Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Janusz Blasiak | -- | 2162 | 2022-03-31 11:41:05 | | | |
| 2 | Dean Liu | -1 word(s) | 2161 | 2022-04-01 03:02:45 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Blasiak, J.; Chojnacki, J.; Pawlowska, E.; Jablkowska, A.; Chojnacki, C. Vitamin D May Protect against Breast Cancer. Encyclopedia. Available online: https://encyclopedia.pub/entry/21214 (accessed on 26 July 2026).
Blasiak J, Chojnacki J, Pawlowska E, Jablkowska A, Chojnacki C. Vitamin D May Protect against Breast Cancer. Encyclopedia. Available at: https://encyclopedia.pub/entry/21214. Accessed July 26, 2026.
Blasiak, Janusz, Jan Chojnacki, Elzbieta Pawlowska, Aleksandra Jablkowska, Cezary Chojnacki. "Vitamin D May Protect against Breast Cancer" Encyclopedia, https://encyclopedia.pub/entry/21214 (accessed July 26, 2026).
Blasiak, J., Chojnacki, J., Pawlowska, E., Jablkowska, A., & Chojnacki, C. (2022, March 31). Vitamin D May Protect against Breast Cancer. In Encyclopedia. https://encyclopedia.pub/entry/21214
Blasiak, Janusz, et al. "Vitamin D May Protect against Breast Cancer." Encyclopedia. Web. 31 March, 2022.
Copy Citation
In general, the protective effect of vitamin D3 against breast cancer is underlined by inhibition of proliferation and migration, stimulation of differentiation and apoptosis, and inhibition of epithelial/mesenchymal transition in breast cells. Vitamin D3 may also inhibit the transformation of normal mammary progenitors into breast cancer stem cells that initiate and sustain the growth of breast tumors. As long noncoding RNAs (lncRNAs) play an important role in breast cancer pathogenesis, and the specific mechanisms underlying this role are poorly understood, we provided several arguments that vitamin D3/VDR may induce protective effects in breast cancer through modulation of lncRNAs that are important for breast cancer pathogenesis.
breast cancer
vitamin D3
1,25(OH)2D
VDR
lncRNA
miRNA
1. Introduction
Despite significant recent progress in diagnosis and therapy, breast cancer is still a serious problem for individuals and societies (reviewed in [1]). There are several reasons for this, including heterogeneity of cases and poorly understood mechanisms of breast cancer pathogenesis. Although surgery, chemo- and radiotherapy, and hormone therapy are still the basic modes of breast cancer treatment, several chemical agents have been considered to assist breast cancer therapy and/or recommended in breast cancer prevention (reviewed in [2]). Some substances that can be administered with the diet, including vitamins, have been reported to improve the treatment of or play a preventive role in breast cancer ([3][4][5][6]). Several reports have presented vitamin D to have a beneficial potential in breast cancer (e.g., [7][8][9][10]).
Vitamin D3 (cholecalciferol), which is derived from animals, is the main type of vitamin D, and its biologically active form, 1α,25-dihydroxyvitamin D3 (1,25(OH)2D, calcitriol), is also an endocrine hormone. The serum concentration of the stable precursor of 1,25(OH)2D, 25-hydroxyvitamin D (25(OH)D, calcidiol) is usually used as a marker of vitamin D3 concentration in an organism. The biological activity of 1,25(OH)2D is underlined by its binding to the vitamin D3 receptor (VDR), a member of the nuclear receptor superfamily of ligand-inducible transcription factors. This 1,25(OH)2D/VDR complex is often associated with retinoid X receptor alpha (RXRA), and together, they bind the vitamin D response elements (VDREs) in the promoters of many genes to activate or repress their transcription [11]. Downstream targets of VDR are a myriad of genes, including those involved in genome maintenance, immune response, and cancer (reviewed in [12][13]).
Although data from epidemiological studies associating the levels of 25(OH)D with a better outcome of breast cancer are not fully consistent, most imply a positive correlation. A meta-analysis suggested that high vitamin D3 levels are weakly associated with a low breast cancer risk but strongly associated with the survival of breast cancer patients [14]. A more recent meta-analysis confirmed that lower blood 25(OH)D levels are associated with decreased survival in breast cancer patients [15]. Experimental studies have provided more convincing arguments for the positive role of vitamin D3 in breast cancer prevention and treatment. The BRCA1 DNA-repair-associated, breast cancer type 1 susceptibility (BRCA1) gene, which is important in the pathogenesis of a subset of breast cancer cases (reviewed in [16]), was transactivated by VDR-induced factors [17]. This study also showed the BRCA1-dependent antiproliferative action of 1,25(OH)2D in breast cancer cell lines. Several other studies have shown the important role of the vitamin D3/VDR axis in breast cancer pathogenesis (reviewed in [8]). The general conclusion from these studies is that VDR may act as a tumor suppressor whose regulation is impaired in breast cancer transformation [18]. However, the mechanism underlying this beneficial potential of vitamin D and its receptor is unknown.
Vitamin D metabolism and signaling in cancer are determined by various factors, including regulatory noncoding RNAs (ncRNAs) (reviewed in [19]). On the other hand, these RNAs can be regulated by vitamin D3/VDR signaling. Although most studies on these mutual interactions have focused on microRNAs (miRNAs), the role of long noncoding RNAs (lncRNAs) is emerging [19].
The basic functions of lncRNAs in the regulation of gene expression can be arbitrarily divided into antisense/signaling, guide, scaffold, and decoy. A lncRNA may pair with a complementary fragment of mRNA, preventing or inhibiting its translation. Transcription can be influenced by a lncRNA through recruitment and/or by guiding transcriptional activators and repressors to the target gene. A lncRNA may serve as a platform (scaffold) to facilitate the assembly of a chromatin remodeling complex to alter the structure of chromatin to be more permissive or repressive for transcription factors. A lncRNA may act as a decoy to recruit other regulatory RNAs or transcription factors and sequester them from their target mRNA or DNA. The “sponging” effect occurs when a single lncRNA binds many miRNAs, preventing interaction with their targets. Several lncRNAs have been identified to modulate VDR signaling in breast cancer [20][21].
Therefore, lncRNAs can be involved in vitamin D3/VDR signaling, and the vitamin may regulate the expression of lncRNAs that may be important in breast cancer. This regulatory circuit justifies studies on the role of lncRNAs as a mediator of the effects of vitamin D in breast cancer. This manuscript summarizes and updates information on the effects of vitamin D3/VDR in breast cancer, the mutual interaction between vitamin D3/VDR and lncRNAs, and the role lncRNAs may play in the effects of vitamin D in breast cancer pathogenesis. In the previous work, researchers reviewed the role of vitamin D in triple-negative and BRCA1-deficient breast cancer cases [8].
2. Vitamin D Signaling in Breast Cancer
Synthesis of vitamin D3 in the skin requires UV radiation, which can induce DNA damage, preferentially cyclobutane pyrimidine dimers and (6-4)-pyrimidine-pyrimidone photoproducts, which are found in many skin cancers, including melanoma (reviewed in [22]). On the other hand, vitamin D3 protects the skin against UV-induced aging and DNA damage causing cancer. This raises the question about the general anticancer properties of vitamin D3.
Usually, the biological effects of vitamin D are classified into nongenomic and genomic (reviewed in [23]). Genomic effects are mediated by VDR, RXR, and VDREs and result in long-term, sometimes delayed, biological consequences. The nongenomic pathway includes rapid cellular effects of 1,25(OH)2D, such as the protection of VDR-defective human fibroblasts against DNA damage induced by UV mediated by endoplasmic reticulum stress protein 57 (ERP57, MARRS) [24]. Therefore, the anticancer effects of vitamin D3 may be underlined by its genomic and nongenomic actions (reviewed in [25]).
The anticancer effect of vitamin D3 is not limited to UV-induced skin cancers. Colston et al. and Abe et al. were the first to show the anticancer properties of vitamin D in vitro [26][27]. Subsequently, many in vivo and in vitro studies showed the protective action of vitamin D against various cancers and identified several genes important in cancer transformation as targets in the genomic action of vitamin D (reviewed in [9]). In general, the antitumor effects of vitamin D are underlined by modulation of specific signaling pathways that are involved in tumor growth (Figure 1). Tumor growth is determined by the fate of cancer cells that may differentiate, arrest the cell cycle, and undergo apoptosis and/or degradation.
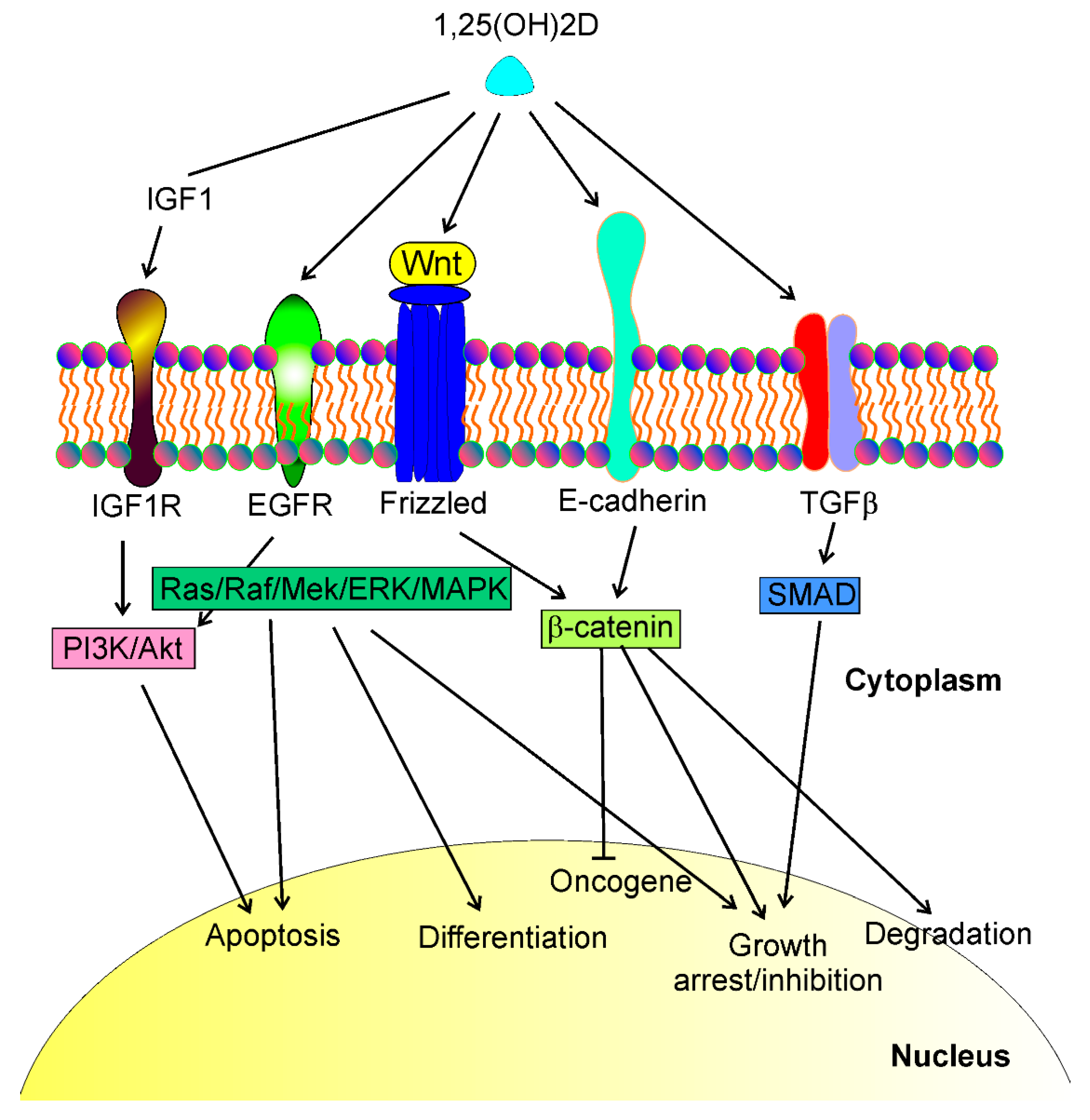
Figure 1. 1,25(OH)2D, a biologically active metabolite of vitamin D, may interact with different transmembrane proteins and affect several signaling pathways that regulate essential phenomena in cancer cells such as apoptosis, differentiation, growth arrest, cell cycle, oncogene and tumor suppressor expression, degradation. These proteins include, but are not limited to, insulin growth factor 1 (IGF1) receptor (IGF1R), epidermal growth factor receptor (EGFR), Wnt/frizzled proteins, E-cadherin, and transforming growth factor-beta (TGFβ). Interactions of 1,25(OH)2D with these proteins lead to a cascade of downstream events resulting in the activation of several signaling pathways, including PI3K/Akt, Ras/Raf/Mek/ERK/MAPK, β-catenin, and SMAD (phosphatidylinositol 3-kinase/AKT serine/threonine kinase, KRAS proto-oncogene/Raf-1 proto-oncogene/mitogen-activated protein kinase kinase 1, SMAD family members, respectively). These signaling pathways may directly regulate the life and death of cancer cells or control the transcription of genes whose products contribute to cancer transformation.
Vitamin D3/VDR signaling is implicated in the morphogenesis of the postnatal mammary gland, negatively regulating its growth [28]. Therefore, impaired vitamin D/VDR signaling may promote the abnormal development of the mammary gland, which may contribute to young age-onset breast cancer cases.
Malignant breast tumors are formed and maintained with the involvement of breast cancer stem cells (BCSCs) [29]. Moreover, BCSCs are involved in progression to metastasis, relapse, and therapy resistance. 1,25(OH)2D and a gemini analog of vitamin D3 (BXL0124) were reported to inhibit mammosphere formation from mammary progenitors and downregulated the pluripotency markers octamer-binding transcription factor 4 (OCT4) and Kruper like factor 4 (KLF-4) in mammospheres [30]. 1,25(OH)2D also repressed markers of stem cell-like phenotypes, including cluster of differentiation 44 (CD44), CD49f, cleaved Notch receptor 1 (c-Notch1), and phosphorylated nuclear factor kappa-light-chain-enhancer of activated B cells (pNFκB) in breast cancer. Subsequently, it was shown that 1,25(OH)2D and BXL0124 downregulated the pluripotency markers OCT4, CD44, and laminin subunit alpha 5 (LAMA5) in the mammospheres of a triple-negative breast cancer (TNBC) cell line [31]. Furthermore, 1,25(OH)2D and its analog repressed other proteins important for the maintenance of BCSCs, including Notch1-3, jagged canonical Notch ligand 1 (JAG1-2), Hes family BHLH transcription factor 1 (HES1), and NFκB. Therefore, vitamin D3/VDR signaling may prevent acquiring a stem-like phenotype by normal mammary gland progenitors and somatic breast cancer cells (Figure 2). Several pathways involved in cancer stemness are modulated by lncRNAs, and their targeting may be considered a new strategy to control tumor development, invasion, metastasis, and therapeutic resistance (e.g., [32][33][34]).
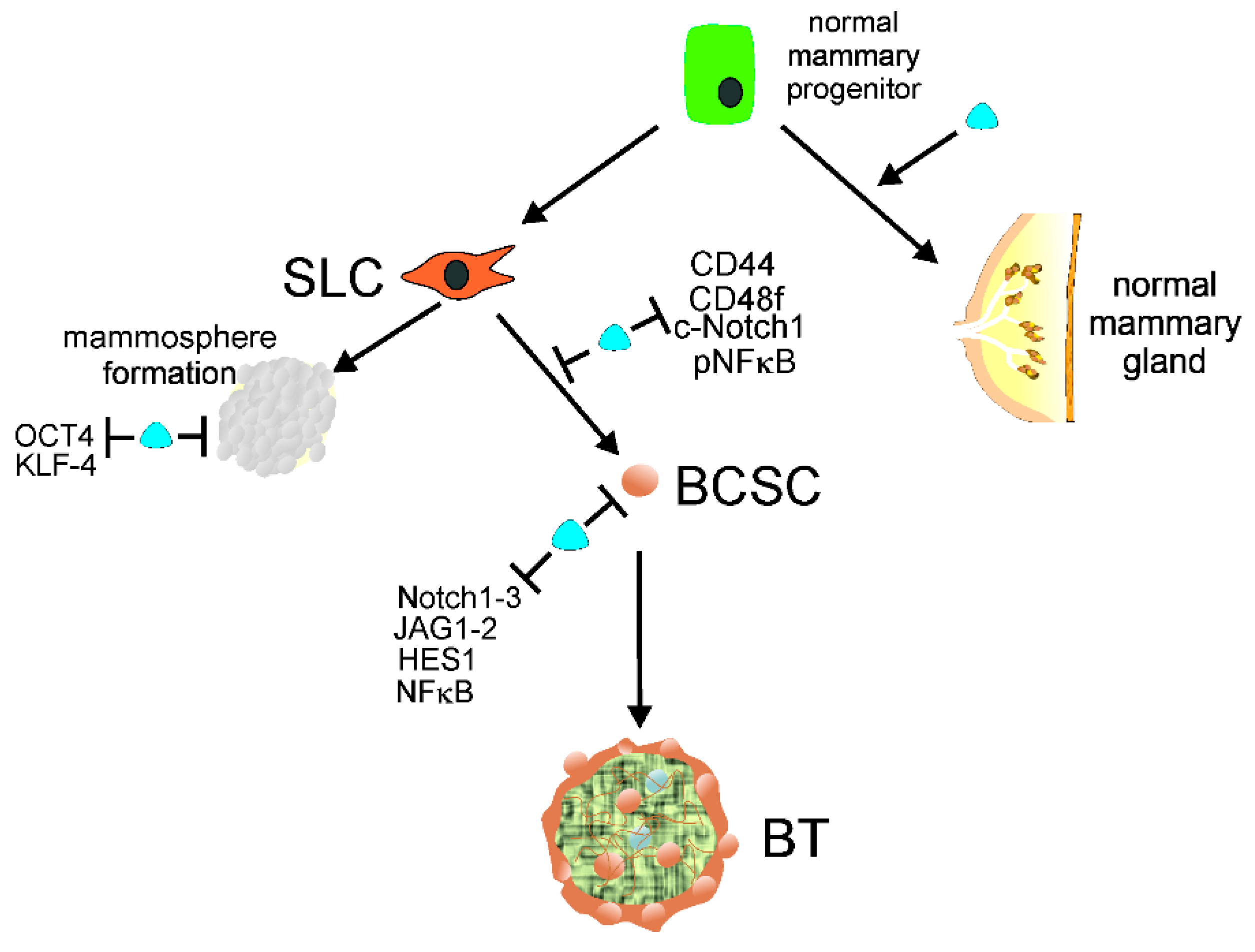
Figure 2. Normal mammary gland progenitors may develop into a normal mammary gland or acquire stem cell-like (SLC) properties to form mammospheres or/and transform into breast cancer stem cells (BCSCs). 1,25(OH)2D, a biologically active metabolite of vitamin D, along with its receptor, VDR, symbolized here as a blue tear, supports the normal development of mammary gland and prevents mammosphere formation by repressing the pluripotency markers OCT4 and KLF-4. Vitamin D may also prevent the conversion of SLC cells into BCSCs by downregulation of other pluripotency markers, including CD44, CD49f, c-Notch1, and pNFκB. Vitamin D may also repress proteins needed for the maintenance of BCSCs, such as Notch1-3, JAG1-2, HES1, and NFκB, preventing the initiation and formation of breast tumors (BTs). Abbreviations are defined in the main text.
The hormonal properties of vitamin D3, as part of the vitamin D3 endocrine system, may be responsible for the increased efficacy of tamoxifen, a selective inhibitor of estrogen receptor (ER) commonly applied in breast cancer therapy [35]. Such an action of vitamin D seems to be especially important in the case of breast cancer in postmenopausal women who no longer maintain ovarian production of estrogen [36]. Aromatase is a key enzyme in estrogen synthesis, and vitamin D was reported to inhibit it and repress the expression of estrogen receptor alpha (ERα) in breast tissue, [37][38][39][40][41]. Therefore, vitamin D is particularly important in breast cancer ER-positive cases in postmenopausal women. Many lncRNAs were reported to play a role in the regulation of estrogen-dependent signaling (reviewed in [42]).
The role of autophagy in cancer in general and in breast cancer in particular is emerging [43][44][45]. Regulation of autophagy is considered a method to prevent breast cancer and break therapeutic resistance (reviewed in [46]). It was shown that the vitamin D/VDR axis in breast cancer is regulated by many miRNAs [47]. Upregulation of miR-214 diminished VDR-mediated signaling in breast cancer cell lines with a concomitant positive correlation between VDR level and an inhibitor of the Hedgehog pathway [48]. The VDR gene has a sequence in its 3′-untranslated region recognized by miR-125b whose overexpression in the MCF-7-line decreases VDR level [49]. On the other hand, 1,25(OH)2D inhibited the expression of miR-125b in MCF-7 cells [50]. These results highlight the role of lncRNAs in mediating effects of vitamin D3 in breast cancer as one of the main mechanisms of biological action of lncRNAs is miRNAs sponging.
Other pathways and mechanisms may contribute to vitamin D3/VDR signaling in breast cancer (reviewed in [10]). The results of research on breast cancer cell lines should be interpreted considering the high heterogeneity of breast cancer cases and different forms of VDR, as well as pleiotropy of biological action of vitamin D [51]. Breast cancer cells, depending on origin, can be inherently vitamin D3 resistant or sensitive or acquire resistance during disease progression due to various effects, including methylation of the VDR promoter [52]. Heterogeneity of breast cancer also involves differences between specific breast compartments, which cause differential distribution of vitamin D3 in different regions of breast tumors due to different activity of enzymes involved in vitamin D3/VDR signaling.
In summary, several epidemiological studies show vitamin D3 deficiency in breast cancer patients. These association studies are supported by experimental research, but the mechanism underlying the potential beneficiary effect of vitamin D3 in breast cancer remains poorly understood. To apply vitamin D3 in breast cancer prevention and therapy effectively and safely, more controlled clinical trials and mechanistic laboratory studies are needed. 1,25(OH)2D may directly interact with several membrane-bound proteins to initiate a cascade of several signaling pathways that control cell growth and cell cycle arrest, cell migration, proliferation, and apoptosis important for cancer transformation.
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
01 Apr 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No