 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Sarah A. Purcell | + 3566 word(s) | 3566 | 2021-10-09 11:02:57 | | | |
| 2 | Jessie Wu | Meta information modification | 3566 | 2021-10-11 03:26:22 | | |
Video Upload Options
Many breast cancer survivors (BCS) gain fat mass and lose fat-free mass during treatment (chemotherapy, radiation, surgery) and estrogen suppression therapy, which increases the risk of developing comorbidities. Whether these body composition alterations are a result of changes in dietary intake, energy expenditure, or both is unclear. Thus, we reviewed studies that have measured components of energy balance in BCS who have completed treatment. Longitudinal studies suggest that BCS reduce self-reported energy intake and increase fruit and vegetable consumption. Although some evidence suggests that resting metabolic rate is higher in BCS than in age-matched controls, no study has measured total daily energy expenditure (TDEE) in this population. Whether physical activity levels are altered in BCS is unclear, but evidence suggests that light-intensity physical activity is lower in BCS compared to age-matched controls. We also discuss the mechanisms through which estrogen suppression may impact energy balance and develop a theoretical framework of dietary intake and TDEE interactions in BCS. Preclinical and human experimental studies indicate that estrogen suppression likely elicits increased energy intake and decreased TDEE, although this has not been systematically investigated in BCS specifically. Estrogen suppression may modulate energy balance via alterations in appetite, fat-free mass, resting metabolic rate, and physical activity. There are several potential areas for future mechanistic energetic research in BCS (e.g., characterizing predictors of intervention response, appetite, dynamic changes in energy balance, and differences in cancer sub-types) that would ultimately support the development of more targeted and personalized behavioral interventions.
1. Introduction
2. Estrogen Suppression in the Regulation of Energy Balance
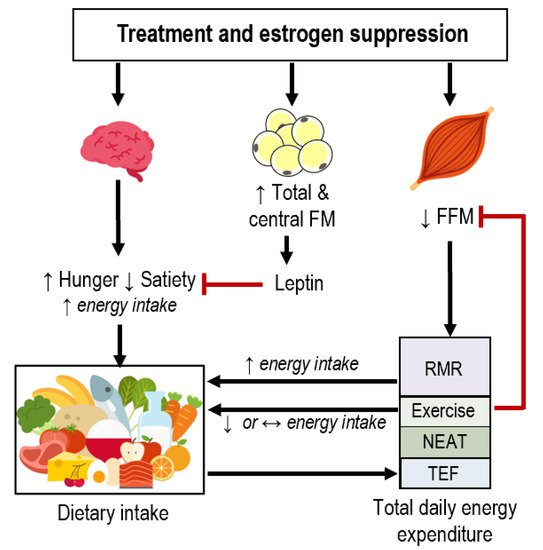
2.1. Estrogen and Appetite
2.2. Estrogen and Total Daily Energy Expenditure
2.3. Relationships between Dietary Intake and Energy Expenditure in Breast Cancer Survivors
3. Psychological Alterations and Energy Balance after Breast Cancer
4. Areas for Future Research and Conclusions
-
Expanded use of more accurate techniques such as doubly labeled water (2H2 and 18O), accelerometers and whole-room indirect calorimetry would help promote further understanding of TDEE and its components in different clinical populations. While these techniques are not practical in large sample sizes, they could provide useful insight on the mechanistic underpinnings of energy balance in BCS (and cancer survivors in general) in smaller samples. Other techniques that include repeated measures of body composition and energy expenditure [61][62] or mathematical models [63] may also help quantify energy balance in this population.
-
Use of stable isotopes to measure intake of food groups could be used to complement recall or record-based methods of dietary intake. For example, 13C/12C can be used describe intake of C4 plants (e.g., corn, cane sugars) and C3 plants (e.g., fruits and vegetables, wheat, nuts, seeds); similarly, 15N/14N can be used to characterize fish and meat intake [64][65]. Use of isotopes paired with repeated measures of dietary recall and TDEE would provide valuable insight of energy balance in BCS.
-
Inter-individual variability in body composition responses to exercise suggests that individuals compensate more or less to the same intervention. In other words, some individuals may increase EI, decrease physical activity, or both in response to exercise training. Elucidating the predictors of response and whether such predictors differ in BCS will help facilitate the design of more efficacious, personalized interventions for weight management.
-
Weight loss can be achieved through alterations in physical activity and dietary intake, but most individuals regain the weight they lost [66]. Physiological and psychological changes in appetite and energy expenditure in the context of an obesogenic environment underpin weight regain [67][68]. Characterization of energy balance during weight loss and maintenance in BCS—and whether this differs from individuals without previous cancer—would help generate more durable strategies for body weight management.
-
Eating behavior and appetite parameters are important determinants of dietary intake. As discussed in this review, there is modest evidence that appetite fluctuates across the menstrual cycle and menopausal transition due to altered sex hormones. Elucidation of the effects of sex hormones on appetite in estrogen-suppressed BCS may support the development of more targeted nutrition interventions.
-
There is increasing cross-sectional evidence that components of dietary intake and TDEE are related. Whether specific components of TDEE predict dietary intake and appetite in instances of energy imbalance is unclear in the general population and in people with chronic disease. Elucidating the complex interrelations among energy balance parameters in the context of different conditions may help better predict intervention response and devise better solutions for weight management.
-
Differentiation of outcomes according to tumor pathology (i.e., ER, PR, and human epidermal growth factor-2 status), patient age, and treatment modalities may also promote personalized intervention strategies. As previously reviewed [15][69], women who are premenopausal at diagnosis have a higher risk of FM gain compared to women who were postmenopausal at diagnosis. This is likely a direct result different treatment modalities and estrogen status; how these factors impact behavior and physiology related to energy balance is unknown.Finally, characterizing energy balance components in other cancer populations is warranted, especially in those that often undergo rigorous chemotherapy or hormonal treatments or are at risk for developing obesity (e.g., colorectal, prostate, ovarian cancers). This review focused on BCS because of the risk of weight gain, effect of hormonal therapies, and the availability of enough evidence to form conservative conclusions regarding dietary intake and energy expenditure. However, there is limited data on how various cancer types and treatment modalities may impact specific components of energy balance after treatment in other cancer types; it is also unclear if energy balance differs among cancer types or compared to individuals without previous cancer.
References
- American Cancer Society. Cancer Facts & Figures 2019; American Cancer Society: Atlanta, GA, USA, 2019.
- Ng, H.S.; Koczwara, B.; Roder, D.; Niyonsenga, T.; Vitry, A. Incidence of Comorbidities in Women with Breast Cancer Treated with Tamoxifen or an Aromatase Inhibitor: An Australian Population-Based Cohort Study. J. Comorbidity 2018, 8, 16–24.
- Bradshaw, P.T.; Stevens, J.; Khankari, N.; Teitelbaum, S.L.; Neugut, A.I.; Gammon, M.D. Cardiovascular Disease Mortality Among Breast Cancer Survivors. Epidemiology 2016, 27, 6–13.
- Caan, B.J.; Cespedes, F.E.M.; Prado, C.M.; Alexeeff, S.; Kroenke, C.H.; Bradshaw, P.; Quesenberry, C.P.; Weltzien, E.K.; Castillo, A.L.; Olobatuyi, T.A.; et al. Association of Muscle and Adiposity Measured by Computed Tomography with Survival in Patients with Nonmetastatic Breast Cancer. JAMA Oncol. 2018, 4, 798–804.
- Godinho-Mota, J.C.M.; Mota, J.F.; Goncalves, L.V.; Soares, L.R.; Schincaglia, R.M.; Prado, C.M.; Martins, K.A.; Freitas-Junior, R. Chemotherapy negatively impacts body composition, physical function and metabolic profile in patients with breast cancer. Clin. Nutr. 2021, 40, 3421–3428.
- Demark-Wahnefried, W.; Peterson, B.L.; Winer, E.P.; Marks, L.; Aziz, N.; Marcom, P.K.; Blackwell, K.; Rimer, B.K. Changes in weight, body composition, and factors influencing energy balance among premenopausal breast cancer patients receiving adjuvant chemotherapy. J. Clin. Oncol. 2001, 19, 2381–2389.
- Makari-Judson, G.; Braun, B.; Jerry, D.J.; Mertens, W.C. Weight gain following breast cancer diagnosis: Implication and proposed mechanisms. World J. Clin. Oncol. 2014, 5, 272–282.
- Ewertz, M.; Jensen, M.-B.; Gunnarsdóttir, K.Á.; Højris, I.; Jakobsen, E.H.; Nielsen, D.; Stenbygaard, L.E.; Tange, U.B.; Cold, S. Effect of Obesity on Prognosis after Early-Stage Breast Cancer. J. Clin. Oncol. 2010, 29, 25–31.
- Protani, M.; Coory, M.; Martin, J.H. Effect of obesity on survival of women with breast cancer: Systematic review and meta-analysis. Breast Cancer Res. Treat. 2010, 123, 627–635.
- Goodwin, P.J.; Ennis, M.; Pritchard, K.I.; Trudeau, M.E.; Koo, J.; Taylor, S.K.; Hood, N. Insulin- and obesity-related variables in early-stage breast cancer: Correlations and time course of prognostic associations. J. Clin. Oncol. 2012, 30, 164–171.
- Prado, C.M.; Purcell, S.A.; Alish, C.; Pereira, S.L.; Deutz, N.E.; Heyland, D.K.; Goodpaster, B.H.; Tappenden, K.A.; Heymsfield, S.B. Implications of low muscle mass across the continuum of care: A narrative review. Ann. Med. 2018, 50, 675–693.
- Rier, H.N.; Jager, A.; Sleijfer, S.; van Rosmalen, J.; Kock, M.C.J.M.; Levin, M.-D. Low muscle attenuation is a prognostic factor for survival in metastatic breast cancer patients treated with first line palliative chemotherapy. Breast 2017, 31, 9–15.
- Del Fabbro, E.; Parsons, H.; Warneke, C.L.; Pulivarthi, K.; Litton, J.K.; Dev, R.; Palla, S.L.; Brewster, A.; Bruera, E. The relationship between body composition and response to neoadjuvant chemotherapy in women with operable breast cancer. Oncologist 2012, 17, 1240–1245.
- Francis, P.A.; Pagani, O.; Fleming, G.F.; Walley, B.A.; Colleoni, M.; Láng, I.; Gómez, H.L.; Tondini, C.; Ciruelos, E.; Burstein, H.J.; et al. Tailoring Adjuvant Endocrine Therapy for Premenopausal Breast Cancer. N. Engl. J. Med. 2018, 379, 122–137.
- Sheean, P.M.; Hoskins, K.; Stolley, M. Body composition changes in females treated for breast cancer: A review of the evidence. Breast Cancer Res. Treat 2012, 135, 663–680.
- Gross, A.L.; May, B.J.; Axilbund, J.E.; Armstrong, D.K.; Roden, R.B.S.; Visvanathan, K. Weight change in breast cancer survivors compared to cancer-free women: A prospective study in women at familial risk of breast cancer. Cancer Epidemiol. Prev. Biomark. 2015, 24, 1262–1269.
- Visram, H.; Kanji, F.; Dent, S.F. Endocrine therapy for male breast cancer: Rates of toxicity and adherence. Curr. Oncol. 2010, 17, 17–21.
- Finkelstein, J.S.; Lee, H.; Burnett-Bowie, S.-A.M.; Pallais, J.C.; Yu, E.W.; Borges, L.F.; Jones, B.F.; Barry, C.V.; Wulczyn, K.E.; Thomas, B.J.; et al. Gonadal steroids and body composition, strength, and sexual function in men. N. Engl. J. Med. 2013, 369, 1011–1022.
- Hirschberg, A.L. Sex hormones, appetite and eating behaviour in women. Maturitas 2012, 71, 248–256.
- Brown, L.M.; Clegg, D.J. Central effects of estradiol in the regulation of food intake, body weight, and adiposity. J. Steroid Biochem. Mol. Biol. 2010, 122, 65–73.
- Butera, P.C. Estradiol and the control of food intake. Physiol. Behav. 2010, 99, 175–180.
- Sotonyi, P.; Gao, Q.; Bechmann, I.; Horvath, T.L. Estrogen promotes parvalbumin expression in arcuate nucleus POMC neurons. Reprod. Sci. 2010, 17, 1077–1080.
- Olofsson, L.E.; Pierce, A.A.; Xu, A.W. Functional requirement of AgRP and NPY neurons in ovarian cycle-dependent regulation of food intake. Proc. Natl. Acad. Sci. USA 2009, 106, 15932–15937.
- Belsito, K.; Vester, B.; Keel, T.; Graves, T.; Swanson, K. Impact of ovariohysterectomy and food intake on body composition, physical activity, and adipose gene expression in cats. J. Anim. Sci. 2009, 87, 594–602.
- Gorzek, J.F.; Hendrickson, K.C.; Forstner, J.P.; Rixen, J.L.; Moran, A.L.; Lowe, D.A. Estradiol and tamoxifen reverse ovariectomy-induced physical inactivity in mice. Med. Sci. Sports Exerc. 2007, 39, 248–256.
- Buffenstein, R.; Poppitt, S.D.; McDevitt, R.M.; Prentice, A.M. Food intake and the menstrual cycle: A retrospective analysis, with implications for appetite research. Physiol. Behav. 1995, 58, 1067–1077.
- Duval, K.; Prud’homme, D.; Rabasa-Lhoret, R.; Strychar, I.; Brochu, M.; Lavoie, J.-M.; Doucet, E. Effects of the menopausal transition on dietary intake and appetite: A MONET Group Study. Eur. J. Clin. Nutr. 2014, 68, 271–276.
- Lovejoy, J.C.; Champagne, C.M.; de Jonge, L.; Xie, H.; Smith, S.R. Increased visceral fat and decreased energy expenditure during the menopausal transition. Int. J. Obes. 2008, 32, 949–958.
- Camporez, J.P.G.; Jornayvaz, F.R.; Lee, H.-Y.; Kanda, S.; Guigni, B.A.; Kahn, M.; Samuel, V.T.; Carvalho, C.R.O.; Petersen, K.F.; Jurczak, M.J.; et al. Cellular Mechanism by Which Estradiol Protects Female Ovariectomized Mice From High-Fat Diet-Induced Hepatic and Muscle Insulin Resistance. Endocrinology 2013, 154, 1021–1028.
- Day, D.S.; Gozansky, W.S.; Van Pelt, R.E.; Schwartz, R.S.; Kohrt, W.M. Sex hormone suppression reduces resting energy expenditure and -adrenergic support of resting energy expenditure. J. Clin. Endocrinol. Metab. 2005, 90, 3312–3317.
- Shea, K.L.; Gavin, K.M.; Melanson, E.L.; Gibbons, E.; Stavros, A.; Wolfe, P.; Kittelson, J.M.; Vondracek, S.F.; Schwartz, R.S.; Wierman, M.E.; et al. Body composition and bone mineral density after ovarian hormone suppression with or without estradiol treatment. Menopause 2015, 22, 1045–1052.
- Melanson, E.L.; Gavin, K.M.; Shea, K.L.; Wolfe, P.; Wierman, M.E.; Schwartz, R.S.; Kohrt, W.M. Regulation of energy expenditure by estradiol in premenopausal women. J. Appl. Physiol. 2015, 119, 975–981.
- Gavin, K.M.; Melanson, E.L.; Hildreth, K.L.; Gibbons, E.; Bessesen, D.H.; Kohrt, W.M. A Randomized Controlled Trial of Ovarian Suppression in Premenopausal Women: No Change in Free-Living Energy Expenditure. Obesity 2020, 28, 2125–2133.
- Hopkins, M.; Finlayson, G.; Duarte, C.; Gibbons, C.; Johnstone, A.M.; Whybrow, S.; Horgan, G.W.; Blundell, J.E.; Stubbs, R.J. Biological and psychological mediators of the relationships between fat mass, fat-free mass and energy intake. Int. J. Obes. 2018, 43, 233–242.
- McNeil, J.; Lamothe, G.; Cameron, J.D.; Riou, M.E.; Cadieux, S.; Lafreniere, J.; Goldfield, G.; Willbond, S.; Prud’homme, D.; Doucet, E. Investigating predictors of eating: Is resting metabolic rate really the strongest proxy of energy intake? Am. J. Clin. Nutr. 2017, 106, 1206–1212.
- Klok, M.D.; Jakobsdottir, S.; Drent, M.L. The role of leptin and ghrelin in the regulation of food intake and body weight in humans: A review. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2007, 8, 21–34.
- Hopkins, M.; Finlayson, G.; Duarte, C.; Whybrow, S.; Ritz, P.; Horgan, G.W.; Blundell, J.E.; Stubbs, R.J. Modelling the associations between fat-free mass, resting metabolic rate and energy intake in the context of total energy balance. Int. J. Obes. 2016, 40, 312–318.
- Blundell, J.E.; Caudwell, P.; Gibbons, C.; Hopkins, M.; Naslund, E.; King, N.A.; Finlayson, G. Body composition and appetite: Fat-free mass (but not fat mass or BMI) is positively associated with self-determined meal size and daily energy intake in humans. Br. J. Nutr. 2012, 107, 445–449.
- Casanova, N.; Beaulieu, K.; Oustric, P.; O’Connor, D.; Gibbons, C.; Duarte, C.; Blundell, J.; Stubbs, R.J.; Finlayson, G.; Hopkins, M. Body Fatness Influences Associations of Body Composition and Energy Expenditure with Energy Intake in Healthy Women. Obesity 2021, 29, 125–132.
- Hopkins, M.; Duarte, C.; Beaulieu, K.; Finlayson, G.; Gibbons, C.; Johnstone, A.M.; Whybrow, S.; Horgan, G.W.; Blundell, J.E.; Stubbs, R.J. Activity energy expenditure is an independent predictor of energy intake in humans. Int. J. Obes. 2019, 43, 1466–1474.
- Brown, J.C.; Cespedes Feliciano, E.M.; Caan, B.J. The evolution of body composition in oncology-epidemiology, clinical trials, and the future of patient care: Facts and numbers. J. Cachexia. Sarcopenia Muscle 2018, 9, 1200–1208.
- Dulloo, A.G.; Jacquet, J.; Girardier, L. Poststarvation hyperphagia and body fat overshooting in humans: A role for feedback signals from lean and fat tissues. Am. J. Clin. Nutr. 1997, 65, 717–723.
- Muller, M.J.; Enderle, J.; Pourhassan, M.; Braun, W.; Eggeling, B.; Lagerpusch, M.; Gluer, C.C.; Kehayias, J.J.; Kiosz, D.; Bosy-Westphal, A. Metabolic adaptation to caloric restriction and subsequent refeeding: The Minnesota Starvation Experiment revisited. Am. J. Clin. Nutr. 2015, 102, 807–819.
- Turicchi, J.; O’Driscoll, R.; Finlayson, G.; Duarte, C.; Hopkins, M.; Martins, N.; Michalowska, J.; Larsen, T.M.; van Baak, M.A.; Astrup, A.; et al. Associations between the proportion of fat-free mass loss during weight loss, changes in appetite, and subsequent weight change: Results from a randomized 2-stage dietary intervention trial. Am. J. Clin. Nutr. 2020, 111, 536–544.
- Melby, C.L.; Paris, H.L.; Sayer, R.D.; Bell, C.; Hill, J.O. Increasing Energy Flux to Maintain Diet-Induced Weight Loss. Nutrients 2019, 11, 2533.
- King, N.A.; Burley, V.J.; Blundell, J.E. Exercise-induced suppression of appetite: Effects on food intake and implications for energy balance. Eur. J. Clin. Nutr. 1994, 48, 715–724.
- Broom, D.R.; Stensel, D.J.; Bishop, N.C.; Burns, S.F.; Miyashita, M. Exercise-induced suppression of acylated ghrelin in humans. J. Appl. Physiol. 2007, 102, 2165–2171.
- Cornier, M.-A.; Melanson, E.L.; Salzberg, A.K.; Bechtell, J.L.; Tregellas, J.R. The effects of exercise on the neuronal response to food cues. Physiol. Behav. 2012, 105, 1028–1034.
- Bales, C.W.; Hawk, V.H.; Granville, E.O.; Rose, S.B.; Shields, T.; Bateman, L.; Willis, L.; Piner, L.W.; Slentz, C.A.; Houmard, J.A.; et al. Aerobic and resistance training effects on energy intake: The STRRIDE-AT/RT study. Med. Sci. Sport. Exerc. 2012, 44, 2033–2039.
- Legget, K.T.; Wylie, K.P.; Cornier, M.-A.; Melanson, E.L.; Paschall, C.J.; Tregellas, J.R. Exercise-related changes in between-network connectivity in overweight/obese adults. Physiol. Behav. 2016, 158, 60–67.
- Dorling, J.; Broom, D.R.; Burns, S.F.; Clayton, D.J.; Deighton, K.; James, L.J.; King, J.A.; Miyashita, M.; Thackray, A.E.; Batterham, R.L.; et al. Acute and Chronic Effects of Exercise on Appetite, Energy Intake, and Appetite-Related Hormones: The Modulating Effect of Adiposity, Sex, and Habitual Physical Activity. Nutrients 2018, 10, 1140.
- Westerterp, K.R. Diet induced thermogenesis. Nutr. Metab. 2004, 1, 1–5.
- Donahoo, W.T.; Levine, J.A.; Melanson, E.L. Variability in energy expenditure and its components. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 599–605.
- Demark-Wahnefried, W.; Hars, V.; Conaway, M.R.; Havlin, K.; Rimer, B.K.; McElveen, G.; Winer, E.P. Reduced rates of metabolism and decreased physical activity in breast cancer patients receiving adjuvant chemotherapy. Am. J. Clin. Nutr. 1997, 65, 1495–1501.
- Demark-Wahnefried, W.; Aziz, N.M.; Rowland, J.H.; Pinto, B.M. Riding the crest of the teachable moment: Promoting long-term health after the diagnosis of cancer. J. Clin. Oncol. 2005, 23, 5814–5830.
- Hodgkinson, K.; Butow, P.; Hunt, G.E.; Pendlebury, S.; Hobbs, K.M.; Wain, G. Breast cancer survivors’ supportive care needs 2–10 years after diagnosis. Support. Care Cancer Off. J. Multinatl. Assoc. Support. Care Cancer 2007, 15, 515–523.
- Rabin, C.; Pinto, B. Cancer-related beliefs and health behavior change among breast cancer survivors and their first-degree relatives. Psychooncology 2006, 15, 701–712.
- Maunsell, E.; Drolet, M.; Brisson, J.; Robert, J.; Deschênes, L. Dietary change after breast cancer: Extent, predictors, and relation with psychological distress. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 1017–1025.
- Corbett, T.; Cheetham, T.; Müller, A.M.; Slodkowska-Barabasz, J.; Wilde, L.; Krusche, A.; Richardson, A.; Foster, C.; Watson, E.; Little, P.; et al. Exploring cancer survivors’ views of health behaviour change: “Where do you start, where do you stop with everything?”. Psychooncology 2018, 27, 1816–1824.
- Bidstrup, P.E.; Dalton, S.O.; Christensen, J.; Tjonneland, A.; Larsen, S.B.; Karlsen, R.; Brewster, A.; Bondy, M.; Johansen, C. Changes in body mass index and alcohol and tobacco consumption among breast cancer survivors and cancer-free women: A prospective study in the Danish Diet, Cancer and Health Cohort. Acta Oncol. 2013, 52, 327–335.
- de Jonge, L.; DeLany, J.P.; Nguyen, T.; Howard, J.; Hadley, E.C.; Redman, L.M.; Ravussin, E. Validation study of energy expenditure and intake during calorie restriction using doubly labeled water and changes in body composition. Am. J. Clin. Nutr. 2007, 85, 73–79.
- Racette, S.B.; Das, S.K.; Bhapkar, M.; Hadley, E.C.; Roberts, S.B.; Ravussin, E.; Pieper, C.; DeLany, J.P.; Kraus, W.E.; Rochon, J.; et al. Approaches for quantifying energy intake and %calorie restriction during calorie restriction interventions in humans: The multicenter CALERIE study. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E441–E448.
- Thomas, D.M.; Scioletti, M.; Heymsfield, S.B. Predictive Mathematical Models of Weight Loss. Curr. Diab. Rep. 2019, 19, 93.
- Owino, V.O.; Slater, C.; Loechl, C.U. Using stable isotope techniques in nutrition assessments and tracking of global targets post-2015. Proc. Nutr. Soc. 2017, 76, 495–503.
- Patel, P.S.; Cooper, A.J.M.; O’Connell, T.C.; Kuhnle, G.G.C.; Kneale, C.K.; Mulligan, A.M.; Luben, R.N.; Brage, S.; Khaw, K.-T.; Wareham, N.J.; et al. Serum carbon and nitrogen stable isotopes as potential biomarkers of dietary intake and their relation with incident type 2 diabetes: The EPIC-Norfolk study. Am. J. Clin. Nutr. 2014, 100, 708–718.
- Franz, M.J.; VanWormer, J.J.; Crain, A.L.; Boucher, J.L.; Histon, T.; Caplan, W.; Bowman, J.D.; Pronk, N.P. Weight-loss outcomes: A systematic review and meta-analysis of weight-loss clinical trials with a minimum 1-year follow-up. J. Am. Diet. Assoc. 2007, 107, 1755–1767.
- Greenway, F.L. Physiological adaptations to weight loss and factors favouring weight regain. Int. J. Obes. 2015, 39, 1188–1196.
- Maclean, P.S.; Bergouignan, A.; Cornier, M.A.; Jackman, M.R. Biology’s response to dieting: The impetus for weight regain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, 581–600.
- Pedersen, B.; Delmar, C.; Lörincz, T.; Falkmer, U.; Grønkjær, M. Investigating Changes in Weight and Body Composition Among Women in Adjuvant Treatment for Breast Cancer: A Scoping Review. Cancer Nurs. 2019, 42, 91–105.

