Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ewelina Pogorzelska-Nowicka | -- | 4059 | 2022-07-28 16:14:40 | | | |
| 2 | Sirius Huang | Meta information modification | 4059 | 2022-08-01 04:07:39 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Pogorzelska-Nowicka, E.; Kurek, M.; Hanula, M.; Wierzbicka, A.; Półtorak, A. Formation of Carcinogens in Processed Meat. Encyclopedia. Available online: https://encyclopedia.pub/entry/25621 (accessed on 25 June 2026).
Pogorzelska-Nowicka E, Kurek M, Hanula M, Wierzbicka A, Półtorak A. Formation of Carcinogens in Processed Meat. Encyclopedia. Available at: https://encyclopedia.pub/entry/25621. Accessed June 25, 2026.
Pogorzelska-Nowicka, Ewelina, Marcin Kurek, Monika Hanula, Agnieszka Wierzbicka, Andrzej Półtorak. "Formation of Carcinogens in Processed Meat" Encyclopedia, https://encyclopedia.pub/entry/25621 (accessed June 25, 2026).
Pogorzelska-Nowicka, E., Kurek, M., Hanula, M., Wierzbicka, A., & Półtorak, A. (2022, July 28). Formation of Carcinogens in Processed Meat. In Encyclopedia. https://encyclopedia.pub/entry/25621
Pogorzelska-Nowicka, Ewelina, et al. "Formation of Carcinogens in Processed Meat." Encyclopedia. Web. 28 July, 2022.
Copy Citation
Meat is a rich source of various nutrients. However, it needs processing before consumption, what in turn generates formation of carcinogenic compounds, i.a., polycyclic aromatic hydrocarbons (PAH), nitrosamines (NOCs), and the most mutagenic heterocyclic aromatic amines (HAAs). It was widely found that many factors affect the content of carcinogens in processed meat.
meat
carcinogens
heterocyclic aromatic amines (HAAs)
artificial digestion
1. Thermally Processed Meat Is a Source of Carcinogenic Compounds
Heterocyclic aromatic amines are a group of compounds with a structure consisting of 2–5 aromatic cycles, 1 or more nitrogen atoms, and 1 exocyclic amino group (with the exception of harman, norharman, and Lys-P1) [1]. HAAs are formed in products with high protein content during heat treatment exceeding a temperature of 120 °C. The main substrates for HAAs formation are free amino acids found in muscle tissue, especially creatine and creatinine. Taking into account that HAAs are produced during Maillard reactions, the degree of their formation also depends on the pH of the meat. Over 25 HAAs were isolated and identified in thermally processed food in the past 40 plus years. HAAs might be divided into two large groups in terms of their polarity. Polar HAAs (amino-imidazo-azarens, IQ group) are formed during food processing in high temperatures of 100–300 °C (smoking, frying, toasting, grilling). Pyroindols and pyro imidazoles are non-polar HAAs which form also during heat treatment of food but at temperatures above 300 °C; therefore, they are called pyrolytic amines. HAAs are probably the most mutagenic and carcinogenic compounds generated in heat-treated food products. It has been reported that they are 2000-fold more mutagenic in comparison to benzo[a]pyrene and 100-fold stronger than aflatoxin [2][3]. Nine heterocyclic aromatic amines (including MeIQ, MeIQx, PhIP, AαC, MeAαC, Glu-P-1, Glu-P-2, Trp-P-1, Trp-P-2) have been recognized by the IARC as possible human carcinogens (group 2B) and one (2-amino-imidazo [4,5-ƒ]quinoline) as a probable carcinogen (2A group). It has been proved that HAAs can induce tumors in rodents and primates (nonhuman) at several locations such as: the lungs, liver, mammary glands, prostate, and colorectum [4]. In the United States, the National Toxicology Program listed four HAAs (PhIP, IQ, MeIQ, 8-MeIQ) as carcinogenic to humans.
Polycyclic aromatic hydrocarbons are pollutants containing two or more fused aromatic rings. They are formed as a result of incomplete combustion of organic matter. PAHs include many harmful components of carcinogenic, mutagenic, and teratogenic nature with benzo(a)pyrene being the most predominant [5]. They are commonly spread in nature (so far 100 compounds are recognized). However, humans are exposed to them mainly through food (>90% of exposure). Cooking methods such as grilling, roasting, smoking, and barbecuing generates PAHs formation. Even though PAHs can be formed also from proteins and carbohydrates, the rapid increase in its content is observed during the fat pyrolysis at temperatures above 200 °C. Many factors affect PAHs formation in cooked meat, such as: temperature, cooking time, fat content, type of fuel used for cooking, and the occurrence of melted fat dropping onto the heat medium. The International Agency for Research on Cancer of the World Health Organization classified three PAHs (benz[a]anthracene, benzo[a]pyrene, dibenz[a,h]anthracene) as probably carcinogenic for humans (2A group) and three (benzo[a]fluoranthene, benzo[k]fluoranthene, indeno[1,2,3-cd]pyrene as possibly carcinogenic to humans (2B group). It is worth noting that according to the EFSA the acceptable level for benzo[a]pyrene is 2 μg/kg product, while the sum of the four PAHs bezno(a) pyrene (BaP), chrysene (CHR), benzo(a)anthracene (BaA), and bezno(b)fluoranthene (BbF) must not exceed 12 μg/kg [6].
In terms of NOCs, N-nitrosodimethylamine (NDMA) is the most carcinogenic, as a dose of 10 µg/kg body weight is sufficient for mouse feed. This is a component that directly depends on the nitrogen concentration in the meat; hence, the amount is significantly higher in cured foods. For example, in the shoulder it is of about 2.46 µg/kg NDMA, while for the cured shoulder it is over 9 µg/kg. The highest NDMA values are observed in smoked products ranging between 10 and 17 µg/kg, while the lowest values of around 1–2 µg are observed for pasteurized canned products [7]. NDMA is also observed in products other than beef and pork. Very high amounts are found in salted fish—12.64–322.92 µg/kg, which is a very big difference compared to fresh fish, where the content of this substance ranges from 0.04 to 3.5 µg/kg. Quite low NDMA values are observed in poultry, as chicken contains about 0.52, while duck meat contains 0.13 µg/kg [8]. The content of NOCs is directly dependent on the heat treatment method used (smoking, steaming) and the curing process.
It is also worth mentioning that ingredients typically found in meat products are not the only ones responsible for the formation of carcinogenic compounds. Substances that are air pollutants, such as toxic trace elements, polychlorinated dibenzo-p-dioxins and dibenzofurans, polychlorinated biphenyls, polybrominated diphenyl ethers, polychlorinated diphenyl ethers, polychlorinated naphthalenes, and perfluoroalkyl substances can also constitute a substrate for the formation of carcinogenic compounds during heat treatment [9].
2. Parameters Affecting HAAs Content in Cooked Meat
2.1. Cooking Parameters Affect Content of Carcinogenic Compounds in Meat
Thermal conditions are crucial factors which directly affect HAAs and PAHs content in meat [10][11]. The speed of HAAs and PAHs formation accelerates with the temperature increase [12]. Steaming, boiling in water, or heating in a microwave, are cooking methods that operate at low temperatures usually not exceeding 100 °C. HAAs are not detected or detected at scarce amounts in meat prepared under such conditions [13][14]. It has been established that mutagenic substances start to form at 125 °C [15]. Further, the dynamics of HAA formation increase significantly at 150 °C, while the highest increase in the content of these mutagenic compounds is observed at 200 °C. In pork fried at 125 °C, 58 ng/g of HAAs was detected while at 150 °C almost two-fold more (91 ng/g of HAAs). Similarly for beef meat, the amount of IQx, MeIQx, PhIP, and 4,8-DiMeIQx generated during thermal processing increased 2.5, 6, and 8 times, respectively, when the temperature rose from 170 to 200 °C [16]. In turn, PAHs start to form at 200 °C and the highest amount is observed after the temperature rises to 500–700 °C. The type of cooking method is the next important factor determining mutagenic activity of heat-treated meat [17][18]. Different cooking methods have different effects on the content of carcinogenic compounds. For instance, grilled drake meat is characterized by the lowest HAAs concentration in comparison to pan-fried, roasted, and deep-fried meat [19]. The source of energy also matters. Chicken grilled with charcoal had a higher content of PAHs than oven-grilled chicken [20]. Generally, it was established that more gentle cooking methods (operating at lower temperatures such as boiling and roasting) generate less mutagenic compounds [1]. Analogously, cooking methods operating at high temperatures (pan frying, grilling, oven roasting) lead to formation of mutagens at higher levels [1]. This trend was observed in various studies. However, temperature itself is not only factor that determines HAAs content in meat. An example of that was found in a study where stone-barbecued beef steak was characterized by a higher amount of HAAs in comparison to wire-barbecued beef steak. The results suggested that there are other important factors such as the type of heat transfer (conduction, convection, radiation) and occurrence of surrounding media (water, fat, metal, air) which changes heat transfer coefficients [21]. The coefficient is better in methods with direct contact. For pan frying, the coefficient is equal to 150 Wm2/K in comparison to 30–40 Wm2/K for air convection. That is why the meat prepared on a contact grill had 10 times higher MeIQx content than meat prepared using a convection oven and deep-fat fryer [22]. In the case of oven roasting, heat is transferred indirectly by convection and similarly for microwave heating where heat is transferred indirectly by radiation. Thus, the meat cooked in a microwave contains much fewer carcinogenic compounds in comparison to deep-fried or charcoal-grilled beef and chicken [23]. In the case of direct heating, for instance by pan frying, there is a possibility to reduce HAAs formation by turning the meat in the pan, frequently. It is also extremely important not to let the melted fat fall onto the heat source. The direct contact of fat with the flame results in the formation of a huge amount of PAHs. PAHs content is influenced by both source of the product and the method of thermal processing. However, the latter is of greater importance. For example, benzo[a]pyrene content was revealed to be 0.7 µg/kg for smoked pork meat and 1.3 µg/kg for chicken meat. In the case of sausages subjected to three processing methods, smoked sausage contained 1.1 µg/kg of benzo[a]pyrene, grilled sausage 0.5 µg/kg, and cooked sausage 0.15 µg/kg [24]. However, those factors mentioned above (temperature and cooking method) occur simultaneously and thus it is important to verify their additive impact on the final content of HAAs.
Time also exerts a significant impact on the formation of carcinogenic compounds. Measuring the aqueous model system of meat extracts, it was noted that the content of 4,8-DiMeIQx after heating (175 °C) for one hour was 1.2 ng/g, when the time was doubled (at the same temperature) the amount of 4,8-DiMeIQx increased three times [25]. The same trend was observed in a study analyzing the HAAs content in ground beef fried on the pen at 150–180 °C. The small level of those substances was found after 3 min of the process; however, the HAAs amount increased with time passage [26]. In the case of beef there are studies documenting that the HAAs do not form in less than five minutes of heat treatment, and certain mutagens (IQ, MeIQ, 4,8-DiMeIQx, AαC, and harman) were detected in a small amount in no less than 10 minutes. However, the impact of time on the HAAs formation is directly linked with temperature of the process (Figure 1). In, turn, PAHs formation is most pronounced during the first few minutes of cooking. It is probably caused by its deposition on the meat surface what cuts off the fat (precursor) from the heat medium and in turn further formation of PAHs is not possible [27].
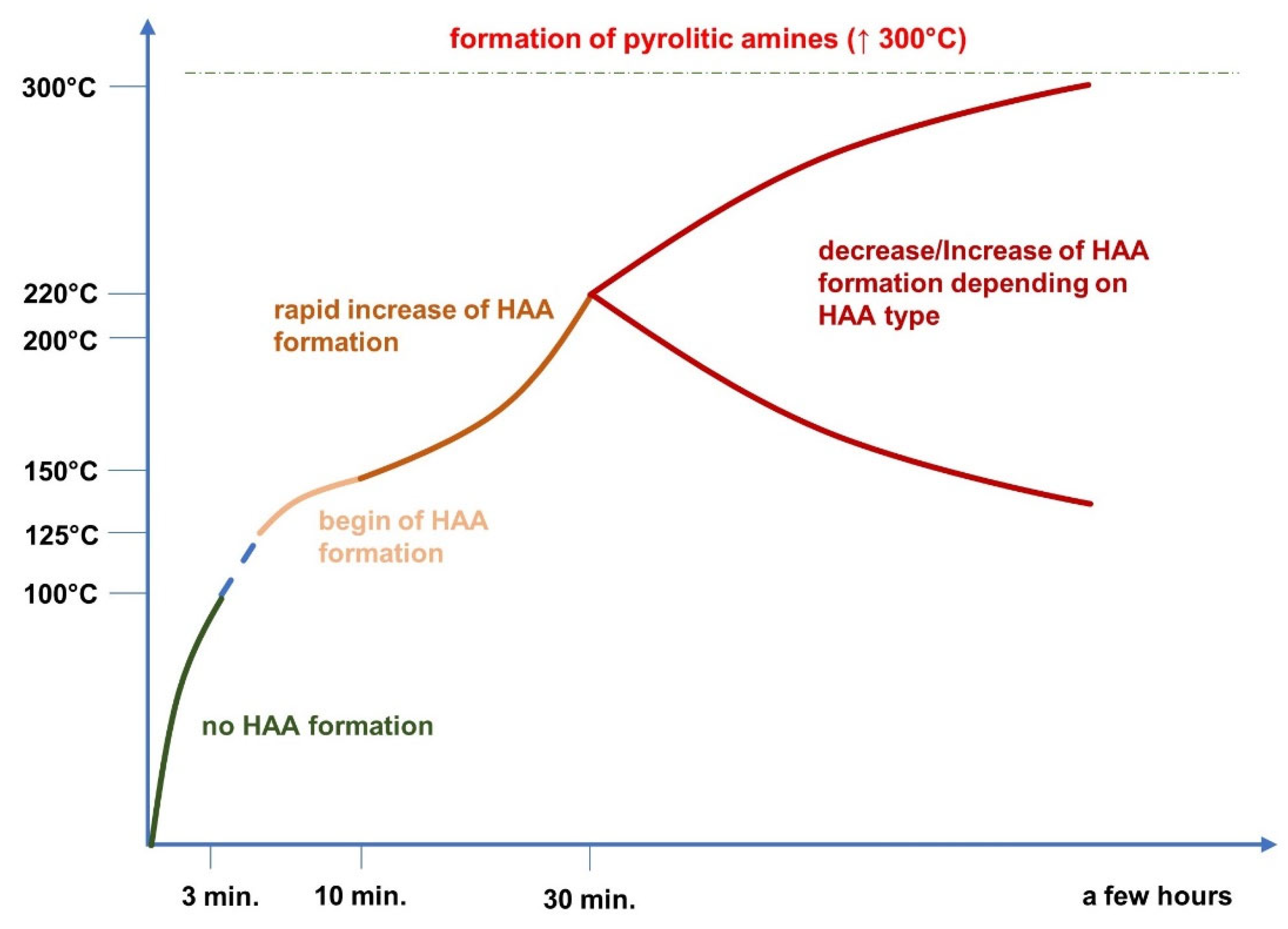
Figure 1. Changes in the content of HAAs depending on applied temperature and time.
Generation of HAAs would be impossible without precursors: creatine, carbohydrates, amino acids, nitrogenous bases, and nucleosides. Creatine under heat treatment converts into the physiologically inert product creatinine. In turn, creatinine is essential during formation of the imidazole ring. Thus, formation of some HAAs such as IQ and IQx is completely dependent on creatinine content in meat [28]. This process is graphically depicted in an article by Cao et al. [29]. It was proven that beef containing a low level of creatinine was less mutagenic than beef with a high creatinine content [30]. However, formation of non-polar HAAs is not affected by creatinine content. Thus, other amino acids are necessary to create HAAs as well. Different amino acids can be precursors for the formation of the same mutagenic compounds, e.g., glycine, serine, threonine, lysine, and alanine, contribute to the formation of MeIQx [31]. Equally important precursors for HAAs formation are reducing sugars (glucose, fructose, lactose, ribose). However, the key is the ratio between the content of amino acids and sugars. When the content of sugar is higher than in the natural state (half the molar quantity of sugar to the amount of amino acids) the HAAs generation is reduced [32]. Thus, there was a 50% decrease in HAAs content observed in meat obtained from pigs with the RN allele (higher glycogen concentration) in comparison to meat acquired from normal pigs [33]. In general, the content of HAAs varies significantly depending on the meat type what is caused by the differences in the content of precursors. For instance, in ground beef, 50 ng/g of PhIP was detected [34] and 2 ng/g of MeIQx. Slightly lower values for PhIP were obtained when meat was fried at 200–230 °C reaching the level of 10–25 ng/g. Interestingly, significantly higher values are observed for poultry meat. For instance, in barbecued breast fillets the PhIP content reached 480 ng/g when in oven-broiled meat only 72 ng/g of PhIP was measured [35]. The lowest HAAs values were observed for pork. Grilled pork steaks contained about 28.62 ng/g of PhIP [36].
The fat content and its oxidative status was proven to have an impact on the formation of mutagenic substances as well. The more efficient ability of fat to transfer heat enables to reach higher temperatures in a relatively short time, what increases generation of HAAs and PAHs in meat. Furthermore, higher fat content reduces the proportion of other precursors needed for HAAs formation such as creatine, sugars, and amino acids. It has been proved that increased level of fat above the optimal ratio decreases HAAs content [37]. Knize et al. [38] have reported that ground beef containing 30% of fat was less mutagenic (150,000 revertants per kg of fresh beef) than beef containing 15% of fat (230,000 revertants/kg). Moreover, the oxidative status of fat is equally important. The oxidized fat increases the level of carcinogenic compounds in tested samples [39].
2.2. Bioactive Compounds Are Effective Inhibitors of Carcinogen Formation in Heat-Treated Meat
An effective method to reduce the formation of HAAs is to enrich meat with substances of antioxidant properties because radicals may take part in the mechanism of HAAs formation and Maillard reaction. It was proved in an electron paramagnetic experiment where the authors observed that antioxidants inhibit radical reactions which take part in HAAs generation [40]. To date, over 100 types of different substances have been tested for possible usage in the inhibition of HAAs formation. One of them was vitamin E. It was shown that vitamin E can efficiently reduce the IQ formation in meat [41]. The addition of vitamin E in a concentration of 1–10% directly onto the meat surface decreased HAAs content in fried beef patties in the range from 45% to 75%. Similar results were observed for meat obtained from cows fed with an increased level of vitamin E. In this meat, a smaller number of mutagenic compounds after cooking were measured [42]. Nonetheless, there are also studies documenting that vitamin E may increase the formation of HAAs in meat [43].
Carotenoids slow down the production of carcinogenic substances in heat-treated meat as well. Addition of tomato extract (1000 mg/kg) to meat juice inhibited MeIQx formation by about 13% and 4,8-DiMeIQx formation by 5% [44]. What is interesting, the same extract suppressed generation of those compounds in a chemical model system to a greater extent (36% for MeIQx and 11% for 4,8-DiMeIQx). Thus, the chemical model system is not an optimal way to assess the inhibitory effect of bioactive compounds on the HAAs formation in real food. Similarly, extracts of other fruits and vegetables rich in carotenoids and xanthophylls (carrots, oranges, apricots, Brussels sprouts, peppers, and tomatoes) were found to be effective in terms of decreasing bacterial mutagenic activity caused by IQ by about 27% (in vitro studies) [45].
Water-soluble vitamins may also decrease the production of carcinogens in food. A study performed by Wong et al. [46] revealed that 6 (ascorbic acid, nicotinic acid, biotin, thiamine, pyridoxamine, and pyridoxine) out of 11 tested water-soluble vitamins (ascorbic acid, vitamin B1(thiamin), vitamin B2 (riboflavin), vitamin B3 (nicotinic acid), vitamin B5 (panthotenic acid), vitamin B6 (pyridoxamine), vitamin B6 (pyridoxine), vitamin B6 (pyridoxal), vitamin B7 (biotin), vitamin B9 (folic acid), and vitamin B12) exhibited an effect against HAAs formation. Pyridoxamine (40%) was the most potent followed by niacin and ascorbic acid (20%). The effectiveness of vitamins was not correlated with their antioxidant potency. It was rather related with their capacity to trap main intermediate compounds for HAAs formation. For instance, pyridoxamine was found to neutralize phenylacetaldehyde, the main compound identified for PhIP formation.
Furthermore, other substances such as tartaric acid obtained from tamarind suppresses the creation of carcinogens in thermally processed meat as well. It was observed that the addition of turmeric decreased harman and norharman content by almost twofold [47].
Nonetheless, polyphenols are the largest group of antioxidants that elongate the shelf-life of meat [48], elevate its healthiness [49], and decreases carcinogen formation in thermally processed meat. There are many studies documenting the effect of various natural sources of polyphenols such as: Rosa rugosa tea extract [50], grape seed and rosemary extract [51], green tea extract [52], artichoke extract [53], apple peel extract [54], pomegranate seed extract [55], and hawthorn extract [56] on HAAs content in meat. Depending on the extract type, inhibition of individual or total HAAs formation was up to 100%. The method of extract distribution greatly affects its inhibition capability against HAAs generation. It has been reported that spreading 0.3% of apple peel extract on the surface (marinating) was much more effective (83% inhibition of PhIP) than mixing the same amount of extract in whole volumes of beef patties (60% inhibition of PhIP) [54]. Time of meat marination also affects the final content of carcinogens in heat-treated meat. Quelhas et al. [57] have observed that the content of HAAs in meat decreased along with the length of the marinating time. Herbs and spices due to the content of various bioactive compounds i.a., polyphenols also exhibit an inhibitory effect against HAAs formation. For instance, turmeric at 5% concentration decreased harman content by about 94.8% while norharman by about 49.56% [58]. Similar effects were documented for red pepper [59], Sichuan pepper, black cumin [60], rosemary, turmeric, galangal, fingerroot [61], thyme, savory, and oregano [62]. However, not every spice and herb decreases the HAAs content in meat products. Some of them, such as fennel, anise, chili, and black pepper even promote HAAs (DMIP, PhIP, MeIQx, 4,8-DiMeIQx) generation in meat (beef patties) [63]. Nonetheless, some other studies showed inhibitory effects of black pepper or chili pepper towards formation of carcinogens in meat [64][65]. Those contradictions may arise from the fact that the inhibitory effects of herbs and spices depends on the presence and share of individual active substances contained in them, the effects of each can mutually neutralize or intensify. For instance, there are studies documenting the inhibitory effect of pure phenolic compounds (ferulic acid, quercetin, protocatechuic acid, chlorogenic acid, p-coumaric acid, quercetin, luteolin, naringenin, and rutin) in terms of suppressing the generation of HAAs in meat [66][67][68]. Some of those compounds mitigate formation of carcinogenic compounds up to 100%. However, there are also phenolic compounds such as carnosic acid and rosmarinic acid which elevate HAAs formation in chemical model systems [69]. Zeng et al. [67] who compared six pure phenolic compounds (ferulic acid, chlorogenic acid, protocatechuic acid, p-coumaric acid, rutin, and luteolin) observed increased content of HAAs in roasted beef patties enriched with pure phenolic acids. The authors mixed them with each other to mimic the composition of selected herbs, and stated that the mixtures of those phenolic compounds needed to be tested rather than single compounds in order to accurately estimate their effect on HAAs level in meat. Moreover, it has been proved that the antioxidant potential of polyphenols is not correlated with their impact on the final content of carcinogens in meat [66]. Studies on 25 phenolic compounds revealed that the most effective inhibitors of PhIP formation were polyphenols that contain two hydroxyl groups at meta positions in aromatic rings [70]. Substitution of those with carboxylic or alkyl groups reduces their inhibitory effect [71]. In turn, addition of hydroxyl and amino groups completely reduces the inhibitory effect of polyphenols. Similarly, ortho and para dihydroxy derivatives were not as effective as PhIP inhibitors. Moreover, alcohol promotes the inhibitory effect of polyphenols against HAAs generation. Viegas et al. [72] have reported that marination of beef with mixtures containing wine and herbs was more effective against (73.5%) heterocyclic aromatic amines formation in comparison to marination with dealcoholized wine and herbs (53.4%).
3. Digestion Significantly Increases the Number of Total Carcinogens Detected in Heat-Treated Meat
The studies on the content of heterocyclic aromatic amines and polycyclic aromatic hydrocarbons concern almost exclusively the content of free compounds. However, due to high temperature, HAAs can be adsorbed on the surfaces of polymeric structures such as proteins or glycogen. Moreover, HAAs can bind with peptides and proteins, what results in the formation of different esters. Kataoka et al. [73] observed that free PhIP under high temperatures reacts with free albumin creating bound HAAs. Thus, it has been hypothesized that HAAs are bound in heat-treated food and thus they can be released under the action of digestive enzymes. This hypothesis was proven by Szterk [74] who treated grilled beef with digestive enzymes (in vitro study). He observed a gradual increase in HAAs content within the passage of a meat sample through each section of the gastrointestinal tract. The increase was observed even though the enzymatic hydrolysis led to partial degradation of proteins and triacylglycerols, what in turn made the biological matrix more complex. Thus, the preparation of samples as well as analysis was way more complicated. Similarly, Chen et al. [75] observed increased concentration of HAAs in smoked sausages after acid hydrolysis. There are only few studies evaluating the impact of selected factors on the accessibility of HAAs from meat subjected to in vitro digestion. Kulp et al. [76] simulated an artificial digestion tract in three locations: mouth (amylase), stomach (pepsin), and small intestine (pancreatin) to determine the percentage release of free HAAs from cooked skinless chicken breast meat and top round “London broil” steaks. They have noted that both amylase and pepsin did not degrade bound HAAs. In turn, pancreatin significantly increased content of free HAAs in meat samples up to 6.6-fold. The bioaccessibility of HAAs was dependent on its polarity. Highly polar compounds were released more efficiently (2.4-fold increase for MeIQx; 3.6—IFP; 6.4—PhIP; and 6.6—DiMeIQx). In the case of low polar carcinogens such as PhIP, major improvement in accessibility (52%) after reduction of particle size of meat made before digestion was observed. Another factor affecting the final content of free HAAs in digested samples was the concentration of pancreatic enzymes. A higher number of enzymes favored HAAs release. Furthermore, increased doneness of meat contributes to lower HAAs bioaccessibility. Shrinkage in well-done meat causes stronger binding of HAAs with meat proteins. Nonetheless, the lower release of carcinogens in that meat did not mean the lower exposure to those compounds because well-done meat is characterized overall by a higher content of HAAs. The meat type also has a great impact on the rate of carcinogens release during digestion. A higher release of MeIQx and DiMeIQx was noted for beef in comparison to chicken meat. However, the cause for these differences is unknown. The authors hypothesized that it is related to the composition differences (fat and protein content ratio) between those types of meat. A modified version of the above-mentioned study was conducted by Kim et al. [77]. In the mentioned study, the researchers built an artificial gastrointestinal digestion tract similar to the previous that simulated the mouth, stomach, small and large intestines but they additionally introduced an agar-based solution of two bacteria Escherichia coli and Lactobacillus sakei separately or as a mixture of both at the stage of intestinal digestion. The criterium for selecting bacteria for study was their domination in microflora. Both chosen bacteria covered a major share of more than 400 species of microorganisms in the human digestive system. What is interesting, the researchers observed the significant decrease in both heterocyclic aromatic amines’ and nitrosamines’ concentrations already at the stage of stomach digestion. Even higher reduction in HAAs content was observed after digestion in the large intestine with enterobacteria for HAAs and with each bacterium (enterobacteria, L. casei, and a mix of both) for nitrosamines. The authors did not propose a clear mechanism behind the decrease in HAAs level after in vitro digestion, but they hypothesized a few possible options instead. The first is based on the rapid change in pH between each section of the digestive tract, especially in the case of the stomach. Salvia in the mouth has a pH of about 6.8, but in the stomach the pH drops drastically to 1.5–3 and then is elevated tremendously by the bile salts to the level of 7–8. Bianchi et al. [78] proved that the degradation of HAAs is triggered by low pH (2.8). The second theory implies that bacteria may decrease the content of carcinogens by binding HAAs (especially lactic acid bacteria) or reducing their content with their own enzymes. For instance, E. coli contains two nitrite reductases and siroheme-dependent reductase which degrade nitrates to nitric oxide. The effect of pH on HAAs content was also tested by Xue et al. [79]. Based on the obtained data, the authors stated that pH, electrolytes, and sample volume had weak effect on in vitro digestion of HAAs. However, they observed a significant effect of enzyme concentration on the rate of HAAs release after digestion. A double enzyme dose increased free HAAs content by 1.6 times. Moreover, there was no significant change in HAAs level after stomach digestion, suggesting that pepsin does not break bonds between proteins and HAAs. In turn, similarly to previous studies, they observed a higher release of free HAAs after intestinal digestion.
References
- Alaejos, M.S.; Afonso, A.M. Factors That Affect the Content of Heterocyclic Aromatic Amines in Foods. Compr. Rev. Food Sci. Food Saf. 2011, 10, 52–108.
- Oz, F.; Kaya, M. Heterocyclic Aromatic Amines in meat. J. Food Process. Preserv. 2011, 35, 739–753.
- Haskaraca, G.; Demirok, E.; Kolsarıcı, N.; Öz, F.; Özsaraç, N. Effect of green tea extract and microwave pre-cooking on the formation of heterocyclic aromatic amines in fried chicken meat products. Food Res. Int. 2014, 63, 373–381.
- Fu, Y.; Zhao, G.; Wang, S.; Yu, J.; Xie, F.; Wang, H.; Xie, J. Simultaneous determination of fifteen heterocyclic aromatic amines in the urine of smokers and nonsmokers using ultra-high performance liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2014, 1333, 45–53.
- Trafialek, J.; Kolanowski, W. Dietary exposure to meat-related carcinogenic substances: Is there a way to estimate the risk? Int. J. Food Sci. Nutr. 2014, 65, 774–780.
- Reinik, M.; Tamme, T.; Roasto, M.; Juhkam, K.; Tenno, T.; Kiis, A. Polycyclic aromatic hydrocarbons (PAHs) in meat products and estimated PAH intake by children and the general population in Estonia. Food Addit. Contam. 2007, 24, 429–437.
- Ciemniak, A. A comparison of N-nitrosodimethylamine contents in selected meat products. Rocz. Panstw. Zakl. Hig. 2006, 57, 341–346.
- Park, J.; Seo, J.; Lee, J.; Kwon, H. Erratum to ‘Distribution of Seven N-Nitrosamines in Food’ . Toxicol. Res. 2018, 34, 371.
- Domingo, J.L.; Nadal, M. Carcinogenicity of consumption of red and processed meat: What about environmental contaminants? Environ. Res. 2016, 145, 109–115.
- Yang, D.; He, Z.; Gao, D.; Qin, F.; Deng, S.; Wang, P.; Xu, X.; Chen, J.; Zeng, M. Effects of smoking or baking procedures during sausage processing on the formation of heterocyclic amines measured using UPLC-MS/MS. Food Chem. 2019, 276, 195–201.
- Zhang, Y.; Yu, C.; Mei, J.; Wang, S. Formation and mitigation of heterocyclic aromatic amines in fried pork. Food Addit. Contam. Part A 2013, 30, 1501–1507.
- Keating, G.A.; Layton, D.W.; Felton, J.S. Factors determining dietary intakes of heterocyclic amines in cooked foods. Mutat. Res./Genet. Toxicol. Environ. Mutagenes. 1999, 443, 149–156.
- Solyakov, A.; Skog, K. Screening for heterocyclic amines in chicken cooked in various ways. Food Chem. Toxicol. 2002, 40, 1205–1211.
- Chiu, C.P.; Yang, D.Y.; Chen, B.H. Formation of Heterocyclic Amines in Cooked Chicken Legs. J. Food Prot. 1998, 61, 712–719.
- Jägerstad, M.; Skog, K.; Arvidsson, P.; Solyakov, A. Chemistry, formation and occurrence of genotoxic heterocyclic amines identified in model systems and cooked foods. Z. Für Lebensm. Und-Forsch. A 1998, 207, 419–427.
- Kondjoyan, A.; Chevolleau, S.; Grève, E.; Gatellier, P.; Santé-Lhoutellier, V.; Bruel, S.; Touzet, C.; Portanguen, S.; Debrauwer, L. Formation of heterocyclic amines in slices of Longissimus thoracis beef muscle subjected to jets of superheated steam. Food Chem. 2010, 119, 19–26.
- Oz, F.; Kotan, G. Effects of different cooking methods and fat levels on the formation of heterocyclic aromatic amines in various fishes. Food Control 2016, 67, 216–224.
- Guo, H.; Wang, Z.; Pan, H.; Li, X.; Chen, L.; Rao, W.; Gao, Y.; Zhang, D. Effects of traditional chinese cooking methods on formation of heterocyclic aromatic amines in lamb patties. Food Sci. Biotechnol. 2014, 23, 747–753.
- Omojola, A.B.; Ahmed, S.A.; Attoh-Kotoku, V.; Wogar, G.I. Effect of cooking methods on cholesterol, mineral composition and formation of total heterocyclic aromatic amines in Muscovy drake meat. J. Sci. Food Agric. 2015, 95, 98–102.
- Farhadian, A.; Jinap, S.; Abas, F.; Sakar, Z.I. Determination of polycyclic aromatic hydrocarbons in grilled meat. Food Control 2010, 21, 606–610.
- Zorrilla, S.E.; Singh, R.P. Heat transfer in double-sided cooking of meat patties considering two-dimensional geometry and radial shrinkage. J. Food Eng. 2003, 57, 57–65.
- Persson, E.; Sjöholm, I.; Skog, K. Heat and mass transfer in chicken breasts—Effect on PhIP formation. Eur. Food Res. Technol. 2002, 214, 455–459.
- Jinap, S.; Mohd-Mokhtar, M.S.; Farhadian, A.; Hasnol, N.D.S.; Jaafar, S.N.; Hajeb, P. Effects of varying degrees of doneness on the formation of Heterocyclic Aromatic Amines in chicken and beef satay. Meat Sci. 2013, 94, 202–207.
- EFSA. Polycyclic Aromatic Hydrocarbons in Food—Scientific Opinion of the Panel on Contaminants in the Food Chain. EFSA J. 2008, 6, 724.
- Bordas, M.; Moyano, E.; Puignou, L.; Galceran, M.T. Formation and stability of heterocyclic amines in a meat flavour model system. J. Chromatogr. B 2004, 802, 11–17.
- Turesky, R.J.; Taylor, J.; Schnackenberg, L.; Freeman, J.P.; Holland, R.D. Quantitation of Carcinogenic Heterocyclic Aromatic Amines and Detection of Novel Heterocyclic Aromatic Amines in Cooked Meats and Grill Scrapings by HPLC/ESI-MS. J. Agric. Food Chem. 2005, 53, 3248–3258.
- Lee, J.-G.; Kim, S.-Y.; Moon, J.-S.; Kim, S.-H.; Kang, D.-H.; Yoon, H.-J. Effects of grilling procedures on levels of polycyclic aromatic hydrocarbons in grilled meats. Food Chem. 2016, 199, 632–638.
- Skog, K.; Jägerstad, M. Incorporation of carbon atoms from glucose into the food mutagens MeIQx and 4,8-DiMeIQx using 14C-labelled glucose in a model system. Carcinogenesis 1993, 14, 2027–2031.
- Cao, H.; Chen, B.H.; Inbaraj, B.S.; Chen, L.; Alvarez-Rivera, G.; Cifuentes, A.; Zhang, N.; Yang, D.J.; Simal-Gandara, J.; Wang, M.; et al. Preventive potential and mechanism of dietary polyphenols on the formation of heterocyclic aromatic amines. Food Front. 2020, 1, 134–151.
- Jackson, L.S.; Hargraves, W.A.; Stroup, W.H.; Diachenko, G.W. Heterocyclic aromatic amine content of selected beef flavors. Mutat. Res./Genet. Toxicol. 1994, 320, 113–124.
- Jägerstad, M.; Skog, K. Formation of Meat Mutagens. Adv Exp Med Biol. 1991, 289, 83–105.
- Skog, K.; Jagerstad, M. Effects of monosaccharides and disaccharides on the formation of food mutagens in model systems. Mutat. Res./Fundam. Mol. Mech. Mutagenes. 1990, 230, 263–272.
- Olsson, V.; Solyakov, A.; Skog, K.; Lundström, K.; Jägerstad, M. Natural Variations of Precursors in Pig Meat Affect the Yield of Heterocyclic Amines—Effects of RN Genotype, Feeding Regime, and Sex. J. Agric. Food Chem. 2002, 50, 2962–2969.
- Zeng, M.; Wang, J.; Zhang, M.; Chen, J.; He, Z.; Qin, F.; Xu, Z.; Cao, D.; Chen, J. Inhibitory effects of Sichuan pepper (Zanthoxylum bungeanum) and sanshoamide extract on heterocyclic amine formation in grilled ground beef patties. Food Chem. 2018, 239, 111–118.
- Sinha, R.; Rothman, N.; Brown, E.D.; Salmon, C.P.; Knize, M.G.; Swanson, C.A.; Rossi, S.C.; Mark, S.D.; Levander, O.A.; Felton, J.S. High concentrations of the carcinogen 2-amino-1-methyl-6-phenylimidazo-pyridine (PhIP) occur in chicken but are dependent on the cooking method. Cancer Res. 1995, 55, 4516–4519.
- Polak, M.L.; Demšar, L.; Zahija, I.; Polak, T. Influence of temperature on the formation of heterocyclic aromatic amines in pork steaks. Czech J. Food Sci. 2020, 38, 248–254.
- Persson, E.; Oroszvári, B.K.; Tornberg, E.; Sjöholm, I.; Skog, K. Heterocyclic amine formation during frying of frozen beefburgers. Int. J. Food Sci. Technol. 2008, 43, 62–68.
- Knize, M.G.; Andresen, B.D.; Healy, S.K.; Shen, N.H.; Lewis, P.R.; Bjeldanes, L.F.; Hatch, F.T.; Felton, J.S. Effects of temperature, patty thickness and fat content on the production of mutagens in fried ground beef. Food Chem. Toxicol. 1985, 23, 1035–1040.
- Zamora, R.; Alcón, E.; Hidalgo, F.J. Effect of lipid oxidation products on the formation of 2-amino-1-methyl-6-phenylimidazopyridine (PhIP) in model systems. Food Chem. 2012, 135, 2569–2574.
- Kikugawa, K. Involvement of free radicals in the formation of heterocyclic amines and prevention by antioxidants. Cancer Lett. 1999, 143, 123–126.
- Balogh, Z.; Gray, J.I.; Gomaa, E.A.; Booren, A.M. Formation and inhibition of heterocyclic aromatic amines in fried ground beef patties. Food Chem. Toxicol. 2000, 38, 395–401.
- Ruan, E.D.; Juárez, M.; Thacker, R.; Yang, X.; Dugan, M.E.R.; Aalhus, J.L. Dietary vitamin E effects on the formation of heterocyclic amines in grilled lean beef. Meat Sci. 2014, 96, 849–853.
- Lan, C.M.; Kao, T.H.; Chen, B.H. Effects of heating time and antioxidants on the formation of heterocyclic amines in marinated foods. J. Chromatogr. B 2004, 802, 27–37.
- Vitaglione, P.; Monti, S.; Ambrosino, P.; Skog, K.; Fogliano, V. Carotenoids from tomatoes inhibit heterocyclic amine formation. Eur. Food Res. Technol. 2002, 215, 108–113.
- Rauscher, R.; Edenharder, R.; Platt, K.L. In vitro antimutagenic and in vivo anticlastogenic effects of carotenoids and solvent extracts from fruits and vegetables rich in carotenoids. Mutat. Res./Genet. Toxicol. Environ. Mutagenes. 1998, 413, 129–142.
- Wong, D.; Cheng, K.-W.; Wang, M. Inhibition of heterocyclic amine formation by water-soluble vitamins in Maillard reaction model systems and beef patties. Food Chem. 2012, 133, 760–766.
- Sepahpour, S.; Selamat, J.; Khatib, A.; Manap, M.Y.A.; Razis, A.F.A.; Hajeb, P. Inhibitory effect of mixture herbs/spices on formation of heterocyclic amines and mutagenic activity of grilled beef. Food Addit. Contam. Part A 2018, 35, 1911–1927.
- Pogorzelska-Nowicka, E.D.; Brodowska, M.; Górska-Horczyczak, E.; Godziszewska, J.; Sakowska, A.; Wojtasik-Kalinowska, I.; Wierzbicka, A. Physicochemical and Biochemical Properties of Ground Pork Formulated with Addition of Prunus cerasus (cv Montmorency) Extract and Subjected to Freezing Storage. Acta Univ. Cibiniensis. Ser. E Food Technol. 2019, 23, 167–178.
- Yeung, A.W.K.; Tzvetkov, N.T.; El-Demerdash, A.; Horbanczuk, O.K.; Das, N.; Pirgozliev, V.; Lucarini, M.; Durazzo, A.; Souto, E.B.; Santini, A.; et al. Apple polyphenols in human and animal health. Anim. Sci. Pap. Rep. 2021, 39, 105–118.
- Jamali, M.; Zhang, Y.; Teng, H.; Li, S.; Wang, F.; Peng, Z. Inhibitory Effect of Rosa rugosa Tea Extract on the Formation of Heterocyclic Amines in Meat Patties at Different Temperatures. Molecules 2016, 21, 173.
- Gibis, M.; Weiss, J. Antioxidant capacity and inhibitory effect of grape seed and rosemary extract in marinades on the formation of heterocyclic amines in fried beef patties. Food Chem. 2012, 134, 766–774.
- Cheng, K.W.; Wong, C.C.; Chao, J.; Lo, C.; Chen, F.; Chu, I.K.; Che, C.M.; Ho, C.T.; Wang, M. Inhibition of mutagenic PhIP formation by epigallocatechin gallate via scavenging of phenylacetaldehyde. Mol. Nutr. Food Res. 2009, 53, 716–725.
- Tengilimoglu-Metin, M.M.; Kizil, M. Reducing effect of artichoke extract on heterocyclic aromatic amine formation in beef and chicken breast meat. Meat Sci. 2017, 134, 68–75.
- Sabally, K.; Sleno, L.; Jauffrit, J.-A.; Iskandar, M.M.; Kubow, S. Inhibitory effects of apple peel polyphenol extract on the formation of heterocyclic amines in pan fried beef patties. Meat Sci. 2016, 117, 57–62.
- Keşkekoğlu, H.; Üren, A. Inhibitory effects of pomegranate seed extract on the formation of heterocyclic aromatic amines in beef and chicken meatballs after cooking by four different methods. Meat Sci. 2014, 96, 1446–1451.
- Tengilimoglu-Metin, M.M.; Hamzalioglu, A.; Gokmen, V.; Kizil, M. Inhibitory effect of hawthorn extract on heterocyclic aromatic amine formation in beef and chicken breast meat. Food Res. Int. 2017, 99, 586–595.
- Quelhas, I.; Petisca, C.; Viegas, O.; Melo, A.; Pinho, O.; Ferreira, I.M.P.L.V.O. Effect of green tea marinades on the formation of heterocyclic aromatic amines and sensory quality of pan-fried beef. Food Chem. 2010, 122, 98–104.
- Wang, Q.; Li, J.; Li, K.; Li, C. Effects of turmeric on reducing heterocyclic aromatic amines in Chinese tradition braised meat products and the underlying mechanism. Food Sci. Nutr. 2021, 9, 5575–5582.
- Oz, F.; Kaya, M. The inhibitory effect of Red Pepper on Heterocyclic Aromatic Amines in fried beef Longissimus dorsi muscle. J. Food Process. Preserv. 2011, 35, 806–812.
- Oz, E. Inhibitory effects of black cumin on the formation of heterocyclic aromatic amines in meatball. PLoS ONE 2019, 14, e0221680.
- Puangsombat, K.; Jirapakkul, W.; Smith, J.S. Inhibitory Activity of Asian Spices on Heterocyclic Amines Formation in Cooked Beef Patties. J. Food Sci. 2011, 76, T174–T180.
- Damašius, J.; Venskutonis, P.R.; Ferracane, R.; Fogliano, V. Assessment of the influence of some spice extracts on the formation of heterocyclic amines in meat. Food Chem. 2011, 126, 149–156.
- Zeng, M.; He, Z.; Zheng, Z.; Qin, F.; Tao, G.; Zhang, S.; Gao, Y.; Chen, J. Effect of Six Chinese Spices on Heterocyclic Amine Profiles in Roast Beef Patties by Ultra Performance Liquid Chromatography-Tandem Mass Spectrometry and Principal Component Analysis. J. Agric. Food Chem. 2014, 62, 9908–9915.
- Zeng, M.; Zhang, M.; Chen, J.; He, Z.; Qin, F.; Hu, C.; Xu, H.; Tao, G.; Zhang, S.; Chen, J. UPLC-MS/MS and multivariate analysis of inhibition of heterocyclic amine profiles by black pepper and piperine in roast beef patties. Chemom. Intell. Lab. Syst. 2017, 168, 96–106.
- Xue, C.; He, Z.; Qin, F.; Chen, J.; Zeng, M. Effects of amides from pungent spices on the free and protein-bound heterocyclic amine profiles of roast beef patties by UPLC–MS/MS and multivariate statistical analysis. Food Res. Int. 2020, 135, 109299.
- Zhu, Q.; Zhang, S.; Wang, M.; Chen, J.; Zheng, Z.-P. Inhibitory effects of selected dietary flavonoids on the formation of total heterocyclic amines and 2-amino-1-methyl-6-phenylimidazopyridine (PhIP) in roast beef patties and in chemical models. Food Funct. 2016, 7, 1057–1066.
- Zeng, M.; Li, Y.; He, Z.; Qin, F.; Chen, J. Effect of phenolic compounds from spices consumed in China on heterocyclic amine profiles in roast beef patties by UPLC–MS/MS and multivariate analysis. Meat Sci. 2016, 116, 50–57.
- Zeng, M.; Li, Y.; He, Z.; Qin, F.; Tao, G.; Zhang, S.; Gao, Y.; Chen, J. Discrimination and investigation of inhibitory patterns of flavonoids and phenolic acids on heterocyclic amine formation in chemical model systems by UPLC-MS profiling and chemometrics. Eur. Food Res. Technol. 2016, 242, 313–319.
- Cheng, K.-W.; Chen, F.; Wang, M. Inhibitory activities of dietary phenolic compounds on heterocyclic amine formation in both chemical model system and beef patties. Mol. Nutr. Food Res. 2007, 51, 969–976.
- Salazar, R.; Arámbula-Villa, G.; Hidalgo, F.J.; Zamora, R. Structural characteristics that determine the inhibitory role of phenolic compounds on 2-amino-1-methyl-6-phenylimidazopyridine (PhIP) formation. Food Chem. 2014, 151, 480–486.
- Gibis, M. Heterocyclic Aromatic Amines in Cooked Meat Products: Causes, Formation, Occurrence, and Risk Assessment. Compr. Rev. Food Sci. Food Saf. 2016, 15, 269–302.
- Viegas, O.; Amaro, L.F.; Ferreira, I.M.P.L.V.O.; Pinho, O. Inhibitory Effect of Antioxidant-Rich Marinades on the Formation of Heterocyclic Aromatic Amines in Pan-Fried Beef. J. Agric. Food Chem. 2012, 60, 6235–6240.
- Kataoka, H.; Miyake, M.; Nishioka, S.; Matsumoto, T.; Saito, K.; Mitani, K. Formation of protein adducts of 2-amino-1-methyl-6-phenylimidazopyridine in cooked foods. Mol. Nutr. Food Res. 2010, 54, 1039–1048.
- Szterk, A. Chemical state of heterocyclic aromatic amines in grilled beef: Evaluation by in vitro digestion model and comparison of alkaline hydrolysis and organic solvent for extraction. Food Chem. Toxicol. 2013, 62, 653–660.
- Chen, J.; He, Z.; Qin, F.; Chen, J.; Zeng, M. Formation of Free and Protein-Bound Heterocyclic Amines in Roast Beef Patties Assessed by UPLC-MS/MS. J. Agric. Food Chem. 2017, 65, 4493–4499.
- Kulp, K.S.; Fortson, S.L.; Knize, M.G.; Felton, J.S. An in vitro model system to predict the bioaccessibility of heterocyclic amines from a cooked meat matrix. Food Chem. Toxicol. 2003, 41, 1701–1710.
- Kim, H.S.; Hur, S.J. Changes in the mutagenicity of heterocyclic amines, nitrite, and N-nitroso compound in pork patties during in vitro human digestion. LWT 2018, 92, 47–53.
- Bianchi, F.; Careri, M.; Corradini, C.; Elviri, L.; Mangia, A.; Zagnoni, I. Investigation of the separation of heterocyclic aromatic amines by reversed phase ion-pair liquid chromatography coupled with tandem mass spectrometry: The role of ion pair reagents on LC–MS/MS sensitivity. J. Chromatogr. B 2005, 825, 193–200.
- Xue, C.; Chen, Q.; He, Z.; Qin, F.; Wang, Z.; Chen, J.; Zeng, M. Release mechanism between sarcoplasmic protein–bound and free heterocyclic amines and the effects of dietary additives using an in-vitro digestion model. Food Chem. 2022, 377, 131993.
More
Information
Subjects:
Food Science & Technology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
01 Aug 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No