Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Arkasubhra Ghosh | -- | 5357 | 2022-03-31 11:16:22 | | | |
| 2 | Vivi Li | Meta information modification | 5357 | 2022-04-01 04:41:22 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ghosh, A.; , .; D’souza, S.; Deshpande, V. Ocular Surface Infection Mediated Molecular Stress Responses. Encyclopedia. Available online: https://encyclopedia.pub/entry/21209 (accessed on 25 June 2026).
Ghosh A, , D’souza S, Deshpande V. Ocular Surface Infection Mediated Molecular Stress Responses. Encyclopedia. Available at: https://encyclopedia.pub/entry/21209. Accessed June 25, 2026.
Ghosh, Arkasubhra, , Sharon D’souza, Vrushali Deshpande. "Ocular Surface Infection Mediated Molecular Stress Responses" Encyclopedia, https://encyclopedia.pub/entry/21209 (accessed June 25, 2026).
Ghosh, A., , ., D’souza, S., & Deshpande, V. (2022, March 31). Ocular Surface Infection Mediated Molecular Stress Responses. In Encyclopedia. https://encyclopedia.pub/entry/21209
Ghosh, Arkasubhra, et al. "Ocular Surface Infection Mediated Molecular Stress Responses." Encyclopedia. Web. 31 March, 2022.
Copy Citation
Infection mediated ocular surface stress responses are activated as early defense mechanisms in response to host cell damage. Integrated stress responses initiate the host response to different types of infections and modulate the transcription of key genes and translation of proteins. The crosstalk between host and pathogen results in profound alterations in cellular and molecular homeostasis triggering specific stress responses in the infected tissues. The amplitude and variations of such responses are partly responsible for the disease severity and clinical sequelae. Understanding the etiology and pathogenesis of ocular infections is important for early diagnosis and effective treatment.
host stress response
ocular surface infections
bacterial keratitis
fungal keratitis
viral keratitis
bacterial conjunctivitis
fungal conjunctivitis
viral conjunctivitis
proteins
gene expression
1. Introduction
The ocular surface of the human eye comprises the cornea, conjunctiva, lacrimal glands, and eyelids [1] and is inhabited by several microorganisms due to constant exposure to the external environment. The cornea is a transparent, avascular tissue that acts as a structural barrier, protects the eye against infections and contributes two-thirds of the refractive power of the eye [2]. The corneal microstructure consists of epithelial cells, keratocytes, endothelial cells and an extracellular matrix consisting primarily of acellular collagen and glycosaminoglycans. Microbial pathogens can infect various layers of the cornea which leads to keratitis [3]. The conjunctiva is a thin, transparent tissue layering the outer surface of the eyeball (bulbar conjunctiva) and extending onto the inner surface of the eyelids (forniceal and palpebral conjunctiva). It is composed of a surface layer of non-keratinizing, stratified, squamous epithelium overlying a vascular stroma composed of loose connective tissue [4]. Infection of the conjunctiva by various pathogens can result in conjunctivitis [5].
The ocular surface has commensal organisms but the presence of certain opportunistic pathogens can result in florid infections with serious clinical consequences [6]. Depending on the structure involved in the infection, the patient can present with corneal ulcers (keratitis) or conjunctivitis when it involves the cornea or conjunctiva, respectively. Corneal ulcers of infectious etiology are a major cause of visual impairment globally and can be caused by bacteria, fungi, viruses, or protists [7]. The incidences of bacterial, fungal, or other microbial infections on the ocular surface vary greatly in numbers globally. The severity of infections can range from mild and non-visually significant to severe sight threatening corneal ulcers requiring intensive topical and systemic therapy and even surgical intervention [8]. Gram-positive infections (commonly caused by Staphylococcus or Streptococcus bacteria) are classically well-localized ulcers with comparatively less local surrounding inflammation, although in some cases, they can result in large corneal ulcers that require therapeutic corneal transplantation. Gram-negative (most commonly Pseudomonas aeruginosa) infections are more aggressive with dense corneal cellularity and suppuration with prominent keratolysis within the infiltrate [9]. They are also associated with more clinical inflammation than a Gram-positive infection. Fungal keratitis is usually a more indolent infection and can have a relatively longer clinical course. Classic features such as the feathery margins, satellite lesions, and dry appearance of the ulcer can guide clinical suspicion and diagnosis [9]. However, since this classic clinical appearance may vary based on other factors such as the patient’s systemic health and prior medications used, additional diagnostic tests are typically required for accurate diagnosis.
Conjunctivitis is a common condition affecting the ocular surface which can be infective or non-infective. The most common organisms causing infective conjunctivitis are viruses (e.g., Adenovirus) and bacteria (e.g., Streptococcus pneumoniae, Haemophilus influenzae, etc.) [10]. Depending on the organism involved, there can be various levels of inflammation. The microbial etiological profile depends on the geography, specific risk factors, occupational and economic background of the population [8]. Since the economic burden of corneal infections is high and the access to health care is relatively limited in developing countries, effective management of microbial keratitis and conjunctivitis is important to prevent sight threatening complications [11].
Understanding the etiopathogenesis and associated clinical features of the most common ocular surface infections has helped unravel the complexities in diagnosis and subsequent treatment of infectious eye disease [11]. An accurate microbiological diagnosis helps enable the clinician to formulate an effective treatment regime, taking into account the specific antimicrobial sensitivity patterns of the pathogens where possible. Microbiological techniques such as microscopic analysis of sample smears and cultures to isolate the infective organism remain the most commonly used methods of diagnosis. Additional techniques such as in vivo confocal microscopy show characteristic features in cases of Acanthamoeba and fungal ulcers and can aid diagnosis. Even though these techniques provide an accurate diagnosis in many cases, there are still certain cases where diagnosing the causative organism and poor host response to the treatment can be a challenge [12]. Particularly, in cases of complex co-infections with multiple organisms, which may happen particularly in tropical regions, it is challenging for standard techniques to identify causation objectively. In most cases of refractory infections, a better understanding of the host tissue response is a critical aspect that can help tune the treatment to optimize the outcomes. Thus, there remains a clinical need for alternative approaches to diagnosis and monitoring which can give researchers more accurate and reliable results to design effective treatment strategies.
Molecular advancements have paved a way to diagnose even those cases where organisms are difficult to culture or visualize by standard diagnostic techniques. High-throughput techniques have aided in the discovery of previously unknown networks based on protein interactions and cellular changes on a global scale. The study of gene regulation and expression facilitates the understanding of normal, abnormal, or pathological cellular processes in the host which are functionally associated with disease. Additionally, gene expression studies proved to be very important in interpreting the contribution of the transcriptome to immune dysregulation [13] and immune response [14].
2. Clinical Presentation of Keratitis and Conjunctivitis and Treatment Modalities
The ocular surface includes the cornea, limbus, and the entire conjunctival surface from superior lid margin to inferior lid margin [1]. Different types of keratitis and conjunctivitis can result in significant ocular discomfort and in severe cases can also impact vision (Figure 1, Table 1).
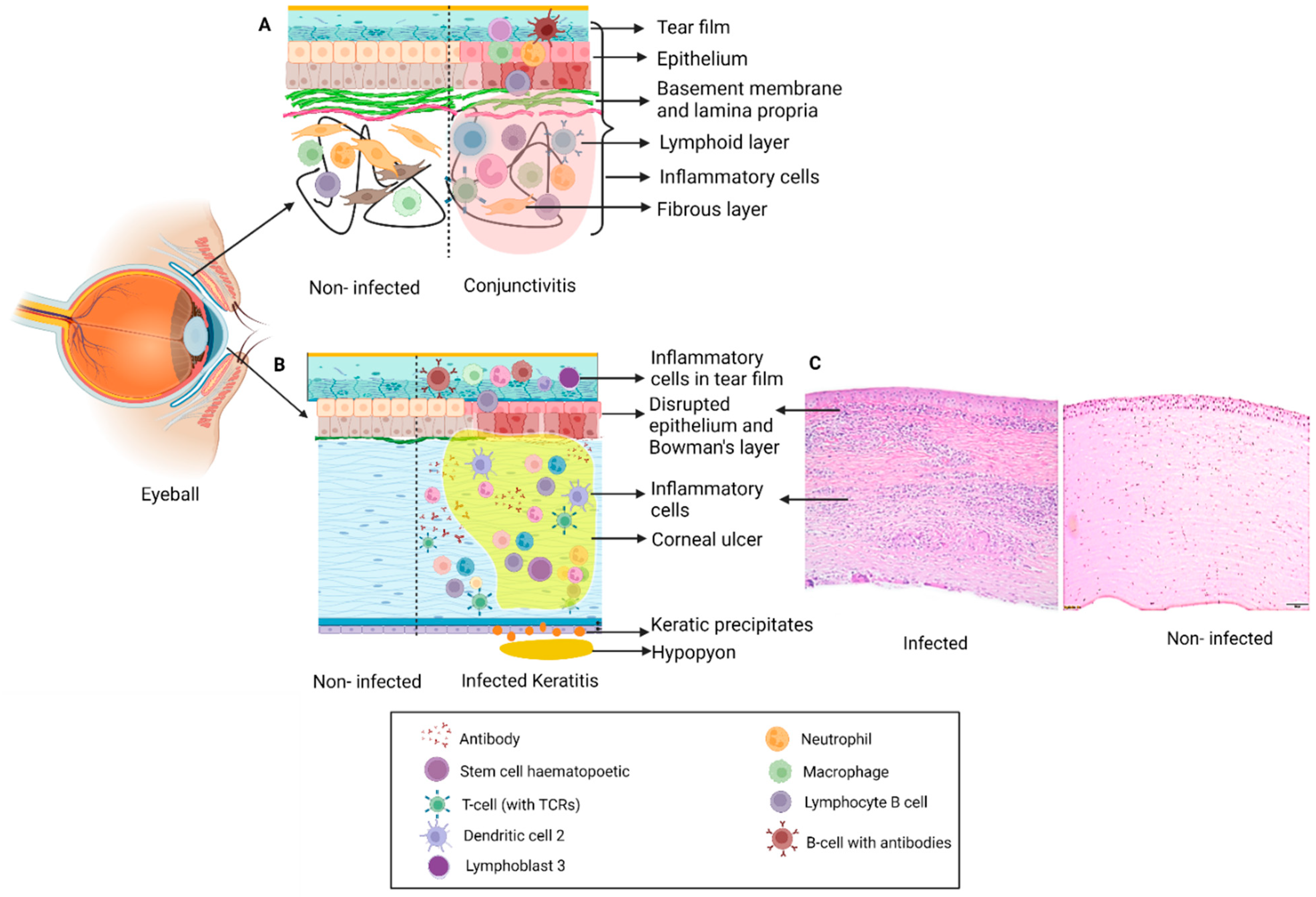
Figure 1. Schematic representation of changes in ocular tissue in: (A) Conjunctivitis: Inflammation of conjunctiva with infiltration of immune cells (neutrophils, lymphocytes, mast cells) which results in disruption in structure and inflammation leading to clinical symptoms and signs. (B) Keratitis: Infection of the cornea by different pathogens results in formation of a corneal ulcer or infiltrate with disruption in Bowman’s layer, epithelial and stromal edema with presence of inflammatory cells. Severe cases can also have inflammation in the anterior chamber in the form of hypopyon and keratic precipitates on the endothelium. (C) H&E staining of section on the right shows the histopathology of an infected cornea illustrating disruption of the Bowman’s layer and dense infiltration of the stroma by polymorphonuclear cells.
Table 1. Types of ocular infections and their pathophysiology.
| Ocular Infection |
Causative Agent | Sign/Symptoms | Treatment |
|---|---|---|---|
| Bacterial Keratitis |
|
Symptoms: Eye pain, blurred vision, photophobia, and discharge Signs: Corneal infiltrate of varying involvement of corneal stromal depth and diameter with overlying epithelial defect. Severe infections can be associated with anterior chamber inflammation and hypopyon formation. Progressive ulceration can result in complications including corneal perforation, scleritis, and endophthalmitis |
Topical antibiotics are used based on microbiological sensitivity. Combination of medications may be required |
| Fungal Keratitis |
|
Symptoms: Eye pain, blurred vision, photophobia, and discharge Signs: Dry looking base of corneal ulcer base with feathery margins. Can also have satellite lesions, endoexudates and hypopyon |
Topical antifungal medications as per severity of infections. Natamycin and voriconazole are the most common medications used. Oral antifungals of the azole group are also given for more severe infections |
| Viral Keratitis |
|
Symptoms: Discomfort, watering, itching, burning, and pain in the eye Signs: Would depend on the form of the herpes viral involvement |
Topical acyclovir and ganciclovir for epithelial forms of disease. Stromal viral keratitis requires topical steroids. Viral endothelitis and recurrent stromal involvement require oral antiviral medications |
| Bacterial Conjunctivitis |
|
Symptoms: Unilateral or bilateral redness, classically purulent discharge, photophobia, tearing, irritation, stinging, burning, and discomfort Signs: Diffuse bulbar conjunctival injection. Discharge is initially watery, becomes purulent as infection progresses |
Acute bacterial conjunctivitis is usually treated with topical fluoroquinolones, macrolides, and aminoglycosides. Tetracycline and macrolides are used to treat chlamydia |
| Trachoma | Chlamydia trachomatis serotypes A, B, Ba, C |
|
SAFE strategy: S: Surgery in case of trichiasis, A: Antibiotics (azithromycin and erythromycin) for C. trachomatis infection, F: Facial cleanliness, E: Environmental change for sanitation |
| Viral Conjunctivitis |
|
Symptoms: Unilateral or bilateral watering, redness, discomfort, and photophobia Signs: Eyelid edema, follicular conjunctivitis, tender preauricular lymphadenopathy. Chronic cases can have mild conjunctival scarring |
Symptomatic relief and cold compresses. Topical antibiotics to prevent secondary infection and topical steroids for severe inflammation in conjunctivitis |
3. Molecular Status of Ocular Surface Infection Mediated Host Molecular Responses
Upon infection, the ocular surface recognizes pathogens as foreign and eliminates them to maintain corneal transparency. The first line of defense includes a combination of mechanical and immunological factors which have evolved to protect the eye. Researchers discuss the roles of various pathogen mediated host molecular responses which may contribute to prevention of eye infection. The emergence of newer high-throughput techniques has revolutionized our ability to evaluate host protein responses on a global scale, facilitating the discovery of previously unexplored mechanisms [15].
3.1. Bacterial Keratitis
Proteomic technologies unraveled the mechanisms of bacterium–host interaction and improved understanding of the pathogenesis of bacterial keratitis. A proteomic study on an infection model for S. aureus in New Zealand white rabbits by Callegan et al. revealed α-toxin as the major virulence factor in keratitis as compared to β- and γ-toxins. The α-toxins cause host ocular damage by destroying the corneal epithelium while β-toxins were found to mediate keratitis and edema in the sclera as well as conjunctiva in this entry [16]. The proteomic analysis of keratitis caused by P. aeruginosa revealed multiple virulence factors such as elastase B (LasB), alkaline protease, exoenzyme S, slime polysaccharide, exotoxin A, endotoxin, leukocidin, phospholipase C, P. aeruginosa small protease (PASP), and protease IV along with cellular structures, such as pili and flagella [17]. SDS-PAGE and Western blot analyses were carried out to purify PASP and LasB and understand their role in bacterial keratitis [18]. The role of exotoxins (Exo S, Exo T, and Exo U) in P. aeruginosa keratitis was characterized by Western blot [18]. Exo S toxins are responsible for invasive infections and Exo U toxin leads to acute cytotoxicity in the host cells [19]. The host response against these microbial virulence factors in a protein array experiment revealed increased expression of IL-8, IL-6, and GRO in infected immortalized cell lines which showed an antibacterial effect [19]. Epithelial derived GRO primarily contributes to the recruitment of polymorphonuclear (PMS) leukocytes and secondarily it induces the corneal inflammation [20]. Global proteomic analysis by LC-MS/MS revealed 133 differentially expressed host proteins in both clinical (P. aeruginosa) and laboratory (P. aeruginosa ATCC 10145) infected strains and control samples. The upregulated proteins from the infected samples were related to pathogenicity and virulence. Two non-ribosomal peptide synthetases (NRPSs) were only present in the keratitis sample, which produced the secondary metabolite L-2-Amino-4-methoxy-trans-3-butenoic acid (AMB), regarded as a potent toxin secreted by P. aeruginosa [19][21]. The role of the nucleotide oligomerization domain (NOD)-like receptor (NLR) family with caspase activation and recruitment domain (CARD) containing 3 (NLRC3) was investigated in C57BL/6J mice after P. aeruginosa infection. Decreased levels of proinflammatory cytokines and activation of the NF-κB signaling pathway were observed when NLRC3 was overexpressed and reduced P. aeruginosa induced keratitis progression. The anti-inflammatory role of NLRC3 in P. aeruginosa induced keratitis suggested NLRC3 as a potential therapeutic target for PA induced keratitis [22]. Apart from the inflammatory proteins, the innate immune system of the host also plays an important role in protection from microbial pathogens. In the cornea, macrophages and dendritic cells play an important role in initiating the innate immune response. Recently, Hazlett et al. showed improved disease outcome after downregulation of one promising target, high mobility group box 1 (HMGB1), which promotes dendritic cell maturation, contributing to tissue pathogenesis and inflammation using small interfering RNA (siRNA) [23]. Activation of PRRs results in production of a cascade of inflammatory cytokines such as IL-1, IL-6, and IL-8 via NF-κB [24]. A myeloid differentiation primary response gene 88 (MyD88) mediated proinflammatory pathway is initiated once TLR4 and TLR5 on macrophages recognize the flagellin and lipopolysaccharide (LPS) of P. aeruginosa [25]. Pretreatment with flagellin has been shown to suppress early mucosal immune responses in mouse models of keratitis infection and leads to increased disease severity [26]. It has been reported that recruitment and persistence of PMN and other white blood cells are associated with corneal scarring [27]. The proteins and chemical compounds present in tear fluid, including iron, lactoferrin, peptidoglycan, phospholipase A2, defensins, and arachidonic acid metabolites [28], are associated with infections. The anti-inflammatory role of thrombomodulin in P. aeruginosa bacterial keratitis was explored and it was demonstrated that treatment with recombinant TM (rTM) results in protection against keratitis in B6 mice [28]. Increased expression of AnxA1 and fpr2, mediators in homeostasis of inflammation and ocular infections in infected mice, was observed and further investigations on the use of AnxA1 as a possible co-adjuvant therapeutic strategy in bacterial keratitis were suggested [29].
Significant alteration of gene expression in infected ocular surface tissues suggested that gene expression patterns and profiles are highly species specific. Studies have reported differentially expressed genes (DEGs) to distinguish bacterial and fungal keratitis. The Gene Expression Omnibus was used to download the expression profile of normal corneas and bacterial and fungal infection [30]. Amongst 451 DEGs found in bacterial keratitis, 148 DEGs were solely responsible for bacterial keratitis and 117 DEGs were co-expressed gene pairs in both fungal and bacterial keratitis. Three hundred and fifty-three specific DEGs were screened in fungal keratitis, among which 50 DEGs were strictly found in fungal keratitis and 87 DEGs were co-expressed gene pairs in both fungal and bacterial keratitis. In the fusional co-expression network by analyzing DEGs, nine biological pathways and seven KEGG pathways were revealed. Results from the network analyses indicated SOD2 DEG as the indicator for fungal keratitis and that DEG in TLR represented bacterial keratitis, representing genes for differential analysis. Although SOD2 does not have any descriptive role, it was shown to activate response to wounding and oxidation–reduction pathways [30]. A retrospective study was performed to evaluate contact lens keratitis by assessing the role of SNPs in IL-10 and IL-17 genes [31]. Buccal swab samples were collected from 88 keratitis patients, amongst them 25 were severe and 185 were healthy contact lens users, to carry out DNA extraction and SNP genotyping by pyrosequencing for IL-10 and IL-17 [31]. However, the SNPs did not show any relation to the severity of contact lens keratitis. SNPs in the minor allele G associated with IL-17 showed escalated risk of severe microbial keratitis. However, this did not conclude that severity of microbial keratitis is completely dependent on genetic variation in IL-17, although the IL-17 pathway is thought to be clinically important in the mechanism of microbial keratitis [31]. Microarray transcriptomic profiling of genes in bacterial keratitis and fungal keratitis patients revealed 185 unique differentially expressed genes in bacterial keratitis, 50 in fungal keratitis, and 339 common to both. In this entry, MMP9, along with other MMPs (MMP1, MMP7, MMP10, MMP12), pro-inflammatory cytokines (IL1B, TNF), and PRRs (TLR2, TLR4), were upregulated in bacterial and fungal keratitis. HIF1A and its induced genes were upregulated uniquely in bacterial keratitis [32]. Gowda et al. found constitutively high expression of Pglyrp-1 in the superficial cells of the corneal epithelium in mouse and human corneas infected with P. aeruginosa which suggested the protective role of Pglyrp-1 at the ocular surface. They showed that Pglyrp-1-/- mice challenged with P. aeruginosa keratitis showed poor bacterial clearance and resolution of keratitis [33], while disease severity was reduced with improved bacterial clearance in Pglyrp-2-/- mice that may be due to compensatory overexpression of defensins (mBD-2 and mBD-3), cathelicidin-related antimicrobial peptides (Cnlp), and Pglyrp-1 [33] (Table 2 and Table 3).
Table 2. Host response proteins in ocular microbial infections.
| Ocular Disease | Techniques Used | Host Molecular Responses | References |
|---|---|---|---|
| Bacterial keratitis |
|
|
|
| Fungal keratitis |
|
|
|
| Viral keratitis |
|
|
|
| Viral conjunctivitis |
ELISA | Elevated levels of hyaluronic acid (HA) can acts as a rapid diagnostic marker | Dreyfuss, J.L. et al Juliana (2015) |
Table 3. Host response genes in ocular microbial infections.
| Ocular Infection | Techniques | Host Molecular Responses | References |
|---|---|---|---|
| Viral Keratitis |
|
|
|
| Ebola virus uveitis | RNA sequencing, RT-qPCR | Type I interferon (IFN) | Smith, J. R. et al. (2017) |
| Bacterial keratitis |
|
|
|
| Fungal keratitis |
|
|
|
| Acanthameoba keratitis | MIP-2 and myeloperoxidase (MPO) assays | MIP-2 induces neutrophil infiltration which acts as a therapeutic strategy in Acanthameoba keratitis | Hurt, M. et al. (2001) |
3.2. Fungal Keratitis
Fungal keratitis (FK) is regarded as one of the most severe corneal infections. A tear proteome delivered comprehensive details regarding host ocular surface protein profile related to tissue injury and defense responses during FK. Differential expression of representative host response proteins causing FK can be implemented as biomarkers to establish the clinical prognosis and titrate the treatment and management strategy in distinct stages of FK [34]. The tear proteome of FK showed expression of a glutaredoxin-related (GRX) protein, which was secreted by Aspergillus sp. under oxidative stress. GRX is known to be involved in several cellular activities, i.e., protein folding, sulfur metabolism, protection of cells from oxidative stress, and DNA synthesis. Amongst the six abundant tear proteins released as host response, prolactin inducible protein and serum albumin were upregulated in the FK group. On the other hand, expressions of cystatin SN precursor, cystatin S precursor, lipocalin, and cystatin were downregulated [35]. Ananthi et al. further performed proteomic analysis in Fusarium keratitis infected eyes and control healthy subjects’ tears from different clinical stages. The tear groups of normal subjects, and early, intermediate, and late clinical stages Fusarium keratitis were pooled and analyzed. The authors performed two-dimensional difference gel electrophoresis (2D-DIGE) to assess the low-abundance proteins and improve protein separation. Liquid chromatography–tandem mass spectrometry (LC-MS/MS) was conducted for further protein identification and segmentation [36]. Different stages of Fusarium keratitis showed different expressions of host response proteins. As the disease progressed towards the late stages of FK, several proteins, i.e., α-1-antitrypsin, zinc-α-2-glycoprotein, haptoglobin α2 chain, albumin, apolipoprotein, lactoferrin, and haptoglobin precursor-β-chain were gradually upregulated. α-1-antitrypsin inactivates the microbial enzymes which act as an acute level reactant. Apolipoproteins are responsible for metabolism and uptake regulation of lipoproteins. The expression of lacritin precursor was downregulated to a negligible level in the early stage, compared to the control. During the later stage of Fusarium keratitis, the level of cystatin SA III and lipocalin was decreased [36]. Kandhavelu et al. segregated tear proteins by 1D-PAGE, glycosylation, and in-gel digestion to identify host response proteins by LC-MS/MS. The tear protein profiles of pooled tears from early Aspergillus keratitis and normal subjects were compared. The presence of proteins specific for neutrophil extracellular traps, proteins involved in wound healing, and complement system proteins was found only in the FK tears. Identification of host defense proteins and wound healing proteins at the early stages of Aspergillus keratitis may help in tracking Aspergillus keratitis progression [37]. Calvillo-Medina et al. analyzed the ability to form biofilms in vitro by F. falciforme isolated from FK corneal scrapes and examined its protein expression. They conducted 2D-PAGE separation for protein identification by MALDI-TOF. They found that 19 proteins were upregulated in biofilms, and amongst them six proteins showed unique expression. Relatively abundant proteins included enolase, ATP-citrate synthase, phosphoglycerate kinase, and transketolase. Some of these proteins were found to be associated with basal metabolism and act as potential virulence factors [38]. Mixed infection in microbial keratitis may occur from antibiosis of bacterial and fungal pathogens. As a result, proteomics analysis might explore the biology behind the mixed biofilm formation caused by bacteria and fungi.
Aspergillus flavus and Fusarium solani predominantly cause corneal mycotic ulceration in tropical countries [39]. Tear proteome profiles of Aspergillus keratitic patients were examined at different stages of infection. Profiling of the proteome was performed by 2D-PAGE and 2D-DIGE was carried out to quantify the protein levels. Upregulation of apolipoprotein, alpha-1-antitrypsin, lactoferrin, haptoglobin, and albumin was observed in the tear fluid of patients [40]. However, expressions of lacrimal lipocalin precursor, cystatin SA III precursor, zinc alpha-2 glycoprotein (ZAG), and lacritin precursor were downregulated. As the disease progressed from early to late stages, all the proteoforms of ZAG were concomitantly downregulated. There was no difference in the ZAG expression level in the keratitic tear film according to gender. Early events of host response showed upregulation of ZAG in Fusarium keratitis infection, highlighting its potential as a diagnostic biomarker. ZAG breaks down lipid in adipocytes but the exact role of ZAG in tears has not been studied intensively [40]. Biofilm-forming capability and antibiotic susceptibility of the ocular isolates of Candida albicans were studied and gene expression has been reported [41] in six keratitis and one orbital cellulitis clinical isolate. Biofilm formation was monitored by scanning electron microscopy (SEM) and confocal laser scanning microscopy (CLSM). Potential biofilm formation was observed in four ocular isolates along with resistance to three antifungal medications also described in one isolate. The rest of the isolates were susceptible to all the antifungal medications [41]. Two to three adherent layers of cells present at 24 h increased to multiple layers in 72 h according to the SEM studies. CLSM revealed that biofilm thickness increased to 17.98 μm at 72 h from 5.2 μm at 24 h [41]. Biofilm positive ocular and non-ocular C. albicans isolates showed upregulation of 27 genes, whose expression was similar in both non-ocular pathogenic C. albicans and biofilm-forming ocular isolates. These 27 genes were involved in the adhesion, initiation, maturation, and dispersal stages of biofilm. The expression pattern followed four different patterns of the biofilm-forming stages of the temporal expression in biofilm-positive ocular isolates. The similarity in gene expression between biofilm-forming ocular and non-ocular C. albicans isolates indicated that upregulated genes can be used as a possible therapeutic target. Transcriptome analysis of fungal keratitis revealed inflammatory cytokine genes, i.e., IL-1B, IL-6, TNF- α, to be significantly associated with fungal keratitis. Pathway enrichment analysis showed Wnt, cGMP–PKG, and Hippo signaling pathways to be responsible for the pathogenesis of fungal keratitis [42]. At the advanced stages of fungal keratitis, the levels of IL-1β, IL-6, IL-8, and IFN-γ in the aqueous humor were shown to be significantly increased [43]. Along with the cytokine expression, significantly higher expression of macrophage inducible Ca2+-dependent lectin receptor (Mincle) was observed during the early period of Aspergillus fumigatus infection in rats, which may play a role in the early host innate immune response of the corneal resistance against fungus [44] (Table 2 and Table 3).
3.3. Viral Infections
Proteomic profiling of corneal epithelial cells infected with HSV-1 was performed. Network analysis revealed the protein groups involved in mRNA splicing, ATP synthesis and post-translational protein folding, RNA processing, and gene expression [45][46].
Herpetic stromal keratitis (HSK) is caused by infection by Herpes Simplex virus (HSV) in the cornea. The interaction between cornea infiltrating inflammatory cells and resident cells in macrophages produces IL-1, TNF-α, and IFN-γ which in turn generate Th1 cells [47]. Corneal lesions and blindness are caused by a major influx of neutrophils and sustained local secretion of immune modulatory factors. IL-17 plays an important role in the massive infiltration of neutrophils into inflamed tissues [48]. Human corneal fibroblast (HCF) expresses IL-17R constitutively and HSK tissues express IL-17. Induction of IL-6 and IL-8 secretion by cultured HCF resulted in a synergistic effect between IL-17 and TNF- [48]. A strong chemotactic effect was observed in the neutrophils due to the secretion of IL-8 by HCF. IL-17 inhibited the secretion of RANTES; on the other hand, it induced the secretion of macrophage inflammatory proteins (MIPs) 1α and 3α. IFN-γ-related protein, i.e., IP10 and matrix metalloproteinase 1, levels were considerably elevated and the monocyte chemotactic protein 1 level remained unaltered. These data suggested that IL-17 might be an important factor in modulating the proinflammatory and neutrophil chemotactic factors in the corneal resident fibroblasts, resulting in the enhancement of the immunopathologic processes in human HSK [48]. The HSV-1 latency reactivation cycle gives rise to significant human pathology [49]. The HSV-1 latency associated transcript (LAT) regulates latency and reactivation by inhibiting apoptosis [50]. The Herpes Simplex virus (HSV-1) latency associated transcript (LAT) is associated with inhibiting apoptosis via hindering the activation of proapoptotic caspases. It was observed that LAT inhibited apoptosis by regulating the expression of apoptotic genes [50]. The molecular mechanism of antiapoptotic functions of LAT at a transcriptional level suggests that (i) LAT probably impedes apoptosis via upregulation of different components of the type I interferon (IFN) pathway; (ii) inhibition of apoptosis by LAT is neither accompanied by downregulation of Toll-like receptor (TLR) nor via caspase cascade at a transcriptional level. These factors suggested that immune exhaustion was not brought about by the antiapoptotic activity of the LAT [50]. miR-155 played a dominant role in HSK by regulating the immune system. HSV-1 infection of mouse cornea resulted in enhanced upregulation of miR-155 at 2, 7, and 15 days post-infection [51]. This upregulation was observed especially in activated CD4+ T cells, along with neutrophils in the infected cornea. On the contrary, the severity of the infection is reduced and accompanied by reduced angiogenesis and infiltration of CD4+ T cells, diminished Th1 and Th17 response in the infected cornea, and draining lymph nodes (DLNs) and lymphoid organs in miR-155 knockout mice [52]. The decreased proliferation of CD4+ T cells results in a decreased number of infiltrating CD4+ T cells, suggesting the role of miR-155 in promoting CD4+ T cell proliferation [52]. The in vivo silencing of miR-155 by injecting antigomir-155 nanoparticles in the conjunctiva diminished the severity of HSK with less infiltration of CD4+ T cells and neutrophils along with decreased production of proinflammatory cytokines, including IL-1, IL-6, IL-17, and IFN and chemokines, e.g., Ccl-2 and Cxcl-1 [52]. The antiangiogenic effect of antigomiR-132 accompanied an enhanced p120RasGAP expression and Ras activity was minimized in the endothelial cells of the cornea [53]. This concluded that p120RasGAP brings about the pro neovascularization function of miR-1320s, suggesting the knockdown of miR-132 can be a potential therapeutic alternative for HSK treatment [53]. The molecular basis of ocular HSV-1 infection has led to the identification of inhibitors of TANK-binding kinase 1 (TBK1) such as BX795, which strongly suppressed infection by multiple strains of HSV-1 in vivo [54]. The antiviral activity of BX795 targeted Akt phosphorylation in infected cells, resulting in the blockage of viral protein synthesis. It established the fact that BX795 can be used as a promising alternative broad-spectrum antiviral application in humans [54]. Recently, the effect of RNA interference in HSV keratitis was studied for prophylaxis and therapy by targeting glycoprotein D (gD) and glycoprotein E (gE). The expression of mRNA encoding gD and gE showed a decrease in the viral titer when used for prophylaxis rather than therapy. This result indicated the prophylactic role of small interfering RNA in HSV keratitis [55]. Yang et al. investigated the HSV-1 epithelial keratitis tear proteome by nano-LC/MS. Three hundred and twenty-six unique proteins were found in HSV keratitis samples. Functional annotation by gene ontology (GO) revealed most of the proteins are involved in antigen presentation, metabolic processes, and TNF mediated and T cell mediated inflammatory pathways. Levels of IL1A, IL12B, DEFB4A, and CAMP proteins were significantly higher in HSV keratitis samples, indicating higher levels of viral inhibition and inflammatory response. These unique proteins can be quantified by ELISA for the discovery of biomarkers for rapid diagnosis of HSV-1 epithelial keratitis [56]. HSK showed a significant decrease in MIP-1α deficient (−/−) hosts although virus replication and clearance did not differ significantly from that seen in infected wild-type (+/+) mice and it was concluded that MIP-1α is not needed to control virus growth in the cornea but is essential for the development of severe stromal keratitis [57]. The levels of Substance P in the infected corneas were evaluated by enzyme immunosorbent assay (EIA), which showed approximately a 15-fold higher amount of Substance P in the corneas with severe HSK lesions in comparison with those with mild HSK lesions, contributing to the clinical severity of HSK lesions in a mouse model [58]. Epithelial cells act as a critical barrier in protecting the cornea from microbial pathogen infection. The expression of TRIM32 was increased after infection with HSV-1 both in murine corneas and cultured human epithelial (HCE) cells. Furthermore, knockdown of the expression of TRIM32 significantly aggravated HSV-1 induced herpetic stromal keratitis (HSK) in mice and promoted the replication of HSV-1 in cultured HCE cells. The decreased expression of IFN-β and suppressed activation of interferon regulatory factor 3 (IRF3) positively regulated HSV-1 infection [45]. The induction of MIP-2 which in turn promoted the recruitment of neutrophils to the infected cornea was tested in a Chinese hamster model [59]. Hyaluronic acid (HA), which is an important component of the extracellular matrix, plays an important role in tissue development, cell migration, cell proliferation, and inflammation and was found to be elevated in tear fluid of patients with adenoviral conjunctivitis [60] (Table 2 and Table 3)
References
- Cher, I. Ocular surface concepts: Development and citation. Ocul. Surf. 2013, 12, 10–13.
- Srinivasan, M. Infective keratitis: A challenge to Indian ophthalmologists. Indian J. Ophthalmol. 2007, 55, 5–6.
- Marquart, M.E.; O’Callaghan, R.J. Infectious keratitis: Secreted bacterial proteins that mediate corneal damage. J. Ophthalmol. 2013, 2013, 369094.
- Sridhar, M.S. Anatomy of cornea and ocular surface. Indian J. Ophthalmol. 2018, 66, 190.
- Garfunkel, L.C.; Kaczorowski, J.; Christy, C. Pediatric Clinical Advisor E-book: Instant Diagnosis and Treatment; Elsevier Health Sciences: Philadelphia, PA, USA, 2007.
- St. Leger, A.J.; Caspi, R.R. Visions of eye commensals: The known and the unknown about how the microbiome affects eye disease. Bioessays 2018, 40, 1800046.
- Amescua, G.; Miller, D.; Alfonso, E. What is causing the corneal ulcer? Management strategies for unresponsive corneal ulceration. Eye 2012, 26, 228–236.
- Shah, A.; Sachdev, A.; Coggon, D.; Hossain, P. Geographic variations in microbial keratitis: An analysis of the peer-reviewed literature. Br. J. Ophthalmol. 2011, 95, 762–767.
- Garg, P.; Rao, G.N. Corneal ulcer: Diagnosis and management. Community Eye Health 1999, 12, 21.
- Azari, A.A.; Barney, N.P. Conjunctivitis: A systematic review of diagnosis and treatment. JAMA 2013, 310, 1721–1730.
- Prajna, V.N.; Nirmalan, P.K.; Saravanan, S.; Srinivasan, M. Economic analysis of corneal ulcers in South India. Cornea 2007, 26, 119–122.
- Go, E.P.; Wikoff, W.R.; Shen, Z.; O’Maille, G.; Morita, H.; Conrads, T.P.; Nordstrom, A.; Trauger, S.A.; Uritboonthai, W.; Lucas, D.A.; et al. Mass spectrometry reveals specific and global molecular transformations during viral infection. J. Proteome Res. 2006, 5, 2405–2416.
- Kosch, R.; Delarocque, J.; Claus, P.; Stefanie, C.; Jung, B.K. Gene expression profiles in neurological tissues during West Nile virus infection: A critical meta-analysis. BMC Genom. 2018, 19, 530.
- Mejias, A.; Dimo, B.; Suarez, N.M.; Garcia, C.; Suarez-Arrabal, M.C.; Jartti, T.; Blankenship, D.; Jordan-Villegas, A.; Ardura, M.I.; Xu, Z.; et al. Whole blood gene expression profiles to assess pathogenesis and disease severity in infants with respiratory syncytial virus infection. PLoS Med. 2013, 10, e1001549.
- Bartee, E.; McCormack, A.; Früh, K. Quantitative membrane proteomics reveals new cellular targets of viral immune modulators. PLoS Pathog. 2006, 2, e107.
- Callegan, M.C.; Engel, L.S.; Hill, J.M.; O’Callaghan, R.J. Corneal virulence of Staphylococcus aureus: Roles of alpha-toxin and protein A in pathogenesis. Infect. Immun. 1994, 62, 2478–2482.
- Strateva, T.; Mitov, I. Contribution of an arsenal of virulence factors to pathogenesis of Pseudomonas aeruginosa infections. Ann. Microbiol. 2011, 61, 717–732.
- Sewell, A.; Dunmire, J.; Wehmann, M.; Rowe, T.; Bouhenni, R. Proteomic analysis of keratitis-associated Pseudomonas aeruginosa. Mol. Vis. 2014, 20, 1182.
- Hilliam, Y.; Kaye, S.; Winstanley, C. Pseudomonas aeruginosa and microbial keratitis. J. Med. Microbiol. 2020, 69, 3–13.
- Sack, R.; Sathe, S.; Beaton, A.R.; McNamara, N.; Fleiszig, S.; Ni, M. Protein array characterization of bioactive proteins secreted by immortalized human corneal epithelium in response to pseudomonas constituents. Curr. Eye Res. 2009, 34, 92–98.
- Bouhenni, R.; Dunmire, J.; Rowe, T.; Bates, J. Proteomics in the study of bacterial keratitis. Proteomes 2015, 3, 496–511.
- Guo, L.; Kong, Q.; Dong, Z.; Dong, W.; Fu, X.; Su, L.; Tan, X. NLRC3 promotes host resistance against Pseudomonas aeruginosa-induced keratitis by promoting the degradation of IRAK1. Int. J. Mol. Med. 2017, 40, 898–906.
- Hazlett, L.D.; McClellan, S.; Somayajulu, M.; Bessert, D. Targeting Inflammation Driven by HMGB1 in Bacterial Keratitis—A Review. Pathogens 2021, 10, 1235.
- Taube, M.; Cendra, D.M.; Elsahn, A. Pattern recognition receptors in microbial keratitis. Eye 2015, 29, 1399–1415.
- Sun, Y.; Karmakar, M.; Roy, S.; Ramadan, R.T.; Williams, S.R.; Howell, S.; Shive, C.L.; Han, Y.; Stopford, C.M.; Rietsch, A.; et al. TLR4 and TLR5 on corneal macrophages regulate Pseudomonas aeruginosa keratitis by signaling through MyD88-dependent and-independent pathways. J. Immunol. 2010, 185, 4272–4283.
- Ross, B.X.; Gao, N.; Cui, X.; Standiford, T.J.; Xu, J.; Yu, F.X. IL-24 promotes Pseudomonas aeruginosa keratitis in C57BL/6 mouse corneas. J. Immunol. 2017, 198, 3536–3547.
- Willcox, M.D. Pseudomonas aeruginosa infection and inflammation during contact lens wear: A review. Optom. Vis. Sci. 2007, 84, 273–278.
- Sueke, H.; Kaye, S.B.; Neal, T.; Hall, A.; Tuft, S.; Parry, C.M. An in vitro investigation of synergy or antagonism between antimicrobial combinations against isolates from bacterial keratitis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4151–4155.
- Boyd, K.; Pagan-Duran, B.; Pink Eye (Conjunctivitis). American Academy of Ophthalmology. EyeSmart® Eye Health. Available online: https://www.aao.org/eye-health/diseases/pink-eye-conjunctivitis-list (accessed on 22 March 2019).
- Tian, R.; Zou, H.; Wang, L.; Liu, L.; Song, M.; Zhang, H. Analysis of differentially expressed genes in bacterial and fungal keratitis. Indian J. Ophthalmol. 2020, 68, 39–46.
- Carnt, N.A.; Cipriani, V.; Stapleton, F.J.; Calder, V.; Willcox, M.D. Association study of single nucleotide polymorphisms in IL-10 and IL-17 genes with the severity of microbial keratitis. Contact Lens Anterior Eye 2019, 42, 658–661.
- Chidambaram, J.D.; Kannambath, S.; Srikanthi, P.; Shah, M.; Lalitha, P.; Elakkiya, S.; Bauer, J.; Prajna, N.V.; Holland, M.J.; Burton, M.J. Persistence of innate immune pathways in late stage human bacterial and fungal keratitis: Results from a comparative transcriptome analysis. Front. Cell. Infect. Microbiol. 2017, 7, 193.
- Gowda, R.N.; Redfern, R.; Frikeche, J.; Pinglay, S.; Foster, J.W. Functions of peptidoglycan recognition proteins (Pglyrps) at the ocular surface: Bacterial keratitis in gene-targeted mice deficient in Pglyrp-2,-3 and-4. PLoS ONE 2015, 10, e0137129.
- Kuo, M.-T.; Chen, J.L.; Hsu, S.L.; Chen, A.; You, H.L. An omics approach to diagnosing or investigating fungal keratitis. Int. J. Mol. Sci. 2019, 20, 3631.
- Ananthi, S.; Chitra, T.; Bini, R.; Prajna, N.V.; Lalitha, P.; Dharmalingam, K. Comparative analysis of the tear protein profile in mycotic keratitis patients. Mol. Vis. 2008, 14, 500–507.
- Ananthi, S.; Venkatesh, P.N.; Lalitha, P.; Valarnila, M.; Dharmalingam, K. Pathogen induced changes in the protein profile of human tears from Fusarium keratitis patients. PLoS ONE 2013, 8, e53018.
- Kandhavelu, J.; Demonte, N.L.; Namperumalsamy, V.P.; Prajna, L.; Thangavel, C.; Jayapal, J.M.; Kuppamuthu, D. Aspergillus flavus induced alterations in tear protein profile reveal pathogen-induced host response to fungal infection. J. Proteom. 2017, 152, 13–21.
- Calvillo-Medina, R.P.; Reyes-Grajeda, J.P.; Barba-Escoto, L.; Bautista-Hernandez, L.A.; Campos-Guillén, J.; Jones, G.H.; Bautista-de Lucio, V.M. Proteome analysis of biofilm produced by a Fusarium falciforme keratitis infectious agent. Microb. Pathog. 2019, 130, 232–241.
- Maharana, P.K.; Sharma, N.; Nagpal, R.; Jhanji, V.; Das, S.; Vajpayee, R.B. Recent advances in diagnosis and management of Mycotic Keratitis. Indian J. Ophthalmol. 2016, 64, 346.
- Parthiban, N.; Sampath, N.L.; Jeya Maheshwari, J.; Prajna, N.V.; Lalitha, P.; Dharmalingam, K. Quantitative profiling of tear proteome reveals down regulation of zinc alpha-2 glycoprotein in Aspergillus flavus keratitis patients. Exp. Eye Res. 2019, 186, 107700.
- Ranjith, K.; Kalyana, C.S.; Adicherla, H.; Sharma, S.; Shivaji, S. Temporal expression of genes in biofilm-forming ocular Candida albicans isolated from patients with keratitis and orbital cellulitis. Investig. Ophthalmol. Vis. Sci. 2018, 59, 528–538.
- Zhang, Q.; Zhang, J.; Gong, M.; Pan, R.; Liu, Y.; Tao, L.; He, K. Transcriptome analysis of the gene expression profiles associated with fungal keratitis in mice based on RNA-Seq. Investig. Ophthalmol. Vis. Sci. 2020, 61, 32.
- Zhang, Y.; Liang, Q.; Liu, Y.; Pan, Z.; Baudouin, C.; Labbé, A.; Lu, Q. Expression of cytokines in aqueous humor from fungal keratitis patients. BMC Ophthalmol. 2018, 18, 105.
- Zhao, G.; Xu, Q.; Lin, J.; Chen, W.; Cui, T.; Hu, L.; Jiang, N. The role of Mincle in innate immune to fungal keratitis. J. Infect. Dev. Ctries. 2017, 11, 89–97.
- Cui, H.; Liu, Y.; Huang, Y. Roles of TRIM32 in corneal epithelial cells after infection with herpes simplex virus. Cell. Physiol. Biochem. 2017, 43, 801–811.
- Berard, A.R.; Coombs, K.M.; Severini, A. Quantification of the host response proteome after herpes simplex virus type 1 infection. J. Proteome Res. 2015, 14, 2121–2142.
- Wang, L.; Wang, R.; Xu, C.; Zhou, H. Pathogenesis of herpes stromal keratitis: Immune inflammatory response mediated by inflammatory regulators. Front. Immunol. 2020, 11, 766.
- Maertzdorf, J.; Osterhaus, A.D.; Verjans, G.M. IL-17 expression in human herpetic stromal keratitis: Modulatory effects on chemokine production by corneal fibroblasts. J. Immunol. 2002, 169, 5897–5903.
- Nicoll, M.P.; Proença, J.T.; Efstathiou, S. The molecular basis of herpes simplex virus latency. FEMS Microbiol. Rev. 2012, 36, 684–705.
- Tormanen, K.; Allen, S.; Mott, K.R.; Ghiasi, H. The latency-associated transcript inhibits apoptosis via downregulation of components of the type I interferon pathway during latent herpes simplex virus 1 ocular infection. J. Virol. 2019, 93, e00103-19.
- Banerjee, A.; Schambach, F.; DeJong, C.S.; Hammond, S.M.; Reiner, S.L. Micro-RNA-155 inhibits IFN-γ signaling in CD4+ T cells. Eur. J. Immunol. 2010, 40, 225–231.
- Huffaker, T.B.; Hu, R.; Runtsch, M.C.; Bake, E.; Chen, X.; Zhao, J.; Round, J.L.; Baltimore, D.; O’Connell, R.M. Epistasis between microRNAs 155 and 146a during T cell-mediated antitumor immunity. Cell Rep. 2012, 2, 1697–1709.
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Murphy, E.A.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E.; et al. MicroRNA-132–mediated loss of p120RasGAP activates the endothelium to facilitate pathological angiogenesis. Nat. Med. 2010, 16, 909–914.
- Jaishankar, D.; Yakoub, A.M.; Yadavalli, T.; Agelidis, A.; Thakkar, N.; Hadigal, S.; Ames, J.; Shukla, D. An off-target effect of BX795 blocks herpes simplex virus type 1 infection of the eye. Sci. Transl. Med. 2018, 10, eaan5861.
- Chen, L.; Pan, Z.-Q.; Zhai, C.-B. Adenovirus-mediated RNA interference against herpes simplex virus infection in vitro. Folia Histochem. Cytobiol. 2021, 59, 302–310.
- Yang, H.; Yang, X.; Wang, Y.; Zheng, X.; Zhang, Y.; Shao, Y. Comparative analysis of the tear protein profile in herpes simplex virus type 1 epithelial keratitis. BMC Ophthalmol. 2020, 20, 355.
- Tumpey, T.M.; Cheng, H.; Cook, D.N.; Smithies, O.; Oakes, J.E.; Lausch, R.N. Absence of macrophage inflammatory protein-1α prevents the development of blinding herpes stromal keratitis. J. Virol. 1998, 72, 3705–3710.
- Twardy, B.S.; Channappanavar, R.; Suvas, S. Substance P in the corneal stroma regulates the severity of herpetic stromal keratitis lesions. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8604–8613.
- Hurt, M.; Apte, S.; Leher, H.; Howard, K.; Niederkorn, J.; Alizadeh, H. Exacerbation of Acanthamoeba keratitis in animals treated with anti-macrophage inflammatory protein 2 or antineutrophil antibodies. Infect. Immun. 2001, 69, 2988–2995.
- Dreyfuss, J.L.; Regatieri, C.V.; Coelho, B.; Barbosa, J.B.; De Freitas, D.; Nader, H.B.; Martins, J.R. Altered hyaluronic acid content in tear fluid of patients with adenoviral conjunctivitis. An. Acad. Bras. Ciências 2015, 87, 455–462.
More
Information
Subjects:
Others
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
598
Revisions:
2 times
(View History)
Update Date:
01 Apr 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No