Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Darya Khlebnikova | -- | 2201 | 2022-03-31 23:08:20 | | | |
| 2 | Vivi Li | + 35 word(s) | 2236 | 2022-04-01 06:20:06 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Khlebnikova, D.; , .; Danilova, N. Flavonoid Accumulation in Summer Savory (Satureja hortensis L.). Encyclopedia. Available online: https://encyclopedia.pub/entry/21252 (accessed on 28 June 2026).
Khlebnikova D, , Danilova N. Flavonoid Accumulation in Summer Savory (Satureja hortensis L.). Encyclopedia. Available at: https://encyclopedia.pub/entry/21252. Accessed June 28, 2026.
Khlebnikova, Darya, , Nina Danilova. "Flavonoid Accumulation in Summer Savory (Satureja hortensis L.)" Encyclopedia, https://encyclopedia.pub/entry/21252 (accessed June 28, 2026).
Khlebnikova, D., , ., & Danilova, N. (2022, March 31). Flavonoid Accumulation in Summer Savory (Satureja hortensis L.). In Encyclopedia. https://encyclopedia.pub/entry/21252
Khlebnikova, Darya, et al. "Flavonoid Accumulation in Summer Savory (Satureja hortensis L.)." Encyclopedia. Web. 31 March, 2022.
Copy Citation
Summer savory (Satureja hortensis L.) is a medicinal and aromatic plant of the Lamiaceae family, a source of valuable secondary metabolites (monoterpenoids, rosmarinic acid, flavonoids). The main components of the phytochemical profile are the monoterpenoids of the p-menthane group (thymol, carvacrol, p-cymene, and γ-terpinen); in addition, the aerial parts of plants are rich in flavonoids and contain rosmarinic acid
Satureja hortensis
in vitro culture
callus
flavonoids
light-emitting diodes
spectral light composition
1. Introduction
Plants remain an indispensable raw material for the light and food industry, and a source of many valuable bioactive molecules/pharmacophores for humans [1][2]. The market for herbal medicines is rapidly growing; more than a quarter of all pharmaceuticals in industrialized countries are of herbal origin. According to a BBC report, the herbal medicines market will grow from $29.4 billion in 2017 to about $39.6 billion by 2022, with an annual growth rate of 6.1% [3]. However, technologies for industrial production of valuable medicinal molecules or substances based on plant cell and tissue cultures are not as widespread as the production of economically beneficial metabolites and cell biomass for the cosmetics and food industry; despite the fact that the need for these technologies increases from year to year [4]. The development of such biotechnological processes is based on extensive knowledge of plant physiology and biochemistry under in vitro conditions, molecular biology, and bioengineering. Thus, expanding the knowledge base for the characteristics of in vitro cultivation of medicinal, aromatic, and valuable agricultural plants, especially their cell cultures, contributes to the development of this industry sector and is an urgent task to research [4].
Summer savory (Satureja hortensis L.) is a herbaceous essential oil plant of the mint family (Lamiaceae). It is used in the traditional medicine of the Middle East, evidence-based medicine, cosmetology, and the food industry as condiments. Summer savory extracts and essential oils have a wide range of biological activities—antimicrobial, antiviral, antioxidant, fungicidal, antinociceptive, antitumor, hypoglycemic, hyperlipidemic, inhibition of amyloid beta protein aggregation, etc. [5]. The main components of the phytochemical profile are the monoterpenoids of the p-menthane group (thymol, carvacrol, p-cymene, and γ-terpinen); in addition, the aerial parts of plants are rich in flavonoids and contain rosmarinic acid [5][6].
Studies have revealed some biological activities associated with the accumulation of flavonoids and phenylpropanoids of summer savory. Mchedlishvili et al. (2005) found that the flavonoid fraction of S. hortensis lowers serum cholesterol levels in rabbits [7]. Accumulation of phenolic compounds (flavonoids and rosmarinic acid) determines the hypoglycemic activity of extracts; studies of local plant populations were conducted in Georgia; as a result, the drug Saturin was developed [6]. The capsule contains an aqueous extract and fine leaf powder. The drug is registered with the Ministry of Labour, Health and Social Welfare of Georgia as a drug for the treatment of diabetes mellitus type 2 [6][8]. Summer savory is actively used in traditional (for the Middle East) and evidence-based medicine. This was the reason for studying its qualitative and quantitative flavonoid content by research teams from different countries like Georgia, Serbia, Romania, Turkey, Finland, Greece, and Russia [6][7][8][9][10][11][12][13][14][15][16][17]. However, there are no data on the flavonoid accumulation in the aseptic culture of summer savory.
Light is an essential abiotic elicitor that affects many physiological processes in the plant, determining growth and development features [18]. Thus, changes in plant morphology and increased production of secondary metabolites in response to the use of different spectrum light sources have been studied in several species [19][20][21][22][23]. The response to different lighting regimes is often caused due to the fact that flavonoid biosynthesis occurs via the phenylpropanoid pathway and is regulated by numerous enzymes (phenylalanine ammonia lyase (PAL), chalcone synthase (CHS), chalcone isomerase (CHI), and flavonol synthase (FLS) genes, etc.), whose activity is both directly and indirectly induced by lighting parameters like spectral composition and intensity [24][25]. Cultivation under monochromatic light sources makes it possible to reveal the influence of one part or another of the spectrum on the course of flavonoid biosynthesis. Thus, cultivation under blue LEDs (460–470 nm) resulted in increased flavonoid accumulation in several species compared to other monochrome light modes (green, red, and yellow) in Anoectochilus roxburghii (Wall.) Lindl. [26], Pisum sativum L. [27], and Glycine max L. Merr. plantlets [28]. The combination of blue and red light also led to an increase in flavonoid accumulation in Anoectochilus roxburghii due to the activation of CHI and FLS genes expression [29].
2. Flavonoid Accumulation in Aseptic Plants
In vitro cultivation of S. hortensis aseptic plants. Aseptic plantlets of S. hortensis were cultivated into test tubes for 30 days; by the end of the initial culture, the plants reached a height of 6.5–7.5 cm and formed 3–5 pairs of true leaves (Figure 1a). Initial culture plants were divided into segments of 2–3 nodes and transplanted for further cultivation. By the end of the 1st subculture, the plants reached a height of 11–13 cm, formed 7–9 nodes and 3–5 lateral shoots. Plants of the 1st subculture developed from segments containing one or two nodes of the upper, middle, or lower parts of the initial culture plants, and after 30 days of cultivation reached a height of 15–20 cm, formed 6–9 nodes and 2–6 lateral shoots, the root system was actively developing. Most plants formed flower buds at the end of the 2nd subculture (Figure 1b) regardless of the segment origin at cutting (nodes of the upper, middle, or lower part of the mother plant). Flowering in plants of the 3rd subculture began after an average of 10 days of cultivation, and by the end of the subculture period, most plants were in the phase of mass flowering (Figure 1c,d). In contrast to Pistelli et al., 2013, researchers cultivated aseptic plants of summer savory on MS medium without auxins, since root formation was active and the addition of auxins was not necessary (Figure 1a,d) [30].

Figure 1. Appearance of S. hortensis aseptic plants: (a) initial culture, (b) 2nd subculture, (c) 3rd subculture, and (d) aseptic plant shoot of the 3rd subculture with flower buds and flowers. Bars—0.5 cm.
Flavonoid content in S. hortensis aseptic plants. The total flavonoid content in different organs was measured during three subcultures to study the characteristics of flavonoid accumulation in S. hortensis aseptic plants (Table 1).
Table 1. Total flavonoid content in various parts of S. hortensis aseptic plants.
| Plant Material | Subculture | Total Flavonoid Content, mg/g FW |
|---|---|---|
| Leaf | 0 | 8.17 ± 1.07 |
| Stem | 0 | 0.34 ± 0.04 |
| Leaf | 1 | 8.20 ± 0.78 |
| Stem | 1 | 0.93 ± 0.15 |
| Leaf | 2 | 8.35 ± 0.17 |
| Stem | 2 | 1.50 ± 0.22 |
| Flower bud | 2 | 7.55 ± 0.29 |
| Leaf | 3 | 7.02 ± 0.90 |
| Stem | 3 | 1.58 ± 0.76 |
| Corolla | 3 | 0.78 ± 0.12 |
| Calyx | 3 | 5.27 ± 0.28 |
Thus, in leaves of aseptic plants, the flavonoid content was 24 times higher than in stems at the end of the initial culture. By the end of the 2nd subculture, it increased insignificantly in leaves, while in stems, it increased almost fivefold; high flavonoids in flower buds were also noted. At the end of the 3rd subculture, there was a significant flavonoid decrease in leaves (by 16%) and an insignificant increase in stems (by 5%); in the corolla, the flavonoid content did not differ significantly from that in stems. In the calyx, it was noticeably inferior to the content in leaves (by 25%) and significantly superior to the content in stems and corolla (by 70% and 85%, respectively).
The total flavonoid content in flower buds of the 2nd subculture plants is higher in comparison to the content in stems. This can be explained by the accumulation of flavonoids in the photosynthetic part of the flower–calyx, and not in the forming corolla. Thus, the flavonoid accumulation in S. hortensis aseptic plants occurs mainly in leaves, flower buds, and calyx; much less is accumulated in stems and corolla of the flower. The flavonoid contents in leaves did not change during cultivation in three subcultures. The organ-specific flavonoid accumulation researchers observed in S. hortensis aseptic plants has been noted by many researchers for several species [31][32].
The study of the flavonoid accumulation in aseptic culture of summer savory has not been previously carried out, however, there is data on the flavonoid content in plants grown under in vivo conditions. According to Masković et al., 2017, the flavonoid content in S. hortensis, depending on the extraction method, can vary from 5.23 ± 0.76 to 28.42 ± 0.29 mg/g of dry extract in routine equivalent [13]. Summer savory growing in Georgia accumulated 1.6–1.7% of flavonoids and phenolcarboxylic acids in air-dry leaves and flowers collected during budding and flowering [6].
3. Flavonoid Accumulation in Callus Tissue
Induction and cultivation of S. hortensis callus tissue. Two types of explants—cotyledon leaves and hypocotyls—were cultured on an MS medium with the addition of 1 mg/L BAP to obtain primary callus. The callus formed a light green color with small yellow areas, heterogeneous in density; looser and denser areas could be distinguished; shoot and root organogenesis took place actively (Figure 2).

Figure 2. Appearance of the primary callus and regenerant plants of S. hortensis: (a) callus from cotyledon leaves, (b) callus from hypocotyls, and (c) regenerant plants. Bars—0.5 cm.
After 2–3 weeks of explant cultivation, meristematic foci were formed on the callus tissue sites, and small shoots of normal morphology, hyperhydric teratomas, and roots were formed due to organogenesis.
Flavonoid content in callus tissue of S. hortensis. The total flavonoid content in the primary callus after 30 days of cultivation (initial culture) was significantly lower than in the leaves and flower buds of aseptic plants: in callus from cotyledon 0.50 ± 0.09 mg/g FW; in callus on hypocotyl explants 0.44 ± 0.11 mg/g FW. This may be due to the need for cell differentiation for flavonoid biosynthesis [31][32][33]. In favor of this hypothesis, the flavonoid content in regenerants (the shoots of normal morphology and hyperhydric teratomas were analyzed together) formed on the primary callus was 1.26 ± 0.21 mg/g FW.
The type of explant (cotyledon or hypocotyl) did not affect the flavonoid accumulation in the primary callus formed on it. Therefore, cotyledon leaves and hypocotyls can be used as explants to obtain callus culture for flavonoid synthesis.
The callus formed on hypocotyl explants was chosen for further experimentation because visually, the growth of this kind of callus tissue was more active than in cotyledon explants. Primary callus tissue areas were cultured for six subcultures. The callus appearance and consistency changed insignificantly; the tissue remained green with light yellow areas; the callus consistency was quite dense, easily to split into smaller aggregates; no organogenesis was observed (Figure 3).

Figure 3. Changes in the appearance of S. hortensis callus tissue over six subcultures: (a) 1st subculture, (b) 2nd subculture, (c) 3rd subculture, (d) 4th subculture, (e) 5th subculture, and (f) 6th subculture. Bars—0.5 cm.
The data on changes in contents of total flavonoids in the callus tissue over six subcultures are shown in Figure 4.
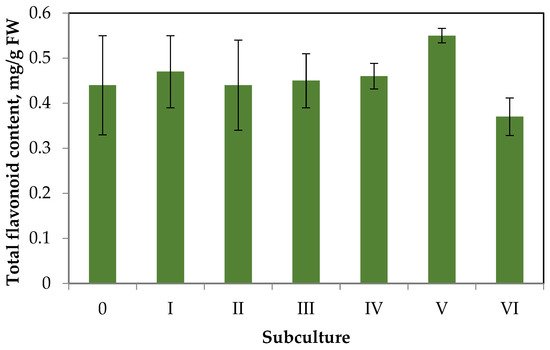
Figure 4. Dynamics of flavonoid accumulation in S. hortensis callus over six subcultures.
Having analyzed the data of Figure 4, researchers can draw the following conclusions. The flavonoid accumulation in callus tissue is much lower than in aseptic plants. This trend is typical for the primary callus tissues of some species, especially in the absence of elicitation [31]. Throughout six subcultures, there were no significant deviations in flavonoid accumulation from the primary callus (content did not exceed 0.6 mg/g FW). Small fluctuations in flavonoid accumulation during the subculture period can be explained by the genetic and morphologic heterogeneity of callus tissue.
4. Effect of Monochromatic Light on Callus Tissue Characteristics and Flavonoid Content
To study the effect of the light regime on flavonoid accumulation, were chosen the following conditions: LED lamps of a monochrome spectrum–blue, green and red.
Appearance and consistency of callus tissue. Callus of the 4th subculture obtained from hypocotyl explants was used for the experiment. Cultivation under blue LEDs during the 5th subculture resulted in the growth of callus tissue of green-yellow color with slight red spots; the callus consistency was slightly watery and friable; it disintegrated into small aggregates, with quite dense areas (Figure 5a). Cultivation under green LEDs led to the formation of an actively growing soft, easily disintegrating into small aggregates, watery callus from a light-yellow color to green. In isolated cases, rhizogenesis was observed; thin roots 2–3 mm in length were formed (Figure 5b). Red light promoted growth of green-yellow callus with small brown areas, with denser consistency than under green light (Figure 5c).

Figure 5. Appearance of callus tissue after cultivation under LED lamps of monochromatic spectrum: (a) blue LEDs; (b) green LEDs; and (c) red LEDs. Bars–0.5 cm.
Flavonoid accumulation in S. hortensis callus tissue under the monochromatic light influence. Cultivation during the 5th subculture under monochromatic LED lamps did not result in significant changes in flavonoid accumulation (Figure 6).
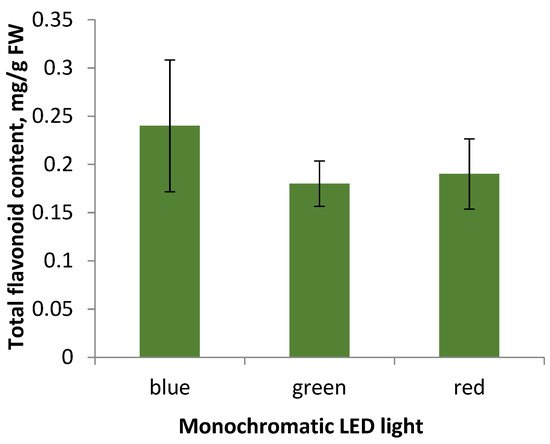
Figure 6. Total flavonoid content in callus tissue of the 5th subculture under different light conditions of cultivation.
Summarizing the data obtained, researchers can conclude that the appearance of callus tissue when cultured under monochrome LED light: blue, green, and red, does not differ significantly, the most actively growing callus under green light is characterized by low levels of flavonoid accumulation, the same as the callus under the other monochrome lamps. Thereby, the factor of lighting conditions in the selected modes did not affect the accumulation of flavonoids in S. hortensis callus tissue samples. However, according to some researchers, the cultivation of aseptic plants under monochrome lighting affected flavonoid accumulation [31]. It may be due to disruption of both the genetic material in the cell and disorders of its expression, which are characteristic of callus tissue as a heterogeneous structure [31].
A number of researchers note an increase of flavonoid accumulation during the cultivation of aseptic plants and cell cultures under blue light [34]. For example, this trend is typical for Saccharina japonica (J.E. Areschoug) C.E. Lane, C. Mayes, Druehl & G.W. Saunders, Saussurea medusa Maxim., and Lactuca sativa L. [34][35][36][37]. In 2018, Li et al. studied the influence of the monochrome spectrum of LEDs, as well as the photoperiod and radiation intensity on the growth of the embryogenic callus of Dimocarpus longan Lour. The following dependence of the flavonoid accumulation on the spectrum of LED lamps was noted–blue light > green light > dark > white light > red light [34].
References
- Dias, D.A.; Urban, S. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336.
- Giri, C.C.; Zaheer, M. Chemical elicitors versus secondary metabolite production in vitro using plant cell, tissue and organ cultures: Recent trends and a sky eye view appraisal. Plant Cell Tissue Organ Cult. 2016, 126, 1–18.
- British Broadcasting Corporation (BBC) Research. Plant-Derived Drugs: Global Markets. Available online: https://www.bccresearch.com/market-research/biotechnology/botanical-and-plant-derived-drugs-global-markets-bio022h.html (accessed on 22 December 2021).
- Eibl, R.; Meier, P.; Stutz, I.; Schildberger, D.; Hühn, T.; Eibl, D. Plant cell culture technology in the cosmetics and food industries: Current state and future trends. Appl. Microbiol. Biotechnol. 2018, 102, 8661–8675.
- Tepe, B.; Cilkiz, M. Pharmacological and phytochemical overview on Satureja. Pharm. Biol. 2016, 54, 375–412.
- Kemertelidze, E.; Sagareishvili, T.; Syrov, V.; Khushbaktova, Z.; Tsutskiridze, L.; Kurashvili, R. Saturin-effective vegetative remedy in treatment of type 2 diabetes mellitus. Georgian Med. News 2012, 2, 47–52.
- Mchedlishvili, D.; Mchedlishvili, D.; Kuchukashvili, Z.; Tabatadze, T. Influence of flavonoids isolated from Satureja hortensis L. on hypercholesterolemic rabbits. Indian J. Pharmacol. 2005, 37, 259–260.
- Kemertelidze, E.; Sagareishvili, T.; Syrov, V.; Khushbaktova, Z. Chemical compounds and pharmacological activity of the leaves of summer savory growing in Georgia. Pharm. Chem. J. 2004, 38, 33–36.
- Exarchou, V.; Nenadis, N.; Tsimidou, M.; Gerothanassis, I.P.; Troganis, A.; Boskou, D. Antioxidant activities and phenolic composition of extracts from Greek oregano, Greek sage, and Summer savory. J. Agric. Food Chem. 2002, 50, 5294–5299.
- Kosar, M.; Dorman, H.J.D.; Hiltunen, R. Effect of an acid treatment on the phytochemical and antioxidant characteristics of extracts from selected Lamiaceae species. Food Chem. 2005, 91, 525–533.
- Chkhikvishvili, I.; Sanikidze, T.; Gogia, N.; Mchedlishvili, T.; Enukidze, M.; Machavariani, M.; Vinokur, Y.; Rodov, V. Rosmarinic acid-rich extracts of summer savory (Satureja hortensis L.) protect Jurkat T cells against oxidative stress. Oxidative Med. Cell. Longev. 2013, 2013, 456253.
- Lung, I.; Soran, M.L.; Tudoran, C.; Măruţoiu, C. Effect of microwave irradiation on polyphenolic compounds from Satureja hortensis L. Cent. Eur. J. Chem. 2013, 11, 535–541.
- Mašković, P.; Veličković, V.; Mitić, M.; Đurović, S.; Zeković, Z.; Radojković, M.; Cvetanović, A.; Švarc-Gajić, J.; Vujić, J. Summer savory extracts prepared by novel extraction methods resulted in enhanced biological activity. Ind. Crops Prod. 2017, 109, 875–881.
- Boroja, T.; Katanić, G.; Rosić, D.; Selaković, J.; Joksimović, D.; Mišić, V.; Stanković, N.; Jovičić, V.; Mihailović, J. Summer savory (Satureja hortensis L.) extract: Phytochemical profile and modulation of cisplatin-induced liver, renal and testicular toxicity. Food Chem. Toxicol. 2018, 118, 252–263.
- Malankina, E.L.; Romanova, N.G.; Solopov, S.G.; Tkacheva, T.N. Garden savory—A promising spicy culture for the Moscow region. Kartof. I Ovoshi 2018, 11, 25–27.
- Plánder, S.; Gontaru, L.; Blazics, B.; Veres, K.; Kéry, Á.; Kareth, S.; Simándi, B. Major antioxidant constituents from Satureja hortensis L. extracts obtained with different solvents. Eur. J. Lipid Sci. Technol. 2012, 114, 772–779.
- Bros, I.; Soran, M.L.; Briciu, R.D.; Cobzac, S.C. HPTLC quantification of some flavonoids in extracts of Satureja hortensis L. obtained by use of different techniques. J. Planar Chromatogr. Mod. TLC 2009, 22, 25–28.
- Gupta, S.D.; Jatothu, B. Fundamentals and applications of light-emitting diodes LEDs in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220.
- Okamoto, K.; Yanagi, T.; Takita, S. Development of plant growth apparatus using blue and red LED as artificial light source. Acta Hortic. 1996, 440, 111–116.
- Jao, R.C.; Fang, W. Growth of potato plantlets in vitro is different when provided concurrent versus alternating blue and red light photoperiods. HortScience 2004, 39, 380–382.
- Jao, R.-C.; Lai, C.-C.; Fang, W.; Chang, S.-F. Effects of red light on the growth of Zantedschia plantlets in vitro and tuber formation using light-emitting diodes. HortScience 2005, 40, 436–438.
- Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.T.L.; Thu, N.T.; Hien, N.T.T.; Khanh, T.D.; Ha, C.H. LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes. Plants 2020, 9, 1344.
- Shulgina, A.A.; Kalashnikova, E.A.; Tarakanov, I.G.; Kirakosyan, R.N.; Cherednichenko, M.Y.; Polivanova, O.B.; Baranova, E.N.; Khaliluev, M.R. Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo. Horticulturae 2021, 7, 195.
- Zhang, X.H.; Zheng, X.T.; Sun, B.Y.; Peng, C.L.; Chow, W.S. Over-expression of the CHS gene enhances resistance of Arabidopsis leaves to high Light. Environ. Exp. Bot. 2018, 154, 33–43.
- Ma, Z.H.; Li, W.F.; Mao, J.; Li, W.; Zuo, C.W.; Zhao, X.; Dawuda, M.M.; Shi, X.Y.; Chen, B.H. Synthesis of light-inducible and light-independent anthocyanins regulated by specific genes in grape ‘Marselan’ (V. vinifera L.). PeerJ 2019, 7, e6521.
- Wang, W.; Su, M.; Li, H.; Zeng, B.; Chang, Q.; Lai, Z. Effects of supplemental lighting with different light qualities on growth and secondary metabolite content of Anoectochilus roxburghii. PeerJ 2018, 6, e5274.
- Liu, H.K.; Chen, Y.Y.; Hu, T.T.; Zhang, S.J.; Zhang, Y.H.; Zhao, T.Y.; Yu, H.E.; Kang, Y.F. The influence of light-emitting diodes on the phenolic compounds and antioxidant activities in pea sprouts. J. Funct. Foods 2016, 25, 459–465.
- Azad, M.O.K.; Kim, W.W.; Park, C.H.; Cho, D.H. Effect of Artificial LED light and far infrared irradiation on phenolic compound, isoflavones and antioxidant capacity in soybean (Glycine max L.) Sprout. Foods 2018, 7, 174.
- Wang, L.; Chen, Q.; Zhuang, S.; Wen, Y.; Cheng, W.; Zeng, Z.; Jiang, T.; Tang, C. Effect of Anoectochilus roxburghii flavonoids extract on H2O2—Induced oxidative stress in LO2 cells and D-gal induced aging mice model. J. Ethnopharmacol. 2020, 254, 112670.
- Pistelli, L.; Noccioli, C.; D’Angiolillo, F.; Pistelli, L. Composition of volatile in micropropagated and field grown aromatic plants from Tuscany Islands. Acta Biochim. Pol. 2013, 60, 43–50.
- Bharati, A.J.; Bansal, Y.K. In vitro production of flavonoids: A review. World J. Pharm. Pharm. Sci. 2014, 3, 508–533.
- Yeoman, M.M.; Yeoman, C.L. Manipulating secondary metabolism in cultured plant cells. New Phytol. 1996, 134, 553–569.
- Del Valle, J.C.; Buide, M.L.; Casimiro-Soriguer, I.; Whittall, J.B.; Narbona, E. On flavonoid accumulation in different plant parts: Variation patterns among individuals and populations in the shore campion (Silene littorea). Front. Plant Sci. 2015, 6, 939.
- Li, H.; Lin, Y.; Chen, X.; Bai, Y.; Wang, C.; Xu, X.; Wang, Y.; Lai, Z. Effects of blue light on flavonoid accumulation linked to the expression of miR393, miR394 and miR395 in longan embryogenic calli. PLoS ONE 2018, 13, e0191444.
- Zhao, D.; Li, M.; Xing, J.; Tong, Z. Effects of light on cell growth and flavonoids biosynthesis in callus cultures of Saussurea medusa Maxim. Acta Photophysiol. Sin. 1999, 25, 127–132.
- Deng, Y.; Yao, J.; Wang, X.; Guo, H.; Duan, D. Transcriptome sequencing and comparative analysis of Saccharina japonica (Laminariales, Phaeophyceae) under blue light induction. PLoS ONE 2012, 7, e39704.
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. Hortscience 2013, 48, 988–995.
More
Information
Subjects:
Plant Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
655
Revisions:
2 times
(View History)
Update Date:
01 Apr 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No