overexpression in Arabidopsis resulted in increased tolerance to drought by regulating stomatal opening [43].
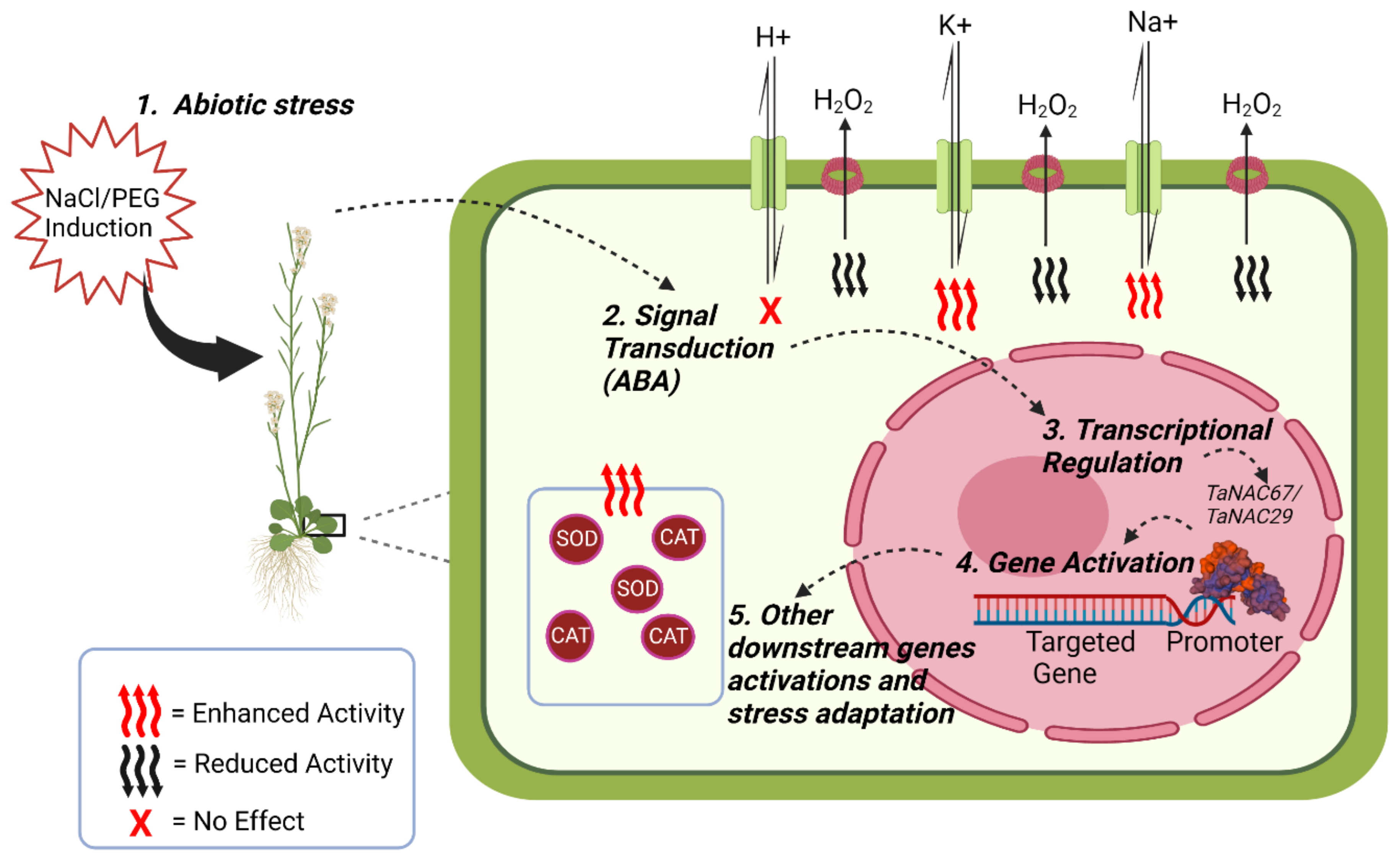
Figure 2. An overview of the regulation of abiotic stress by TaNAC67 and TaNAC29 in transgenic Arabidopsis. (1) Transgenic Arabidopsis overexpressing TaNAC67 and TaNAC29 acquired tolerance to abiotic stress after NaCl and PEG treatment, respectively. (2) NaCl and PEG-induced abiotic stress tolerance leads to transduction of the ABA signal (3 and 4). Transduction of the ABA signal leads to transcriptional regulation by TaNACs to activate their targeted genes. (5) As a consequence of targeted gene activation, some other downstream genes are activated and lead to general stress management. Transgenics overexpressing TaNAC29 accumulated less H2O2 and showed increased CAT and SOD enzyme activities that made transgenics salt and drought tolerant. Similarly, transgenics overexpressing TaNAC67 had higher efflux rates of K+ and Na+ ions, and no significant effect on the H+ ion flux rate allowed transgenics to be more salt-tolerant.
4. Role of Wheat NACs in Biotic Stress
Plants are widely attacked by pathogens including bacteria, viruses, and fungi throughout their life cycle. After a pathogenic attack, plants generally show two types of innate immune response, pathogenic-triggered immunity (PTI) and second, effector-triggered immunity (ETI) [44]. PTI is the first layer of immunity in plants against all microbial attacks, while EFI is a specific immune response that is activated by the interaction between the R protein of plants and the effector proteins of a pathogen [45][46]. The role of NACs in plants is not limited to abiotic stress. Their functional characterization in biotic stress by overexpression or knockdown experiments in Arabidopsis, rice, wheat, and other plants has been performed in a broad way. Numerous NAC proteins have been reported to play a pivotal role in plant immunity by being positive or negative modulators. Many of them modulate immunity by host hypersensitive response (HR) and stomatal regulation of the pathogen [44]. The HR represents one type of programmed cell death (PCD) that prevents the spreading of the pathogen by triggering a primary immune response [47]. OsANC4 is a rice NAC TF that triggers the HR. It was reported that infection of Acidovorax avenae led to enhanced PCD in plants that overexpressed OsNAC4 [48]. Moreover, Arabidopsis overexpressing NAC4 also showed enhanced HR-mediated PCD against pathogenic attack [49].
Stomata not only passively regulate pathogen entry but also play an important role in innate immune responses. Their opening and closing by guard cells represent the first encounter during the plant-pathogen interaction. Three Arabidopsis NACs, ANAC19, ANAC55, and ANAC72, have been reported to regulate stomatal innate immunity by activating signaling pathways (Figure 3) [44][50]. Since Arabidopsis is considered the basic model plant, it will be important to find orthologues of ANAC19, ANAC55, and ANAC72 in wheat to confer stomatal innate immunity.
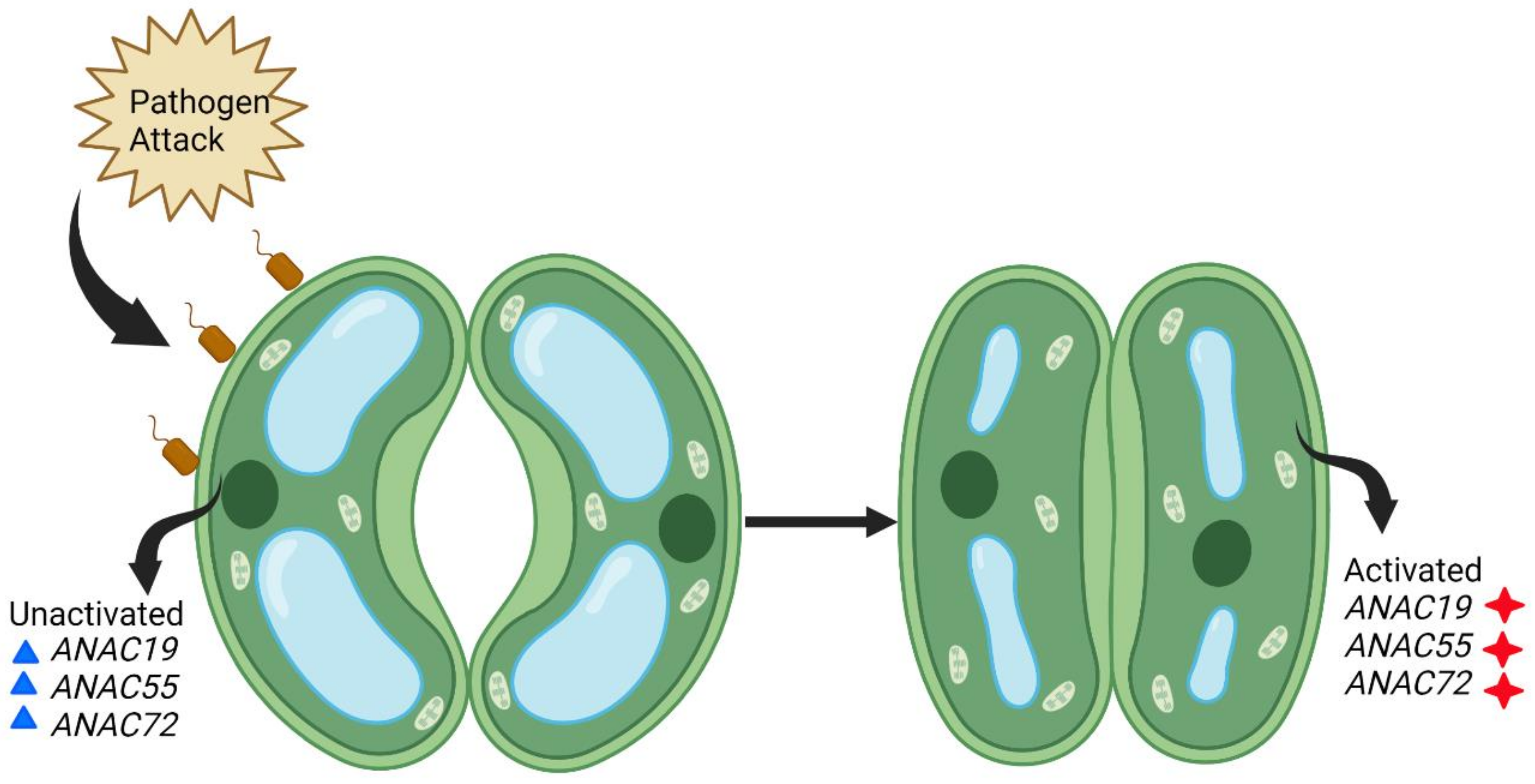
Figure 3. Stomatal innate immunity conferred by three Arabidopsis NACs against biotic stress. Pathogen (brown) attack induces activation of the ANAC19, ANAC55, and ANAC72 genes (inactivated genes are in blue and activated in red) and signal pathways in Arabidopsis and, as a consequence, the stomata confer innate immunity by closing the guard cells.
Like other crops, multiple biotic stresses greatly affect wheat yield around the world. Wheat leaf rust is one of the most serious diseases. At least 186 TaNAC transcripts were obtained from stripe rust and powdery mildew resistant wheat [51]. In a recent study, Zhang et al. [52] reported that TaNAC35 played an important role in the negative regulation of leaf rust resistance against Puccinia triticina. Puccinia striiformis is another fungal pathogen causing stripe rust in wheat. TaNAC30 has been reported to be a negative regulator of resistance to this disease [53]. When wheat plants were infected with P. striiformis, the expression of TaNAC30 increased. On the contrary, silencing of TaNAC30 resulted in increased resistance to rust fungus along with the accumulation of hydrogen peroxide (H2O2) in wheat cells. Similarly, TaNAC2 was also documented to be a negative regulator of resistance to stripe rust in wheat plants. Inhibited expression of TaNAC2 not only reduced P. striiformis (Pst) hyphal growth but also resulted in the accumulation of H2O2, leading to increased resistance to strip rust in an early stage of wheat development [54]. Another NAC, TaNAC8, encodes a protein containing 481 amino acids, and the gene has its orthologue OsNAC8 in rice. TaNAC8 is preferably expressed in seeds rather than in flowers and stems. TaNAC8 expression was induced by the stripe rust-causing fungus, P. striiformis, as well as hormone treatment such as ET and methyl-jasmonate; however, ABA and SA did not induce gene expression. The results of the study indicated that TaNAC8 responded to strip rust infection and some abiotic stresses such as PEG, salinity, and low temperature [55]. Another wheat NAC, TaNAC4, which encodes 308 amino acid proteins, has been reported to have a similar function to TaNAC8. TaNAC4 is a homoeolog of rice OsNAC4 and is preferentially expressed in the roots of wheat seedlings compared to leaves and stems. The expression of this gene is induced by the stripe rust pathogen and by the exogenous application of various hormones, including methyl jasmonate, ET, and ABA; however, there was no notable effect of SA. Furthermore, TaNAC4 has also been reported to regulate various abiotic stresses such as wounding, salinity, and low temperature [56]. Data obtained on TaNAC-responsive genes to stripe rust and powdery mildew offered a great source of information on the functions of NAC in resistance to biotic stress in wheat [51].
The expression of many NAC genes is regulated by microRNAs. It was proved for TaNAC21/22, which was a target gene for tae-miR164 [57]. TaNAC21/22 has been reported to be a negative regulator of resistance to wheat stripe rust disease. Another disease, powdery mildew, is one of the most notorious diseases of wheat crops around the world, and its causative pathogen is Blumeria graminis (Bgt). Three homoeologs of TaNAC6, TaNAC6-A, TaNAC6-B, and TaNAC6-C were characterized for their role in resistance to powdery mildew, and each of the homoeologs responded differently to the Bgt infection. TaNAC6A overexpression increased its resistance to Bgt through the JA pathway. In general, TaNAC6s played a role in resistance to powdery mildew at the base and the broad spectrum levels [58]. Another wheat disease, Fusarium graminearum, is the causative agent of Fusarium Head Blight (FHB) disease. Enhanced resistance to FHB was reported in wheat lines overexpressing TaNACL-D1. This gene encoding NAC-like TF, TaNACL-D1, was found to interact with TaFROG, encoding an orphan protein. Furthermore, TaNACL-D1 was proved to be an orthologue of a Poaceae NAC; however, the C-terminal of TaNACL-D1 is Triticeae specific. Previously, TaFROG was reported to regulate signal pathway activation by interaction with sucrose nonfermenting1 related kinase1 (SnRK1) and improve resistance to Fusarium head blight disease [59]. Biotic stresses pose serious threats to wheat yield and production, and wheat NACs have great potential to cope with these challenges.
References
- Li, J.; Xie, L.; Tian, X.; Liu, S.; Xu, D.; Jin, H.; Song, J.; Dong, Y.; Zhao, D.; Li, G. TaNAC100 acts as an integrator of seed protein and starch synthesis exerting pleiotropic effects on agronomic traits in wheat. Plant J. 2021, 108, 829–840.
- Luo, G.; Shen, L.; Zhao, S.; Li, R.; Song, Y.; Song, S.; Yu, K.; Yang, W.; Li, X.; Sun, J. Genome-wide identification of seed storage protein gene regulators in wheat through coexpression analysis. Plant J. 2021, 108, 1704–1720.
- Shewry, P.R.; Underwood, C.; Wan, Y.; Lovegrove, A.; Bhandari, D.; Toole, G.; Mills, E.C.; Denyer, K.; Mitchell, R.A. Storage product synthesis and accumulation in developing grains of wheat. J. Cereal Sci. 2009, 50, 106–112.
- Shen, L.; Luo, G.; Song, Y.; Xu, J.; Ji, J.; Zhang, C.; Gregová, E.; Yang, W.; Li, X.; Sun, J. A novel NAC family transcription factor SPR suppresses seed storage protein synthesis in wheat. Plant Biotechnol. J. 2021, 19, 992–1007.
- Sonnewald, U.; Kossmann, J. Starches—From current models to genetic engineering. Plant Biotechnol. J. 2013, 11, 223–232.
- Bahaji, A.; Li, J.; Sánchez-López, Á.M.; Baroja-Fernández, E.; Muñoz, F.J.; Ovecka, M.; Almagro, G.; Montero, M.; Ezquer, I.; Etxeberria, E. Starch biosynthesis, its regulation and biotechnological approaches to improve crop yields. Biotechnol. Adv. 2014, 32, 87–106.
- Kumar, R.; Mukherjee, S.; Ayele, B.T. Molecular aspects of sucrose transport and its metabolism to starch during seed development in wheat: A comprehensive review. Biotechnol. Adv. 2018, 36, 954–967.
- Wang, J.-C.; Xu, H.; Zhu, Y.; Liu, Q.-Q.; Cai, X.-L. OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. J. Exp. Bot. 2013, 64, 3453–3466.
- Bai, A.-N.; Lu, X.-D.; Li, D.-Q.; Liu, J.-X.; Liu, C.-M. NF-YB1-regulated expression of sucrose transporters in aleurone facilitates sugar loading to rice endosperm. Cell Res. 2016, 26, 384–388.
- Xiong, Y.; Ren, Y.; Li, W.; Wu, F.; Yang, W.; Huang, X.; Yao, J. NF-YC12 is a key multi-functional regulator of accumulation of seed storage substances in rice. J. Exp. Bot. 2019, 70, 3765–3780.
- Zhang, J.; Chen, J.; Yi, Q.; Hu, Y.; Liu, H.; Liu, Y.; Huang, Y. Novel role of ZmaNAC36 in co-expression of starch synthetic genes in maize endosperm. Plant Mol. Biol. 2014, 84, 359–369.
- Zhang, Z.; Dong, J.; Ji, C.; Wu, Y.; Messing, J. NAC-type transcription factors regulate accumulation of starch and protein in maize seeds. Proc. Natl. Acad. Sci. USA 2019, 116, 11223–11228.
- Liu, Y.; Hou, J.; Wang, X.; Li, T.; Majeed, U.; Hao, C.; Zhang, X. The NAC transcription factor NAC019-A1 is a negative regulator of starch synthesis in wheat developing endosperm. J. Exp. Bot. 2020, 71, 5794–5807.
- Li, W.; He, X.; Chen, Y.; Jing, Y.; Shen, C.; Yang, J.; Teng, W.; Zhao, X.; Hu, W.; Hu, M. A wheat transcription factor positively sets seed vigour by regulating the grain nitrate signal. New Phytol. 2020, 225, 1667–1680.
- Wang, J.; Chen, Z.; Zhang, Q.; Meng, S.; Wei, C. The NAC transcription factors OsNAC20 and OsNAC26 regulate starch and storage protein synthesis. Plant Physiol. 2020, 184, 1775–1791.
- Wasson, A.; Richards, R.; Chatrath, R.; Misra, S.; Prasad, S.S.; Rebetzke, G.; Kirkegaard, J.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498.
- Chen, D.; Chai, S.; McIntyre, C.L.; Xue, G.-P. Overexpression of a predominantly root-expressed NAC transcription factor in wheat roots enhances root length, biomass and drought tolerance. Plant Cell Rep. 2018, 37, 225–237.
- Mao, H.; Li, S.; Wang, Z.; Cheng, X.; Li, F.; Mei, F.; Chen, N.; Kang, Z. Regulatory changes in TaSNAC8-6A are associated with drought tolerance in wheat seedlings. Plant Biotechnol. J. 2020, 18, 1078–1092.
- Qing, C.; Du, L.-y.; Wen, M.; Niu, R.-y.; Wu, B.-w.; Guo, L.-j.; Meng, M.; Liu, X.-l.; Zhao, H.-x. MiR164-TaNAC14 module regulates root development and abiotic-stress tolerance of wheat seedlings. J. Integr. Agric. 2022, in press.
- Long, S.P.; Ort, D.R. More than taking the heat: Crops and global change. Curr. Opin. Plant Biol. 2010, 13, 240–247.
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98.
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248.
- Singh, S.; Koyama, H.; Bhati, K.K.; Alok, A. The biotechnological importance of the plant-specific NAC transcription factor family in crop improvement. J. Plant Res. 2021, 134, 475–495.
- Jensen, M.K.; Skriver, K. NAC transcription factor gene regulatory and protein–protein interaction networks in plant stress responses and senescence. Iubmb Life 2014, 66, 156–166.
- Mao, X.; Zhang, H.; Qian, X.; Li, A.; Zhao, G.; Jing, R. TaNAC2, a NAC-type wheat transcription factor conferring enhanced multiple abiotic stress tolerances in Arabidopsis. J. Exp. Bot. 2012, 63, 2933–2946.
- Guo, W.; Zhang, J.; Zhang, N.; Xin, M.; Peng, H.; Hu, Z.; Ni, Z.; Du, J. The wheat NAC transcription factor TaNAC2L is regulated at the transcriptional and post-translational levels and promotes heat stress tolerance in transgenic Arabidopsis. PLoS ONE 2015, 10, e0135667.
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268.
- Hu, L.; Li, H.; Pang, H.; Fu, J. Responses of antioxidant gene, protein and enzymes to salinity stress in two genotypes of perennial ryegrass (Lolium perenne) differing in salt tolerance. J. Plant Physiol. 2012, 169, 146–156.
- Wang, Q.; Wu, C.; Xie, B.; Liu, Y.; Cui, J.; Chen, G.; Zhang, Y. Model analysing the antioxidant responses of leaves and roots of switchgrass to NaCl-salinity stress. Plant Physiol. Biochem. 2012, 58, 288–296.
- Garapati, P.; Xue, G.-P.; Munné-Bosch, S.; Balazadeh, S. Transcription factor ATAF1 in Arabidopsis promotes senescence by direct regulation of key chloroplast maintenance and senescence transcriptional cascades. Plant Physiol. 2015, 168, 1122–1139.
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.-I.; Asensi-Fabado, M.A.; Munne-Bosch, S.; Antonio, C.; Tohge, T. JUNGBRUNNEN1, a reactive oxygen species–responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 2012, 24, 482–506.
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Jia, J.; Kong, X. The novel wheat transcription factor TaNAC47 enhances multiple abiotic stress tolerances in transgenic plants. Front. Plant Sci. 2016, 6, 1174.
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87.
- Saad, A.S.I.; Li, X.; Li, H.-P.; Huang, T.; Gao, C.-S.; Guo, M.-W.; Cheng, W.; Zhao, G.-Y.; Liao, Y.-C. A rice stress-responsive NAC gene enhances tolerance of transgenic wheat to drought and salt stresses. Plant Sci. 2013, 203, 33–40.
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992.
- Xue, G.-P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant 2011, 4, 697–712.
- Xue, G.-P.; Bower, N.I.; McIntyre, C.L.; Riding, G.A.; Kazan, K.; Shorter, R. TaNAC69 from the NAC superfamily of transcription factors is up-regulated by abiotic stresses in wheat and recognises two consensus DNA-binding sequences. Funct. Plant Biol. 2006, 33, 43–57.
- Mao, X.; Chen, S.; Li, A.; Zhai, C.; Jing, R. Novel NAC transcription factor TaNAC67 confers enhanced multi-abiotic stress tolerances in Arabidopsis. PLoS ONE 2014, 9, e84359.
- Chen, Z.; Newman, I.; Zhou, M.; Mendham, N.; Zhang, G.; Shabala, S. Screening plants for salt tolerance by measuring K+ flux: A case study for barley. Plant Cell Environ. 2005, 28, 1230–1246.
- Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A. Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 2007, 145, 1714–1725.
- Cuin, T.; Zhou, M.; Parsons, D.; Shabala, S. Genetic behaviour of physiological traits conferring cytosolic K+/Na+ homeostasis in wheat. Plant Biol. 2012, 14, 438–446.
- Ma, J.; Zhang, M.; Lv, W.; Tang, X.; Zhao, D.; Wang, L.; Li, C.; Jiang, L. Overexpression of TaSNAC4-3D in Common Wheat (Triticum aestivum L.) Negatively Regulates Drought Tolerance. Front. Plant Sci. 2022, 13, 945272.
- Mei, F.; Chen, B.; Li, F.; Zhang, Y.; Kang, Z.; Wang, X.; Mao, H. Overexpression of the wheat NAC transcription factor TaSNAC4-3A gene confers drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2021, 160, 37–50.
- Yuan, X.; Wang, H.; Cai, J.; Li, D.; Song, F. NAC transcription factors in plant immunity. Phytopathol. Res. 2019, 1, 3.
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539.
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 10.1146.
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256.
- Kaneda, T.; Taga, Y.; Takai, R.; Iwano, M.; Matsui, H.; Takayama, S.; Isogai, A.; Che, F.S. The transcription factor OsNAC4 is a key positive regulator of plant hypersensitive cell death. EMBO J. 2009, 28, 926–936.
- Lee, M.H.; Jeon, H.S.; Kim, H.G.; Park, O.K. An Arabidopsis NAC transcription factor NAC4 promotes pathogen-induced cell death under negative regulation by microRNA164. New Phytol. 2017, 214, 343–360.
- Du, M.; Zhai, Q.; Deng, L.; Li, S.; Li, H.; Yan, L.; Huang, Z.; Wang, B.; Jiang, H.; Huang, T. Closely related NAC transcription factors of tomato differentially regulate stomatal closure and reopening during pathogen attack. Plant Cell 2014, 26, 3167–3184.
- Lv, S.; Guo, H.; Zhang, M.; Wang, Q.; Zhang, H.; Ji, W. Large-scale cloning and comparative analysis of TaNAC genes in response to stripe rust and powdery mildew in wheat (Triticum aestivum L.). Genes 2020, 11, 1073.
- Zhang, N.; Yuan, S.; Zhao, C.; Park, R.F.; Wen, X.; Yang, W.; Liu, D. TaNAC35 acts as a negative regulator for leaf rust resistance in a compatible interaction between common wheat and Puccinia triticina. Mol. Genet. Genom. 2021, 296, 279–287.
- Wang, B.; Wei, J.; Song, N.; Wang, N.; Zhao, J.; Kang, Z. A novel wheat NAC transcription factor, TaNAC30, negatively regulates resistance of wheat to stripe rust. J. Integr. Plant Biol. 2018, 60, 432–443.
- Zhang, X.-m.; Zhang, Q.; Pei, C.-l.; Li, X.; Huang, X.-l.; Chang, C.-y.; Wang, X.-j.; Huang, L.-l.; Kang, Z.-s. TaNAC2 is a negative regulator in the wheat-stripe rust fungus interaction at the early stage. Physiol. Mol. Plant Pathol. 2018, 102, 144–153.
- Xia, N.; Zhang, G.; Sun, Y.-F.; Zhu, L.; Xu, L.-S.; Chen, X.-M.; Liu, B.; Yu, Y.-T.; Wang, X.-J.; Huang, L.-L. TaNAC8, a novel NAC transcription factor gene in wheat, responds to stripe rust pathogen infection and abiotic stresses. Physiol. Mol. Plant Pathol. 2010, 74, 394–402.
- Xia, N.; Zhang, G.; Liu, X.-Y.; Deng, L.; Cai, G.-L.; Zhang, Y.; Wang, X.-J.; Zhao, J.; Huang, L.-L.; Kang, Z.-S. Characterization of a novel wheat NAC transcription factor gene involved in defense response against stripe rust pathogen infection and abiotic stresses. Mol. Biol. Rep. 2010, 37, 3703–3712.
- Feng, H.; Duan, X.; Zhang, Q.; Li, X.; Wang, B.; Huang, L.; Wang, X.; Kang, Z. The target gene of tae-miR164, a novel NAC transcription factor from the NAM subfamily, negatively regulates resistance of wheat to stripe rust. Mol. Plant Pathol. 2014, 15, 284–296.
- Zhou, W.; Qian, C.; Li, R.; Zhou, S.; Zhang, R.; Xiao, J.; Wang, X.; Zhang, S.; Xing, L.; Cao, A. TaNAC6s are involved in the basal and broad-spectrum resistance to powdery mildew in wheat. Plant Sci. 2018, 277, 218–228.
- Perochon, A.; Kahla, A.; Vranić, M.; Jia, J.; Malla, K.B.; Craze, M.; Wallington, E.; Doohan, F.M. A wheat NAC interacts with an orphan protein and enhances resistance to Fusarium head blight disease. Plant Biotechnol. J. 2019, 17, 1892–1904.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
3 times
(View History)
Update Date:
27 Dec 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No