Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Andrea Giannini | + 3661 word(s) | 3661 | 2021-10-18 08:37:50 | | | |
| 2 | Vivi Li | Meta information modification | 3661 | 2021-10-19 05:45:56 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Giannini, A. Neuroendocrine Changes during Menopausal Transition. Encyclopedia. Available online: https://encyclopedia.pub/entry/15106 (accessed on 25 June 2026).
Giannini A. Neuroendocrine Changes during Menopausal Transition. Encyclopedia. Available at: https://encyclopedia.pub/entry/15106. Accessed June 25, 2026.
Giannini, Andrea. "Neuroendocrine Changes during Menopausal Transition" Encyclopedia, https://encyclopedia.pub/entry/15106 (accessed June 25, 2026).
Giannini, A. (2021, October 18). Neuroendocrine Changes during Menopausal Transition. In Encyclopedia. https://encyclopedia.pub/entry/15106
Giannini, Andrea. "Neuroendocrine Changes during Menopausal Transition." Encyclopedia. Web. 18 October, 2021.
Copy Citation
Menopause is the permanent cessation of menstrual cycles following the loss of ovarian follicular activity. Quality of life of postmenopausal woman is the result of a series of psychobiological transformations, that see in the reduction of sex hormones and steroids the etiopathogenetic determinant moment. Symptoms of menopause range from somatic side such as metabolic changes, increased cardiovascular disease, irregular vaginal bleeding, urogenital symptoms, vaginal dryness, osteoporosis and risk of bones fractures to changes of central nervous system as vasomotor symptoms, sleep disruption, mood changes, migraine, sexual dysfunctions.
neuroendocrine changes
menopause
symptoms
central nervous system
quality of life
1. Introduction
Quality of life of postmenopausal woman is the result of a series of psychobiological transformations, that see in the reduction of sex hormones and steroids the etiopathogenetic determinant moment. During climacteric period 80% of women complain about subjective bothersome symptomatology that often negatively impact the quality of life (QoL) in personal, social and working activities. Symptoms of menopause range from somatic side (such as metabolic changes, increased cardiovascular disease, irregular vaginal bleeding, urogenital symptoms, vaginal dryness, osteoporosis and risk of bones fractures) to central nervous system (CNS) changes (vasomotor symptoms, sleep disruption, mood changes, migraine, sexual dysfunctions). Changes in neuroendocrine and neurotransmitter systems, related to decreased estrogen seem to negatively affect psycho-physical wellbeing of women and could explain, at least in part, the increased incidence of mental disorders and neurological in the postmenopausal era, respect to women in fertility age.
This group of manifestations includes alterations of psychological balance represented above all from irritability, emotional instability with crisis of sudden crying, insomnia, fatigue, apathy. In particularly predisposed patients, such imbalances can hesitate in real psychiatric illnesses, or anxiety disorders and depressive syndrome [1][2]. Other alterations due to postmenopausal estrogen deficiency lead to alterations in hypothalamic thermoregulatory homeostasis, contributing to the onset of heat flashes and sweats. Several studies instead show a correlation between menopause and the onset of conditions such as Alzheimer’s disease or Parkinson’s disease, thus suggesting a plausible link between neurodegenerative disease and climacteric period.
However, independently from these two pathologies there is a decline of cognitive faculties of the woman (memory changes or cognitive impairment), related to hypoestrogenism. Two other elements to take into consideration when talking about alterations of the central nervous system are migraines, which increase in menopausal women, and to disorders of the sexual sphere (sexual arousal and satisfaction). It is fundamental to know the mechanisms underlying the CNS modifications during menopause, related to hypoestrogenism, so as to be able to create appropriate target therapy for the patients, improving their quality of life. In fact, the central nervous system is now one of the major targets of sex steroids that cannot be disregarded when dealing with the problem of choosing a particular type of menopausal hormonal therapy (MHT). Due to the constant increase of the ageing population and of the number of women experiencing climacteric periods, it is mandatory to understand the relevance of menopausal transition and the postmenopausal period that has relevance for every women worldwide. In fact, nowadays women spent more than one third of their lifespan in the transition and in the postmenopausal period, with potential negative impact in their general quality of life, family, relationships and working activities. The care of women during the climacteric transition involves several healthcare providers who have to monitor and promote healthy activities, prevention strategies, and to prescribe medical or alternative compounds according to the women’s need and personal risk factors. The concept that women are differently vulnerable to menopausal changes strength the concepts of developing a personalized approach and a gender-specific medicine.
2. Neuroendocrine Changes in Menopause
Menopause signs start with the reduction of follicle reserves and the subsequent impairment of ovarian function [3]. From a clinical standpoint, a suitable and reproducible classification of the nomenclature and stages for reproductive ageing was developed during the Stages of Reproductive Aging Workshop (STRAW) in 2001 and implemented in 2012 (STRAW+10). This system aims to classify womens‘ adult life in three different phases: reproductive life, menopausal transition and post-menopausal period. These three phases are than divided into seven stages with final menstrual period (FMP) as the most central event. The STRAW+10 criteria classify bleeding patterns for the early and the late menopausal transition and suggested changes to the criteria for the late reproductive stage (Stage −3) and the early postmenopausal period (Stage +1), provided specified on the length of the late transition (Stage −1) and early postmenopausal period (Stage +1). Moreover, the system suggested applying these rules regardless of the age, body constitution and weight, lifestyle and ethnicity. This progression is characterized by several hormonal modifications that are described as the menopausal transition that take some years to conclude. These changes begin with a significant reduction of the concentrations of inhibin B, thus determining a lack of negative feedback on pituitary FSH secretion [3][4].
The decreased secretion of inhibin B causes an increase in FSH production throughout the menstrual cycle, especially during the follicular phase. Ovulation is anticipated and progesterone secretion slowly decreases. In the premenopausal period the FSH increases, but reduced levels of estrogens, progesterone and LH still exist, but the production of estradiol slowly becomes insufficient and only rarely determines the peak of LH, causing anovulatory cycles. In this case, besides FSH, LH also tends to increase. In menopausal transition, the increase in FSH is 10 times bigger than that of LH; this increase is secondary to the lack of inhibin and to the estrogen feedback, as already mentioned. The levels of estradiol are slowly reduced in favor of an increase in estrone (produced by the aromatization of ovarian and adrenal androstenedione). Estrone has 1/3 the activity compared to estradiol. In addition to inhibin and progesterone, AMH also decreases. In contrast, GnRH increases the frequency and magnitude of the peak secretion (increase in gonadotropins). FSH and LH levels reach their highest levels about 1–3 years after menopause. In women undergoing ovariectomy, the levels of the gonadotropins increase more rapidly. Hypoestrogenism in menopause is associated with a lower sensitivity of the hypothalamus-hypophysis-gonadal axis and its structural and functional modifications. This explains the menopausal symptoms that appear in the transitional period such as hot flashes, insomnia and night sweats. In fact, estrogen and progesterone regulate, in addition to that, the proliferation and maturation of oligodendrocytes, glial cells, increasing number of synapses, promote angiogenic activity of endothelial cells and modulate mitochondrial activity. It is possible to identify two different type of estrogen receptors: ER alpha and beta. We find ER-alpha in the prefrontal cortex, hippocampus, amygdala and hypothalamus (memory and emotional process regions). ER-beta is expressed in the hippocampus, neocortex claustrum, thalamus and in mitochondria (where it exerts a protective action against oxidative diseases like Alzheimer’s disease). The interaction of estradiol with these receptors allow the release and metabolism regulation of neurotransmitters such as dopamine, serotonin, acetylcholine, β-endorphin and some neurosteroids such as allopregnanolone and dehydroepiandrosterone (DHEA) [5].
From a neurobiological standpoint, the monoamine neurotransmitters serotonin, norepinephrine and dopamine seem to modulate both hot flashes and mood disorders. Thus, an impairment of these regulatory pathways can favor the onset of depression when that dysregulation occurs within prefrontal cortex and limbic system that are SNC areas involved in mood control. Moreover, a deregulation of the hypothalamic thermoregulatory centers impacts the onset of vasomotor symptoms. In this view, the treatment of hot flashes with the administration of menopausal hormonal therapy (MHT) can prevent or reduce depressive symptoms in vulnerable women, instead of treating only mood disorders without controlling the vasomotor symptoms. However, in addition to the cessation of estrogenic secretion by the ovaries, a production of sex hormones at the intracranial level remains. Nowadays, gonadal steroid hormones are well recognized to be actively involved in the processes of CNS growth, differentiation, physiology, and ageing. Dehydroepiandrosterone sulphate (DHEAS) levels rise during reproductive age, reaching maximum peripheral concentrations around 30 years of age while around the seventh decade the levels of DHEAS progressively decline to 10–20% of the previous highest concentrations [6]. DHEAS is the principal peripheral product of adrenal glands which is the main source of circulating steroids. Moreover, ovarian thecal cells produce around 20% of peripheral DHEA under the regulation of LH and CNS [7][8]. Consistent evidence from the literature indicates that DHEA acts on CNS through antagonizing GABA-A receptor in a dose-dependent manner thus determining an increase in neuronal excitability and resulting in improvement of memory performances [9][10]. Furthermore, the conditions of low sexual desire and responsiveness in younger women and low responsiveness in older women seem to be significantly associated to low peripheral levels of DHEA relative to age [11]. In extragonadal tissues such as adipose tissue, brain, and bone, DHEA and DHEAS are converted either to androstenedione to testosterone that may be aromatized to estrone (E1) or estradiol (E2) or dihydrotestosterone (DHT). For that reason, DHEAS is considered a therapeutic option for the treatment of the androgen deficiency syndrome in menopause. During menopause, a reduction of another neurotransmitter—allopregnanolone (ALLO)—is also observed ALLO is a 3,5-reduced metabolite of progesterone produced by the complex 5-α-reductase and 3-α-hydroxysteroid oxidoreductase in the CNS, adrenals and ovaries [12][13]. It is the main neurosteroid derived from cholesterol in the neocortex, subcortical and hippocampus [13][14][15]. ALLO has anxiolytic and relaxant properties and it affects mood and behavior. Reduced serum and cerebrospinal fluid concentration of ALLO have been reported in women with depression [16][17][18]. New data confirm a putative effect of ALLO in γ-aminobutyric acid receptor A (GABA-A) receptors. This effect seems to be modulated by ovarian steroids, and estrogens and progesterone control the GABA response through a long-term genomic action [19][20]. In addition, hypoestrogenism causes a reduction secretion of β-endorphin (in β-EP), which is the main biologically relevant and active endogenous opioid peptide with neuroendocrinologic, analgesic and thermoregulatory properties. The postmenopausal reduction was shown to be associated with vasomotor symptoms and mood disorders [4][21]. DHEA supplementation results in the increase of plasmatic, hippocampal and hypothalamic β-EP concentrations and this effect seem to be dose-related thus improving menopause symptoms. MHT is able to positively influence on climacteric symptomatology of the CNS, by restoring an adequate adrenergic tone, serotoninergic and opioid, rebalancing the whole complex of induced neuroendocrine modifications from menopause [2].
2.1. Vasomotor Symptoms and Sleep Disruption
Vasomotor symptoms, in particular hot flushing, are the most frequent symptoms in menopause. Vasomotor symptoms occur in 75–80% of women during the late premenopause and early postmenopause period. In 5% of persons who experience these changes, hot flushes can remain decades after menopause [22][23][24]. A hot flush is an unexpected event of vasodilatation in the face and neck, with abundant sweating with subsequent shivering and feeling of coldness or redness. The duration can range from a few seconds to even 60 min and can be moderate or severe according to the number and the strength of the events. Black race, smoking and overweight are risk factors for vasomotor symptoms [25][26]. The number of daily hot flushes and their intensity can impair general QoL, modify lifestyle and determine stress and anxiety [27]. This bothersome symptom is caused by a deregulation of the hypothalamic thermoregulatory anterior nuclei and the preoptic region. The variation of the corporeal temperature from the baseline stimulates endocrine mediators to start regulatory actions to increase or reduce the production and the dispersion of energy as required to return to baseline temperature. A compression of the thermoregulatory area of the hypothalamus intermittently releases heat in response to a slight increase of the temperature [24]. Impairment of thermoregulation can be associated with deregulation of the serotoninergic and noradrenergic pathway that occur during the fluctuations of estrogens concentrations. Indeed, vasomotor symptoms decrease with passing years, demonstrating that they are triggered by hormone fluctuations and not only hypoestrogenism. The brain undergoes a readjustment to the different sex hormone concentrations during menopausal transition. Many studies have shown a correlation between hot flashes and alterations in glucose metabolism. During menopause the glucose levels in the brain are reduced and even at peripheral level there is an altered glucose tolerance. In fact, ketone bodies and the acid metabolism of fats increase. The SWAN study [28] showed a correlation between hot flushes and dysregulation of glucose metabolism, demonstrated by an increase in fasting blood glucose and HOMA scores. In addition, the onset of hot flashes are more frequent during the first 4 h of sleep, thus determining significant disruption of rest and reduction of the sleep quality. Insomnia and sleep disturbance are prevalent symptoms of perimenopause transition. They are maximal during late perimenopause and persist in post-menopause, independently of vasomotor symptoms [29]. The mechanism of sleep disturbance is not known, but the superchiasmatic nucleus may have a role given its estrogen receptor beta expression in relation to circadian rhythm. Furthermore, low levels of inhibin B are a marker of poor sleep quality. Moreover, many prospective studies indicate an association with poor sleep quality and dementia [30], with an accumulation of β-amyloid in the brain (Figure 1).
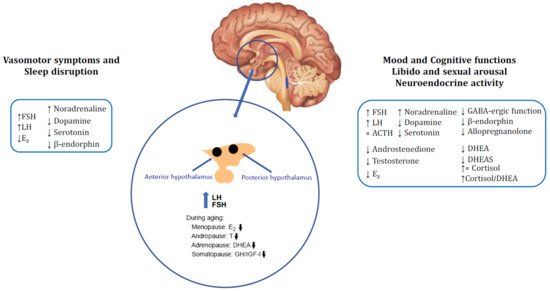
Figure 1. Brain neuroendocrine changes of menopause and the related symptoms: Modifications of gonadotropins, E2, neuromodulators and opioid tone in the hypothalamus causing onset of vasomotor symptoms and consequent sleep disruption. Mood, cognitive functions, libido, sexual arousal and neuroendocrine activity are close related to the impairment of GABAergic, opioid and neurosteroid milieu in central nervous system (CNS) occurred with ageing. Menopausal condition determines reduction in estrogens and ovarian and adrenal steroids (testosterone, androstenedione, DHEA and DHEA-S) which results in increase cortisol/DHEA ratio leading to sexual, mood and cognitive dysfunctions. ↑: increased, ↓: decreased, = unchanged.
2.2. Mood Disorders
Postmenopausal changes in neuropeptide levels and neurotransmitters are implicated in the pathogenesis of characteristic mood disorders of this woman’s life phase. Oscillations in the concentrations of steroids, more than their reduction, could cause depression during menopausal transition. Specially, fluctuations of estrogen impact an impaired regulation of CNS serotonin and noradrenaline paths. Beneficial effects of estrogen administration on mood in postmenopausal women depend on the direct action of gonadal steroids on neuronal activity and on the serotonergic system.
Estrogens modify the concentration of serotonin determining an increase of the rate of degradation of monoamine oxidase (MAO), enzyme responsible for the catabolism of serotonin. Effects exerted by estrogens on the mood cannot be excluded and they can also be partly linked to a direct effect on neuronal activity, given that such steroids would be able to modulate the cerebral blood flow, the glucose levels, as well as neuronal growth and synaptic activity. In the postmenopausal period, those women who start MHT across the perimenopausal transition or at the beginning of menopause seem to maintain glucose metabolism in some brain areas which present neurological functions dependent from the estrogen milieu, such as the hippocampus, entorhinal cortex, medial temporal cortex and posterior cingulate. On the contrary, glucose metabolism in women who do not undertake MHT during perimenopause and menopause, deteriorates in estrogen-dependent areas of the CNS, but seem to be improved in the pons, caudate and precuneus [31][32].
Studies that demonstrated this evidence enrolled symptomatic women and compared them to women who were premenopausal or asymptomatic, so the data derived are possibly only relevant to women who experience symptoms. Some symptoms like depression, irritability, anxiety as well as psychological processes may be affected by the fluctuation of neurosteroid synthesis and secretion. As previously mentioned, ALLO modulates stress, mood and the character traits, expressing anxiolytic properties and sedative-hypnotic, acting as an agonist on GABA-A receptors, [33][34]. MHT can modify the levels of neurosteroids increasing ALLO and reducing DHEA. In this view, the raise of ALLO with MHT could be responsible of the reduction of the anxiety and of sedative effects on women in menopause [35]. However, DHEA is synthesized as a neurosteroid, from the CNS and acts as an antagonist on the receptor GABA-A, with a consequent increase in excitability neuronal and anxiety in menopause women. DHEA levels undergo a decrease starting from the third decade of life (at the age of 70 the concentration plasma of that steroid is reduced to 20% of the maximum peak observed between 18 and 25 years) [36]. Therefore, it has been shown that administration of DHEA in these women leads to an improvement in quality of life of these women, in their cognitive faculties, like memory, anxiety and sexual dysfunction (Figure 1).
2.3. Neurological Disorders, Alzheimer’s Disease and Parkinson’s Disease
Female individuals have a higher constitutional risk of developing chronic and degenerative disorders as the reduction of the short-term memory and the increase in incidence of Alzheimer’s disease (AD), compared to men of the same age [37]. There is a close link between estrogens and cognitive and memory functions. Estrogens play a key role in the physiopathology of degenerative CNS diseases, as they act in areas like the prefrontal cortex, hippocampus and striatum that control learning, registering, and retrieving information, judgment, language skills. Sex hormones and their receptors contribute to the processes of synaptogenesis [38], neuronal growth and neuronal transmission. Also, they limit the inflammatory response in the CSN that cause dystrophy and dementia. Among degenerative pathologies we must recall Alzheimer’s disease. Although the etiology of the memory impairment related with AD is multifactorial, it has been described that estrogens have a protective effect and prolong the latency period of the disease [39]. Some epidemiological data support the hypothesis that MHT may reduce the incidence of AD in women [37][40]. In fact, there is dose-dependent association between estrogens supplementation and frequency of AD, as demonstrated by the reduction of the relative risk of AD with the increase of the estrogen dose and duration of therapy (the risk of AD is reduced of about 50% in patients who have used estrogen, if therapy was undertaken at an early age) [37]. Reduction of cognitive performances, observed in patients suffering from AD, recognizes as neurochemical defect the alteration of the system cholinergic neurotransmitter. In fact, estrogens provide trophic support to cholinergic cells, stimulating the release of acetylcholine and choline acetyltransferase (ChAT), both cholinergic functioning markers; they are also involved in the regulation of survival, regeneration and neuronal plasticity and develop antioxidant neuroprotective action and appear inhibit the formation of β-amyloid [39]. In postmenopausal women affected by AD there were studied gonadotrophin levels, founding a relation among increased β-amyloid and high level of FSH and LH [41]. Many other studies show that the expression of apolipoprotein E, which is responsible of transport of lipids in the blood, is stimulated in hippocampus and prefrontal cortex, through the interaction of 17β-estradiol with estrogen receptors. In women the correlation between apolipoprotein E and AD is more evident than in men, demonstrating the main role of female sex hormones in protecting from neurodegenerative diseases. Lastly, the Mayo Clinic Cohort Study of Oophorectomy and Aging-2, a cohort of patients who experienced bilateral salpingo-oophorectomy before age 50 and reaching natural menopause, shows an accelerated accumulation of multimorbidity and a dysregulation of numerous cellular, tissue, organ and system essential ageing process, for example DNA methylation levels [42]. Surgical menopause may lead to medial temporal lobe structural abnormalities, smaller amygdala volumes, thinner para-hippocampal-entorhinal cortices and β-amyloid deposition thus, increasing the risk of cognitive dysfunction or dementia and presence of AD, compared to women control group [43]. In conclusion, MHT significantly improves cognitive capabilities, positively modulating patterns of activity brain during psychometric tests, facilitating the blood flow towards structures that modulate the memory and improving mood. Another possible correlation is between hypoestrogenism and Parkinson’s disease. There are data in the literature which attest a role of estrogens as potentials neuroprotective agents of dopaminergic neurons. Sawada and coll. have shown that estradiol is protective on mesencephalic dopaminergic neurons from oxidative stress and apoptosis induced by bleomycin [44]. Moreover, estrogens inhibit the uptake of neurotoxins that induce degeneration of dopaminergic neurons [45]. Clinical studies show that estrogen replacement therapy produces less severe symptoms in women with PD in the early stage of the disease, especially if hormone therapy is undertaken before starting levodopa therapy [46]. However, the data are conflicting and there are different opinions in the literature regarding the efficacy of MHT in patients suffering from Parkinson’s disease. Therefore further experimental and clinical studies are necessary to define which role sex steroids may have in PD.
2.4. Migraine
Women who had migraine during their fertility age and, specially, in the premenstrual period, are more susceptible of headaches during the menopause transition. However, the prevalence of migraine in menopause ranges from 10% to 29% and decreases in post-menopause [47]. The reason is that migraine is triggered by sudden declines of estrogen levels and their action in hippocampus and prefrontal cortex. Lower estrogen levels cause vasodilatation, release and plasma extravasation of proinflammatory mediators and consequently cause migraine [48]. Over time the brain adapts its systems to lower hormone and steroid levels. In any case, not all women benefit from hormone therapy, and further scientific evidence is needed to explain the phenomenon.
2.5. Sexual Dysfunctions
Sexual dysfunction is frequent in post-menopause and it depends on hypoestrogenism as well as cultural factors. The prevalence is about 8–50% and it is reported to rise with age from the third decade as well as after oophorectomy. Sexual dysfunction in menopause involves vaginal dryness, reduced libido and reduced pleasure during sexual intercourse. Postmenopausal hormonal deficiency negatively influences sexual function since estrogenic support fails. Estrogens maintain the trophism and elasticity of the vaginal walls, vaginal lubrification and trophism of the entire genitourinary tract. The reduced vulvovaginal receptivity in turn determines dyspareunia and consequently reduced libido. However, the mood and cognitive changes of CNS, described above, contribute to determine disorders of the sexual sphere in a woman during menopause. Sexual behavior in women is significantly determined by endogenous sex steroid concentrations. Several reports suggest that testosterone therapy can be beneficial for patients with hypoactive sexual desire disorder [49]. In parallel, many studies have reported a negative correlation between low peripheral DHEA levels and the impairment of sexual function in pre- and postmenopausal women. In the CNS the neurosteroid DHEA is a regulator of neurotransmitter receptors. In fact, androgen deficiency syndrome, a consequence of a reduced androgen synthesis at both adrenal and ovarian levels, is recognized as a distinctive trait of reduced libido, a sense of psychological well-being and motivation, as well as a cause of easy fatigability [31]. In addition, to improve genital and urinary trophism MHT can play an important role in symptom relief, even if it does not seem to have primary effects on libido.
References
- Genazzani, A.R.; Gambacciani, M. Hormone replacement therapy: The prospectives for the 21st century. Maturitas 1999, 32, 11–17.
- Genazzani, A.R.; Spinetti, A.; Gallo, R.; Bernardi, F. Menopause and the central nervous system: Intervention options. Maturitas 1999, 3, 103–110.
- Davis, S.R. Menopause. Nat. Rev. Dis. Primers 2015, 1, 15004.
- Harlow, S.D.; Gass, M.; Hall, J.E.; Lobo, R.; Maki, P.; Rebar, R.W.; Sherman, S.; Sluss, P.M.; de Villiers, T.J. Executive summary of the stages of reproductive aging workshop. Addressing the unfinished agenda of staging reproductive aging. J. Clin. Endocrinol. Metab. 2012, 97, 1159–1168.
- Farrag, A.F.; Khedr, E.M.; Abdel-Aleem, H.; Rageh, T.A. Effects of Surgical Menopause on Cognitive Functions. Dement. Geriatr. Cogn. Disord. 2002, 13, 193–198.
- Orentreich, N.; Brind, J.L.; Vogelman, J.H.; Andres, R.; Baldwin, H. Long-term longitudinal measurements of plasma dehydroepiandrosterone sulfate in normal men. J. Clin. Endocrinol. Metab. 1992, 75, 1002–1004.
- Yamaji, T.; Ibayashi, H. Serum deydroepiandrosterone sulphate in normal and pathological canditions. J. Clin. Endocrinol. Metab. 1969, 29, 273–278.
- Corpechot, C.; Robert, P.; Axelson, M.; Sjovall, J.; Baulieu, E.E. Characterization and measurement of deydriepiandrosterone sulfate in the rat brain. Proc. Natl. Acad. Sci. USA 1981, 78, 4704–4707.
- Majewska, M.D.; Demirgoren, S.; Spivak, C.E.; London, E.D. The neurosteroid DHEA is an allosteric antagonist of the GABA A receptor. Brain Res. 1990, 526, 143–146.
- Baulieu, E.E. Dehydroepiandrosterone: A fountain of youth? J. Clin. Endocrinol. Metab. 1996, 81, 3147–3151.
- Davis, S.; Shah, S.M.; McKenzie, D.P.; Kulkarni, J.; Davison, S.; Bel, R.J. Dehydroepiandrosterone Sulfate Levels Are Associated with More Favorable Cognitive Function in Women. J. Clin. Endocrinol. Metab. 2008, 93, 3.
- Baulieu, E.E. Neurosteroids: A new function in the brain. Biol. Cell 1991, 71, 3–10.
- Robel, P.; Baulieu, E.E. Neurosteroids. Biosynthesis and function. Trends Endocrinol. Metab. 1994, 5, 1–8.
- Stoffel-Wagner, B. Neurosteroid metabolism in the human brain. Eur. J. Endocrinol. 2001, 145, 669–679.
- Stoffel-Wagner, B.; Watzka, M.; Steckelbroeck, S.; Ludwig, M.; Clusmann, H.; Bidlingmaier, F.; Casarosa, E.; Luisi, S.; Elger, C.E.; Beyenburg, S. Allopregnanolone serum levels and expression of 5-alpha-reductase and 3-alpha hydroxysteroid dehydrogenase isoforms in hippocampal and temporal cortex of patients with epilepsy. Epilepsy Res. 2003, 54, 11–19.
- Sundstrom, I.; Andersson, A.; Nyberg, S.; Purdy, R.H.; Bäckström, T. Patients with premenstrual syndrome have a different sensitivity to a neuroactive steroid during the menstrual cycle compared to control subjects. Neuroendocrinology 1998, 67, 126–138.
- Khisti, R.T.; Mandhane, S.N.; Chopde, C.T. The neurosteroid 3-alpha-hydroxy-5-alpha-pregnan-20-one induces catalepsy in mice. Neurosci. Lett. 1998, 251, 85–88.
- Johansson, I.M.; Birzniece, V.; Lindblad, C.; Olsson, T.; Bäckström, T. Allopregnanolone inhibits learning in the Morris water maze. Brain Res. 2002, 934, 125–131.
- Genazzani, A.R.; Bernardi, F.; Pluchino, N.; Begliuomini, S.; Lenzi, E.; Casarosa, E.; Luisi, M. Endocrinology of menopausal transition and its brain implications. CNS Spectr. 2005, 10, 449–457.
- Słopien, R.; Junik, R.; Meczekalski, B.; Halerz-Nowakowska, B.; Maciejewska, M.; Warenik-Szymankiewicz, A.; Sowińskib, J. Influence of hormonal replacement therapy on the regional cerebral blood flow in postmenopausal women. Maturitas 2003, 46, 255–262.
- Słopien, R.; Jasniewicz, J.; Meczekalski, B.; Warenik-Szymankiewicza, A.; Lianeri, M.; Jagodzińskib, P.P. Polymorphic variants of genes encoding MTHFR, MTR and MTHFD1 and the risk of depression in postmenopausal women in Poland. Maturitas 2008, 8, 252–255.
- Downs, J.L.; Wise, P.M. The role of the brain in female reproductive aging. Mol. Cell Endocrinol. 2009, 299, 32–38.
- Santoro, N. Symptoms of menopause: Hot flushes. Clin. Obstet. Gynecol. 2008, 51, 539–548.
- Freedman, R.R. Menopausal hot flashes: Mechanisms, endocrinology, treatment. J. Steroid Biochem. Mol. Biol. 2014, 142, 115–120.
- Richard-Davis, G.; Wellons, M. Racial and ethnic differences in the physiology and clinical symptoms of menopause. Semin. Reprod. Med. 2013, 31, 380–386.
- Duffy, O.K.; Iversen, L.; Aucott, L.; Hannaford, P.C. Factors associated with resilience or vulnerability to hot flushes and night sweats during the menopausal transition. Menopause 2013, 20, 383–392.
- Thurston, R.C.; Chang, Y.; Mancuso, P.; Matthews, K.A. Adipokines, adiposity, and vasomotor symptoms during the menopause transition: Findings from the Study of Women’s Health across the Nation. Fertil. Steril. 2013, 100, 793–800.
- Thurston, R.C.; Khoudary, S.R.E.; Sutton-Tyrrell, K.; Crandall, C.J.; Sternfeld, B.; Joffe, H.; Gold, E.B.; Selzer, F.; Matthews, K.A. Vasomotor symptoms and insulin resistance in the study of women’s health across the nation. J. Clin. Endocrinol. Metab. 2021, 97, 3487–3494.
- Kravitz, H.M.; Joffe, H. Sleep during the perimenopause: A Swan story. Obstet. Gynecol. Clin. N. Am. 2011, 38, 567–586.
- Yaffe, K.; Falvey, C.M.; Hoang, T. Connections between sleep and cognition in older adults. Lancet Neurol. 2014, 13, 1017–1028.
- Rasgon, N.L.; Silverman, D.; Siddarth, P.; Miller, K.; Ercoli, L.M.; Elman, S.; Lavretsky, H.; Huang, S.-C.; Phelpsc, M.E.; Small, G.W. Estrogen use and brain metabolic change in postmenopausal women. Neurobiol. Aging 2005, 26, 229–235.
- Maki, P.M.; Resnick, S.M. Longitudinal effects of estrogen replacement therapy on PET cerebral blood flow and cognition. Neurobiol. Aging 2000, 21, 373–383.
- Majewska, M.D. Neurosteroids: Endogenous bimodal modulators of the GABA A receptors. Mechanism of action and physiological significance. Prog. Neurobiol. 1992, 38, 379–395.
- Mellon, S.H. Neurosteroids: Action and clinical relevance. J. Clin. Endocrinol. Metab. 1994, 78, 1003–1008.
- Bernardi, F.; Pieri, M.; Stomati, M.; Luisi, S.; Palumbo, M.; Pluchino, N.; Ceccarelli, C.; Genazzani, A.R. Effect of different hormonal replacement therapies on circulating allopregnanolone and dehydroepiandrosterone levels in postmenopausal women. Gynecol. Endocrinol. 2003, 17, 65–77.
- Davis, S.R.; Burger, H.G. Androgens and the postmenopausal woman, Clinical Review. J. Clin. Endocrinol. Metab. 1996, 81, 2759–2763.
- Paganini-Hill, A.; Henderson, V. Estrogen deficiency and risk of Alzheimer’s Disease in women. Am. J. Epidemiol. 1994, 140, 256–261.
- Yao, J.; Irwin, R.W.; Zhao, L.; Nilsen, J.; Hamilton, R.T.; Brinton, R.D. Mitochondrial bioenergetic deficit precedes Alzheimer’s pathology in female mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2009, 106, 14670–14675.
- Sherwin, B.B. Estrogen effects on cognition in menopausal women. Neurology 1997, 48, S21–S26.
- Zandi, P.P.; Carlson, M.C.; Plassman, B.L.; Welshbohmer, K.A.; Mayer, L.S.; Steffens, D.C.; Breitner, J.C. Hormone replacement therapy and incidence of Alzheimer disease in older women: The Cache County Study. JAMA 2002, 288, 2123–2129.
- Brinton, R.D. Investigative models for determining hormone therapy-induced outcomes in brain: Evidence in support of a healthy cell bias of estrogen action. Ann. N. Y. Acad. Sci. 2005, 1052, 57–74.
- Rocca, W.A.; Rocca, L.G.; Smith, C.Y.; Grossardt, B.R.; Faubion, S.; Shuster, L.T.; Kirkland, J.L.; LeBrasseur, N.K.; Schafer, M.J.; Mielke, M.; et al. Loss of ovarian hormones and accelerated somatic and mental aging. Physiology 2018, 33, 374–383.
- Zeydan, B.; Tosakulwong, N.; Schwarz, C.G.; Senjem, M.L.; Gunter, J.L.; Reid, R.I.; Rocca, G.L.; Lesnick, T.G.; Smith, C.Y.; Bailey, K.R.; et al. Association of bilateral salpingo-oophorectomy before menopause onset with medial temporal lobe neurodegeneration. JAMA Neurol. 2019, 76, 1.
- Sawada, H.; Ibi, M.; Kihara, T.; Urushitani, M.; Akaika, A.; Shimohama, S. Estradiol protects mesencephalic dopaminergic neurons from oxidative stress-induced neuronal death. J. Neurosci. Res. 1998, 54, 707–719.
- Dluzen, D.E.; MCDermott, J.L. Gender differences in neurotoxicity of the nigrostriatal dopaminergic system: Implications for Parkinson’s disease. J. Gend. Specif. Med. 2000, 3, 36–42.
- Saunders-Pullman, R.; Gordon-Elliott, J.; Parides, M.; Fahn, S.; Saunders, H.R.; Bressman, S. The effect of estrogen replacement on early Parkinson’s disease. Neurology 1999, 52, 1417–1421.
- Short, R.A.; Bowen, R.L.; O’Brien, P.C.; Graff-Radford, N.R. Elevated gonadotropin levels in patients with Alzheimer disease. Mayo Clin. Proc. 2001, 76, 906–909.
- Hagemann, G.; Ugur, T.; Schleussner, E.; Mentzel, H.J.; Fitzek, C.; Witte, O.W.; Gaser, C. Changes in Brain Size during the Menstrual Cycle. PLoS ONE 2011, 6, e14655.
- Laumann, E.; Paik, A.; Rosen, R.C. Sexual dysfunction in the United States: Prevalence and predictors. JAMA 1999, 281, 537–544.
More
Information
Subjects:
Obstetrics & Gynaecology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
650
Revisions:
2 times
(View History)
Update Date:
19 Oct 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No