 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Cheikh Tidiane DIAGNE | + 1871 word(s) | 1871 | 2020-09-16 10:44:17 | | | |
| 2 | Vicky Zhou | Meta information modification | 1871 | 2020-09-23 08:16:06 | | | | |
| 3 | Vicky Zhou | -27 word(s) | 1844 | 2020-09-23 10:50:03 | | | | |
| 4 | Vicky Zhou | -1 word(s) | 1843 | 2020-10-27 08:48:51 | | |
Video Upload Options
Mayaro virus (MAYV), isolated for the first time in Trinidad and Tobago, has captured the attention of public health authorities worldwide following recent outbreaks in the Americas. It has a propensity to be exported outside its original geographical range, because of the vast distribution of its vectors. Moreover, most of the world population is immunologically naïve with respect to infection with MAYV which makes this virus a true threat. The recent invasion of several countries by Aedes albopictus underscores the risk of potential urban transmission of MAYV in both tropical and temperate regions. In humans, the clinical manifestations of MAYV disease range from mild fever, rash, and joint pain to arthralgia. In the absence of a licensed vaccine and clinically proven therapeutics against Mayaro fever, prevention focuses mainly on household mosquito control. However, as demonstrated for other arboviruses, mosquito control is rather inecient for outbreak management and alternative approaches to contain the spread of MAYV are therefore necessary. Despite its strong epidemic potential, little is currently known about MAYV.
1. Introduction
Mayaro virus (MAYV) is a single-stranded RNA virus. It belongs to the Togaviridae family and was first detected in 1954 in Trinidad in the sera of forest workers [1][2][3]. Like other alphaviruses, MAYV can infect, replicate and disseminate in both vertebrate and invertebrate hosts [4][5][6]. In humans, the virus causes Mayaro fever, which is characterized by long-lasting arthralgia similar to that occurring in Dengue fever. Although mortality among infected people is low as yet, Mayaro fever may become a major public health problem, particularly in rural areas, with increasing prevalence in the Amazon region due to ecosystem changes [7]. Although several outbreaks of Mayaro fever have already been reported more than three decades ago in Northern Brazil [8], MAYV is now spreading rapidly to other regions in Latin America [9]. It is likely that the global burden of Mayaro fever is still largely underestimated because of the lack of adequate and accurate diagnostics. At the same time, the Aedes mosquito, which can spread MAYV and other viruses, is invading new habitats and regions at an alarming pace [10]. Perhaps the most significant threat of a potential MAYV epidemic comes from increased colonization of urbanized areas by Aedes aegypti. In order to better predict the capacity of the virus to spread and infect new areas, further studies are urgently needed. At present, it cannot be excluded that MAYV infections, like those of ZIKV and Chikungunya (CHIKV) viruses, will one day become a major health issue. At present, MAYV is still neglected and few studies have been carried out on its pathogenesis, the biology of its potential vectors or the dynamics of its transmission.
2. Clinical Manifestations and Pathogenesis of Mayaro Fever in Humans
Phylogenetically, MAYV is closely related to CHIKV and, like the latter, causes a debilitating flu-like illness in the infected host that is indistinguishable from Chikungunya fever. The main symptoms include chills, fever, gastrointestinal manifestations, eye pain, myalgia and arthralgia [11][12]. In particular, arthralgia can last for months to years, making the Mayaro fever even more debilitating than flavivirus infections, whose symptoms last only one to three weeks [1][8][13][14][15]. Acute symptoms generally last three to five days in most patients with Mayaro disease [13][16]. Arthralgia and myalgia account for 50–89% and 75% of infected patients, respectively [17]. Dizziness and itching are the other clinical manifestations of the disease [18][19].
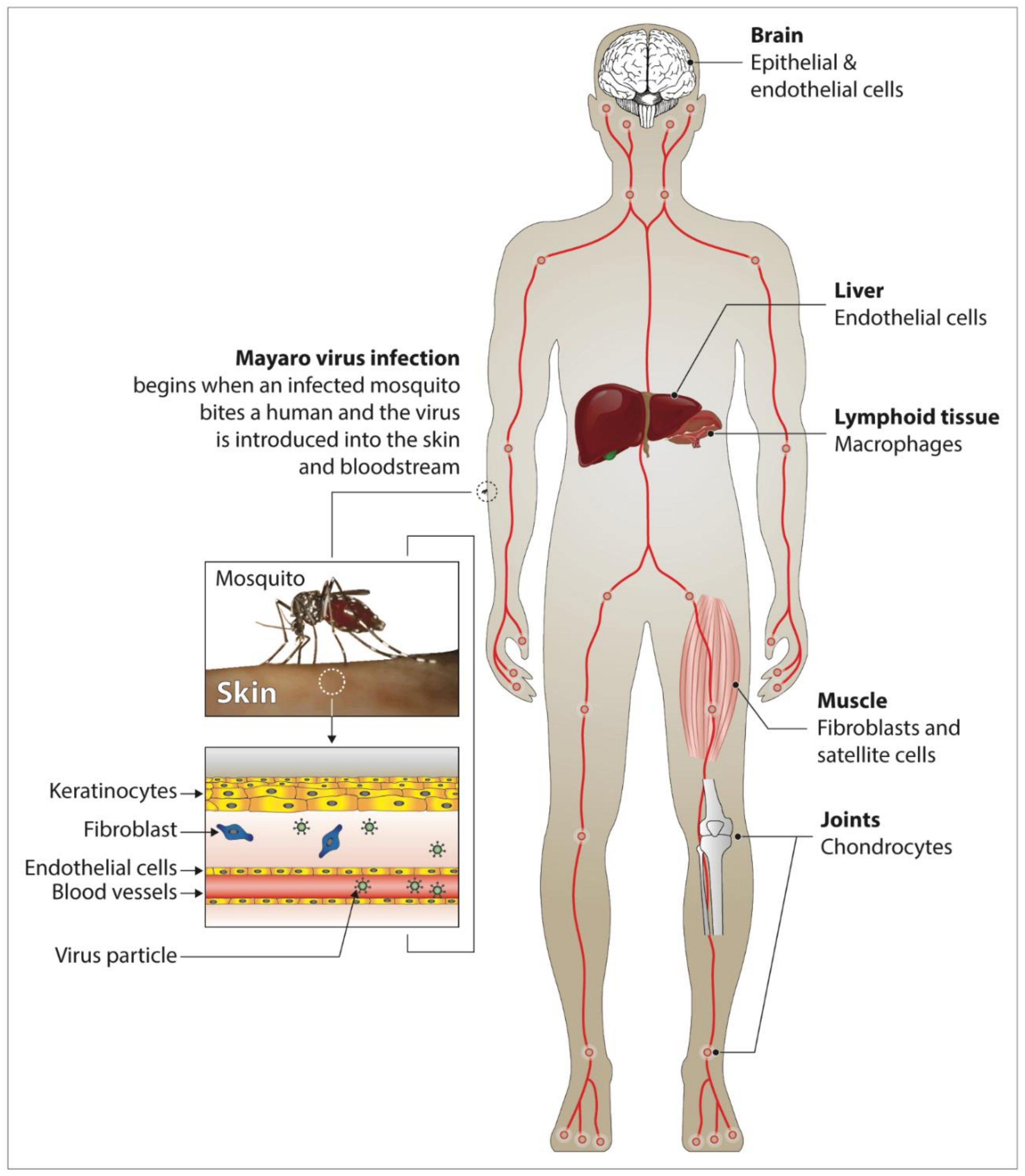
Severe complications can occur due to MAYV infection, among which are myocarditis, hemorrhagic and neurological manifestations [20]. After a bite from an infected vector, MAYV spreads via the blood vessels into the body of the susceptible host (Figure 1). The virus replicates in white blood cells (e.g., monocytes, macrophages) and spreads to bones, muscles and joints via the main sites of replication, the spleen and the liver [13][21][22][23][24][25][26]. The severity of Mayaro fever is associated with the production of pro-inflammatory cytokines and mediators (MCP-1, IL-2, IL-9, IL-13, IL-7, VEGF, IL-17, and IP-10) both in humans and experimental mouse models [27][28][29]. Some of these cytokines have been shown to be associated with other pathologies involving bones and joints [25][30][31].
Mayaro fever can also induce oxidative stress (OS) in the liver of infected mice and in vitro infected HepG2 cells [32][33]. The induction of OS has also been observed in several other arbovirus infections, including Chikungunya [34][35][36]. The latter process is likely to play a role in the pathogenesis of MAYV, because of its important role in the initiation and control of the production of many soluble mediators such as reactive oxygen species (ROS), and is furthermore involved in apoptosis and inflammation [37]. Currently, little is known about the beneficial or detrimental effects of this process on the infected host. Many cell types have been shown to be implicated in the pathogenesis of MAYV.
The role of macrophages, being targets for MAYV infection, in the development of arthritis has been demonstrated through the secretion of TNF-α and ROS [38]. In this respect, it is of interest to note that high serum levels of TNF-α have previously been reported in MAYV-infected patients and mice [29][38]. A recent study has also shown that in vitro infection of bone marrow-derived macrophages with MAYV results in the overexpression of key inflammasome proteins such as NLRP3, AIM2, ASC and caspase 1 [39]. Interestingly, the authors demonstrated that the induction of ROS was linked to the activation of the NLRP3 pathway. Because many studies have shown that ROS production can be associated with immune regulation [40], it would be of interest to investigate whether ROS secretion impacts the polarization of macrophages during MAYV infection. Results from studies using experimental mouse models and human cell lines have revealed that osteoblasts are susceptible to infection with alphaviruses, resulting in the secretion of MCP-1, IL-6, and IL-1β [21][41][42].
Primary human chondrocytes, osteoblasts and synoviocytes, which are the main cell types involved in arthralgia, are permissive to MAYV infection, indicating that these cells may be involved in the pathogenesis of the disease, notably as a result of the overexpression of arthritis-related genes [43]. Infection of the host by MAYV results in the sensitization of monocytes and the induction of their osteoclastogenic activity which leads, in turn, to bone erosion and cartilage damage. Taking into consideration the debilitating condition of MAYV-infected patients, which can last for months to years, further studies should be carried out to determine the mechanisms leading to severe arthritis as a result of MAYV infection. This also begs the question as to why some patients develop arthritis and others do not.
It would also be interesting to develop studies leading to the identification of biomarkers or risk factors associated with severe forms of the disease. There is considerable variation in antibody production and persistence from one host to another. MAYV infection induces a transient production of immunoglobulin M (IgM) antibodies, indicating the occurrence of recent infection that generally lasts for at least three months after the onset of clinical symptoms [12][44][45]. In addition, the presence of virus-specific immunoglobulin G (IgG) serum antibodies that persists throughout the lifespan of an infected person is an indicator of prior infection with MAYV, in particular when present at increased levels [12][44][45]. Interestingly, a study by Santiago et al. shows that neutralizing antibodies alone are not sufficient to prevent the occurrence of chronic arthritis [28]. Earnest and collaborators generated a series of neutralizing broad-spectrum mouse antibodies by the use of recombinant MAYV E2 protein [46] and showed their therapeutic utility against MAYV, thereby underscoring the importance that should be given to IgG subclasses and their effector functions.
3. Conclusions
Variabilities in susceptibility and ability to disseminate and transmit MAYV have been reported depending on the extrinsic incubation conditions, the virus strains used and the mosquito species tested. These variations in the expression of oral receptivity have been repeatedly described in natural populations of vector mosquitoes. Several factors, both intrinsic and extrinsic to the vectors, may explain these variations. Among these, possible interactions between the virus and the microbiota of the mosquito’s midgut seem to be particularly important, knowing that this symbiosis could provide important physiological functions to the host, including the synthesis of essential nutrients, resistance to infection and stimulation of the immune system. In many studies on the involvement of the immune system, bacteria have been used, but it was only very recently that their role in the immune response against virus infections was discovered. Differences in the expression of genes involved in the immune response are probably due to changes in the specific composition of the bacterial flora present in the midgut of the host mosquito and are important factors influencing the variation in transmission. To date, no data on potential vectors are available on this aspect.
Therefore, for further analysis, studies should be carried out on the diversity and relative proportions of the different taxa of this midgut microbiota. Given that virus–host interactions are often accompanied by variations in viral proteins, resulting in the emergence of disease, it would be interesting to investigate the contribution of recombination phenomena to the natural evolution of MAYV strains and the epidemiological risk that recombinant strains may present. From a technical point of view, the identification and subsequent inactivation, by homologous recombination, of genes involved in the replication or adsorption of viral particles, for example, could significantly reduce transmission. It would also be helpful to improve our knowledge of the mechanisms underlying MAYV replication in mosquitos. In this respect, further studies providing valuable information on the basic ecology and spatiotemporal dynamics of vector abundance and their association with MAYV should be carried out.
Transmission probability models could be used to estimate the natural periodicity of virus incidence cycles in each potential host. These predictive models will certainly make it possible to improve monitoring methods, thereby providing predictive capacity for the identification and early warning of emergence risks, as well as their control, by appropriate and accepted preventive methods. From the same perspective, two axes should be explored: (i) understanding how human contact with MAYV via vectors may increase with intensive use of forest resources; (ii) understanding how wildlife populations and communities, acting as reservoirs and potential vectors, respond to changes in the environment. These modifications will have an impact on wildlife populations, leading to the displacement of native forest species and the introduction of new ones capable of colonizing disturbed areas, resulting in the emergence of arboviruses, including MAYV, which were previously confined to the biotopes of natural reservoirs, thereby increasing the risk of contact with new mammalian and human hosts.
References
- Anderson, C.R.; Downs, W.G.; Wattley, G.H.; Ahin, N.W.; Reese, A.A. Mayaro Virus: A New Human Disease Agent. Am. J. Trop. Med. Hyg. 1957, 6, 1012–1016.
- LeDuc, J.W.; Pinheiro, F.P.; da Rosa, A.P.A.T. An Outbreak of Mayaro Virus Disease in Belterra, Brazil II. Epidemiology. Am. J. Trop. Med. Hyg. 1981, 30, 682–688.
- Schaeffer, M.; Gajdusek, D.C.; Lema, A.B.; Eichenwald, H. Epidemic Jungle Fevers Among Okinawan Colonists in the Bolivian Rain Forest. Am. J. Trop. Med. Hyg. 1959, 8, 372–396.
- Carvalho, C.A.M.; Silva, J.L.; Oliveira, A.C.; Gomes, A.M.O. On the entry of an emerging arbovirus into host cells: Mayaro virus takes the highway to the cytoplasm through fusion with early endosomes and caveolae-derived vesicles. PeerJ 2017, 5, e3245.
- Mezencio, J.M.S.; de Souza, W.; Fonseca, M.E.F.; Rebello, M.A. Ultrastructural study of Mayaro virus replication in BHK-21 cells. Arch. Virol. 1990, 114, 229–235.
- Mezencio, J.M.S.; de Souza, W.; Fonseca, M.E.F.; Rebello, M.A. Replication of Mayaro virus in Aedes albopictus cells: An electron microscopic study. Arch. Virol. 1989, 104, 299–308.
- Vasconcelos, P.F.; Travassos da Rosa, A.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dégallier, N.; Travassos da Rosa, J.F. Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses. Cad. Saúde Pública 2001, 17, S155–S164.
- Pinheiro, F.P.; Freitas, R.B.; da Rosa, J.F.T.; Gabbay, Y.B.; Mello, W.A.; LeDuc, J.W. An Outbreak of Mayaro Virus Disease in Belterra, Brazil I. Clinical and Virological Findings. Am. J. Trop. Med. Hyg. 1981, 30, 674–681.
- Izurieta, R.O.; DeLacure, D.A.; Izurieta, A.; Hoare, I.A.; Reina Ortiz, M. Mayaro virus: The jungle flu. Virus Adapt. Treat. 2018, 10, 9–17.
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347.
- Aguilar-Luis, M.A.; del Valle-Mendoza, J.; Silva-Caso, W.; Gil-Ramirez, T.; Levy-Blitchtein, S.; Bazán-Mayra, J.; Zavaleta-Gavidia, V.; Cornejo-Pacherres, D.; Palomares-Reyes, C.; del Valle, L.J. An emerging public health threat: Mayaro virus increases its distribution in Peru. Int. J. Infect. Dis. 2020, 92, 253–258.
- Mourão, M.P.G.; de Souza Bastos, M.; de Figueiredo, R.P.; Gimaque, J.B.L.; dos Santos Galusso, E.; Kramer, V.M.; de Oliveira, C.M.C.; Naveca, F.G.; Figueiredo, L.T.M. Mayaro Fever in the City of Manaus, Brazil, 2007–2008. Vector-Borne Zoonotic Dis. 2011, 12, 42–46.
- Tesh, R.B.; Watts, D.M.; Russell, K.L.; Damodaran, C.; Calampa, C.; Cabezas, C.; Ramirez, G.; Vasquez, B.; Hayes, C.G.; Rossi, C.A.; et al. Mayaro Virus Disease: An Emerging Mosquito-Borne Zoonosis in Tropical South America. Clin. Infect. Dis. 1999, 28, 67–73.
- Coimbra, T.L.M.; Santos, C.L.S.; Suzuki, A.; Petrella, S.M.C.; Bisordi, I.; Nagamori, A.H.; Marti, A.T.; Santos, R.N.; Fialho, D.M.; Lavigne, S.; et al. Mayaro virus: Imported cases of human infection in São Paulo State, Brazil. Rev. Inst. Med. Trop. São Paulo 2007, 49, 221–224.
- Talarmin, A.; Chandler, L.J.; Kazanji, M.; de Thoisy, B.; Debon, P.; Lelarge, J.; Labeau, B.; Bourreau, E.; Vié, J.C.; Shope, R.E.; et al. Mayaro virus fever in French Guiana: Isolation, identification, and seroprevalence. Am. J. Trop. Med. Hyg. 1998, 59, 452–456.
- Mackay, I.M.; Arden, K.E. Mayaro virus: A forest virus primed for a trip to the city? Microbes Infect. 2016, 18, 724–734.
- Arenívar, C.; Rodríguez, Y.; Rodríguez-Morales, A.J.; Anaya, J.-M. Osteoarticular manifestations of Mayaro virus infection. Curr. Opin. Rheumatol. 2019, 31, 512–516.
- Azevedo, R.S.S.; Silva, E.V.P.; Carvalho, V.L.; Rodrigues, S.G.; Neto, J.P.N.; Monteiro, H.A.O.; Peixoto, V.S.; Chiang, J.O.; Nunes, M.R.T.; Vasconcelos, P.F.C. Mayaro Fever Virus, Brazilian Amazon. Emerg. Infect. Dis. 2009, 15, 1830–1832.
- Neumayr, A.; Gabriel, M.; Fritz, J.; Günther, S.; Hatz, C.; Schmidt-Chanasit, J.; Blum, J. Mayaro Virus Infection in Traveler Returning from Amazon Basin, Northern Peru. Emerg. Infect. Dis. 2012, 18, 695–696.
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rodríguez, Y.; Pacheco, Y.; Anaya, J.-M.; Ramírez-Santana, C. Mayaro: An emerging viral threat? Emerg. Microbes Infect. 2018, 7, 1–11.
- Dupuis-Maguiraga, L.; Noret, M.; Brun, S.; Grand, R.L.; Gras, G.; Roques, P. Chikungunya Disease: Infection-Associated Markers from the Acute to the Chronic Phase of Arbovirus-Induced Arthralgia. PLoS Negl. Trop. Dis. 2012, 6, e1446.
- Chow, A.; Her, Z.; Ong, E.K.S.; Chen, J.; Dimatatac, F.; Kwek, D.J.C.; Barkham, T.; Yang, H.; Rénia, L.; Leo, Y.-S.; et al. Persistent Arthralgia Induced by Chikungunya Virus Infection is Associated with Interleukin-6 and Granulocyte Macrophage Colony-Stimulating Factor. J. Infect. Dis. 2011, 203, 149–157.
- Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; Brochard, P.; Guigand, L.; Dubreil, L.; Lebon, P.; Verrier, B.; et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Invest. 2010, 120, 894–906.
- Noret, M.; Herrero, L.; Rulli, N.; Rolph, M.; Smith, P.N.; Li, R.W.; Roques, P.; Gras, G.; Mahalingam, S. Interleukin 6, RANKL, and Osteoprotegerin Expression by Chikungunya Virus-Infected Human Osteoblasts. J. Infect. Dis. 2012, 206, 455–457.
- Morrison, T.E.; Oko, L.; Montgomery, S.A.; Whitmore, A.C.; Lotstein, A.R.; Gunn, B.M.; Elmore, S.A.; Heise, M.T. A Mouse Model of Chikungunya Virus–Induced Musculoskeletal Inflammatory Disease: Evidence of Arthritis, Tenosynovitis, Myositis, and Persistence. Am. J. Pathol. 2011, 178, 32–40.
- Gardner, J.; Anraku, I.; Le, T.T.; Larcher, T.; Major, L.; Roques, P.; Schroder, W.A.; Higgs, S.; Suhrbier, A. Chikungunya Virus Arthritis in Adult Wild-Type Mice. J. Virol. 2010, 84, 8021–8032.
- Tappe, D.; Pérez-Girón, J.V.; Just-Nübling, G.; Schuster, G.; Gómez-Medina, S.; Günther, S.; Muñoz-Fontela, C.; Schmidt-Chanasit, J. Sustained Elevated Cytokine Levels during Recovery Phase of Mayaro Virus Infection. Emerg. Infect. Dis. 2016, 22, 750–752.
- Santiago, F.W.; Halsey, E.S.; Siles, C.; Vilcarromero, S.; Guevara, C.; Silvas, J.A.; Ramal, C.; Ampuero, J.S.; Aguilar, P.V. Long-Term Arthralgia after Mayaro Virus Infection Correlates with Sustained Pro-inflammatory Cytokine Response. PLoS Negl. Trop. Dis. 2015, 9, e0004104.
- Santos, F.M.; Dias, R.S.; de Oliveira, M.D.; Costa, I.C.T.A.; de Souza Fernandes, L.; Pessoa, C.R.; da Matta, S.L.P.; Costa, V.V.; Souza, D.G.; da Silva, C.C.; et al. Animal model of arthritis and myositis induced by the Mayaro virus. PLoS Negl. Trop. Dis. 2019, 13, e0007375.
- Churchman, S.M.; Ponchel, F. Interleukin-7 in rheumatoid arthritis. Rheumatology 2008, 47, 753–759.
- Yoo, S.-A.; Yoon, H.-J.; Kim, H.-S.; Chae, C.-B.; Falco, S.D.; Cho, C.-S.; Kim, W.-U. Role of placenta growth factor and its receptor flt-1 in rheumatoid inflammation: A link between angiogenesis and inflammation. Arthritis Rheum. 2009, 60, 345–354.
- Camini, F.C.; da Silva Caetano, C.C.; Almeida, L.T.; da Costa Guerra, J.F.; de Mello Silva, B.; de Queiroz Silva, S.; de Magalhães, J.C.; de Brito Magalhães, C.L. Oxidative stress in Mayaro virus infection. Virus Res. 2017, 236, 1–8.
- da Silva Caetano, C.C.; Camini, F.C.; Almeida, L.T.; Ferraz, A.C.; da Silva, T.F.; Lima, R.L.S.; de Freitas Carvalho, M.M.; de Freitas Castro, T.; Carneiro, C.M.; de Mello Silva, B.; et al. Mayaro Virus Induction of Oxidative Stress is Associated With Liver Pathology in a Non-Lethal Mouse Model. Sci. Rep. 2019, 9, 15289.
- Dhanwani, R.; Khan, M.; Alam, S.I.; Rao, P.V.L.; Parida, M. Differential proteome analysis of Chikungunya virus-infected new-born mice tissues reveal implication of stress, inflammatory and apoptotic pathways in disease pathogenesis. Proteomics 2011, 11, 1936–1951.
- Hosakote, Y.M.; Jantzi, P.D.; Esham, D.L.; Spratt, H.; Kurosky, A.; Casola, A.; Garofalo, R.P. Viral-mediated Inhibition of Antioxidant Enzymes Contributes to the Pathogenesis of Severe Respiratory Syncytial Virus Bronchiolitis. Am. J. Respir. Crit. Care Med. 2011, 183, 1550–1560.
- Kayesh, M.E.H.; Ezzikouri, S.; Sanada, T.; Chi, H.; Hayashi, Y.; Rebbani, K.; Kitab, B.; Matsuu, A.; Miyoshi, N.; Hishima, T.; et al. Oxidative Stress and Immune Responses During Hepatitis C Virus Infection in Tupaia belangeri. Sci. Rep. 2017, 7, 9848.
- Gil, L.; Martínez, G.; Tápanes, R.; Castro, O.; González, D.; Bernardo, L.; Vázquez, S.; Kourí, G.; Guzmán, M.G. Oxidative Stress in Adult Dengue Patients. Am. J. Trop. Med. Hyg. 2004, 71, 652–657.
- Cavalheiro, M.G.; Costa, L.S.D.; Campos, H.S.; Alves, L.S.; Assunção-Miranda, I.; Poian, A.T.D.; Cavalheiro, M.G.; Costa, L.S.D.; Campos, H.S.; Alves, L.S.; et al. Macrophages as target cells for Mayaro virus infection: Involvement of reactive oxygen species in the inflammatory response during virus replication. An. Acad. Bras. Ciênc. 2016, 88, 1485–1499.
- de Castro-Jorge, L.A.; de Carvalho, R.V.H.; Klein, T.M.; Hiroki, C.H.; Lopes, A.H.; Guimarães, R.M.; Fumagalli, M.J.; Floriano, V.G.; Agostinho, M.R.; Slhessarenko, R.D.; et al. The NLRP3 inflammasome is involved with the pathogenesis of Mayaro virus. PLOS Pathog. 2019, 15, e1007934.
- Yang, Z.; Min, Z.; Yu, B. Reactive oxygen species and immune regulation. Int. Rev. Immunol. 2020, 1–7.
- Chen, W.; Foo, S.-S.; Rulli, N.E.; Taylor, A.; Sheng, K.-C.; Herrero, L.J.; Herring, B.L.; Lidbury, B.A.; Li, R.W.; Walsh, N.C.; et al. Arthritogenic alphaviral infection perturbs osteoblast function and triggers pathologic bone loss. Proc. Natl. Acad. Sci. USA 2014, 111, 6040–6045.
- Rulli, N.E.; Rolph, M.S.; Srikiatkhachorn, A.; Anantapreecha, S.; Guglielmotti, A.; Mahalingam, S. Protection from Arthritis and Myositis in a Mouse Model of Acute Chikungunya Virus Disease by Bindarit, an Inhibitor of Monocyte Chemotactic Protein-1 Synthesis. J. Infect. Dis. 2011, 204, 1026–1030.
- Bengue, M.; Ferraris, P.; Baronti, C.; Diagne, C.T.; Talignani, L.; Wichit, S.; Liegeois, F.; Bisbal, C.; Nougairède, A.; Missé, D. Mayaro Virus Infects Human Chondrocytes and Induces the Expression of Arthritis-Related Genes Associated with Joint Degradation. Viruses 2019, 11, 797.
- Torres, J.R.; Russell, K.L.; Vasquez, C.; Barrera, R.; Tesh, R.B.; Salas, R.; Watts, D.M. Family Cluster of Mayaro Fever, Venezuela. Emerg. Infect. Dis. 2004, 10, 1304–1306.
- Figueiredo, L.T.M.; Nogueira, R.M.R.; Cavalcanti, S.M.B.; Schatzmayr, H.; da Rosa, A.T.; Figueiredo, L.T.M.; Nogueira, R.M.R.; Cavalcanti, S.M.B.; Schatzmayr, H.; da Rosa, A.T. Study of two different enzyme immunoassays for the detection of Mayaro virus antibodies. Mem. Inst. Oswaldo Cruz 1989, 84, 303–307.
- Earnest, J.T.; Basore, K.; Roy, V.; Bailey, A.L.; Wang, D.; Alter, G.; Fremont, D.H.; Diamond, M.S. Neutralizing antibodies against Mayaro virus require Fc effector functions for protective activity. J. Exp. Med. 2019, 216, 2282–2301.


