Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Eleonora Cominelli | + 3485 word(s) | 3485 | 2021-09-29 08:24:08 |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Cominelli, E.; Pilu, R.; Sparvoli, F. Phytic Acid and Transporters. Encyclopedia. Available online: https://encyclopedia.pub/entry/14884 (accessed on 25 June 2026).
Cominelli E, Pilu R, Sparvoli F. Phytic Acid and Transporters. Encyclopedia. Available at: https://encyclopedia.pub/entry/14884. Accessed June 25, 2026.
Cominelli, Eleonora, Roberto Pilu, Francesca Sparvoli. "Phytic Acid and Transporters" Encyclopedia, https://encyclopedia.pub/entry/14884 (accessed June 25, 2026).
Cominelli, E., Pilu, R., & Sparvoli, F. (2021, October 08). Phytic Acid and Transporters. In Encyclopedia. https://encyclopedia.pub/entry/14884
Cominelli, Eleonora, et al. "Phytic Acid and Transporters." Encyclopedia. Web. 08 October, 2021.
Copy Citation
Phytic acid has two main roles in plant tissues: storage of phosphorus and regulation of different cellular processes. From a nutritional point of view, it is considered an antinutritional compound because, being a cation chelator, its presence reduces mineral bioavailability from the diet. In recent decades, the development of low phytic acid (lpa) mutants has been an important goal for nutritional seed quality improvement, mainly in cereals and legumes. Different lpa mutations affect phytic acid biosynthetic genes. However, other lpa mutations isolated so far, affect genes coding for three classes of transporters: a specific group of ABCC type vacuolar transporters, putative sulfate transporters, and phosphate transporters.
phytic acid
low phytic acid mutants
MRP transporter
ABCC transporter
SULTR transporter
Pht
phosphate transporter
sulfate transporter
1. Introduction
Phytic acid (PA), chemically myo-inositol-1,2,3,4,5,5-hexakisphosphate, is the major form of phosphorus (P) storage in seeds (up to 85% of total P) and in other plant organs, such as pollen, roots, tubers, and turions. However, PA is not only an important molecule for P storage but, together with its precursors (lower InsPs and myo-inositol) and its derivative molecules (InsP7 and InsP8 inositol pyrophosphates), it is involved in the regulation of different cell signaling and plant processes in vegetative tissues, such as abiotic and biotic stress response, storage and polar transport of auxin, P homeostasis, photomorphogenesis, chromatin modification, and remodeling and mRNA nuclear export [1].
In seeds, where P amounts may even be 1000-fold higher than those detected in vegetative tissues, PA is accumulated during development, reaching a plateau at the end of the “cell expansion phase” [2][3]. PA is synthesized in the cytosol through two different routes: (i) The lipid-independent pathway, the most used in the seed, consisting of the sequential phosphorylation of the 6-carbon myo-inositol and soluble inositol phosphates (InsPs), and (ii) the lipid-dependent pathway, using precursors that include phosphatidylinositol (PtdIns) and PtdIns phosphates. PA is transferred from the cytosol to the vacuole where it is accumulated into globoids, spherical inclusions found within protein bodies [4][5][6][7]. Interestingly, the amount and distribution of PA in different seed/grain portions vary among different species. In cereals, there are differences between Zea mays L. (maize) kernels, where PA is mainly present in the embryo and scutellum, and the small grains of Hordeum vulgare L. (barley), different Triticum (wheat) species and Oryza sativa L. (rice), where 80% of PA is stored in the aleurone and bran (maternal teguments) and only a limited amount accumulates in the embryo [8]. However, in legumes, more than 95% seed PA is accumulated in the cotyledons [9]. During germination, phytases degrade PA and in this way, P is remobilized to support seedling growth [10]. Due to its high negative charge at physiological pH (~6–7), PA easily precipitates in the form of phytate salts binding cations, such as iron, zinc, potassium, calcium, magnesium, some of them (mainly iron and zinc) important from a nutritional point of view, in this way reducing their bioavailability. Only ruminants are able to degrade PA, due to the presence of microbial phytases in their digestive tracts, while for monogastric animals, including humans, mainly in those populations whose diet is largely based on staple crops, the presence of PA decreases seeds’ nutritional value [11][12]. Moreover, as undigested PA is excreted by non-ruminants, such as swine, fowl, and fish, the supplementation of feed with nutrient P is a common practice, in order to provide for an animal’s nutritional requirement. In this way P concentrations increase in manure, consequently in soils, finally contributing to P pollution in runoff water [13]. Hence, PA is considered an antinutrient and in recent decades, many efforts were spent to isolate and develop low phytic acid (lpa) crops, in which a 45–90% reduction of PA was achieved [1]. Unfortunately, it was shown that the reduction in PA content may affect plant growth, plant stress response and seed development and germination, thus limiting the efficacy of the introgression of the lpa trait into breeding programs [14]. The negative pleiotropic effects of the lpa mutations depend on the previously mentioned important roles PA has in different regulatory processes.
Hence, it is very important to identify the best strategy in order to specifically decrease PA content in the seeds without affecting plant and seed performance and possibly contribute to reducing the environmental impact. The lpa mutations so far isolated can be classified into three classes, depending on the step of the biosynthetic pathway or transport they affect: (i) Mutations altering the MIPS activity, the first steps of the biosynthetic pathway (from glucose 6-P to myo-inositol[3]-monophosphate), (ii) mutations affecting the following phosphorylation of the InsP6 pathway (from myo-inositol[3]-monophosphate to PA), (iii) mutations perturbing the final transport of PA.
Only mutants belonging to class (ii) accumulate InsPs intermediates. The mutations belonging to the (i) and (iii) classes induce a decrease of PA amount, accompanied by a molar equivalent increase of inorganic phosphate (Pi) in the homozygous mutants. Moreover, they are usually perturbed in different branches of the biosynthetic pathway common to PA and other compounds (e.g., galactinol, raffinose, stachyose, and ascorbic acid). Mutants in three classes of transporters have been characterized for their lpa phenotype, affected in: (i) A specific group of ABCC type vacuolar transporters [15], orthologues to the Arabidopsis thaliana (L.) Heinh AtMRP5 (also referred to as AtABCC5) [16][17], also known as multidrug resistance-associated proteins (MRPs), (ii) putative sulfate transporters, orthologues of the Arabidopsis AtSULTR3;3 [18][19] and AtSULTR3;4 proteins [20]; (iii) the rice OsPht1;4 phosphate transporter [21][22]. Only in the case of an ABCC transporter it was shown that the protein is able to actually transport PA [16], while the cases of SUTR3;3/SULTR3;4 and Pht1;4 transporters it is not yet clear the role of these proteins in controlling phytic acid homeostasis.
2. PA-MRP Transporters
MRP proteins are transmembrane transporters involved in several functions, such as organic ions transport, xenobiotic detoxification, oxidative stress tolerance, and transpiration control [23]. The first evidence of the involvement of an MRP-type ABC transporter in PA transport was reported for the maize ZmMRP4 protein from the analysis of the insertional lpa1 mutant [24]. ZmMRP4 gene is orthologous to AtMRP5, which had already been characterized some years ago as an anion transporter involved in root growth, lateral root formation, regulation of stomatal movement, guard cell hormonal signaling, and water use efficiency [25][26][27], aspects not immediately attributable to PA transport. The biochemical demonstration that PA transport was dependent on the presence of a functional MRP transporter in an ATP-dependent manner was given for AtMRP5, showing a very high affinity for PA (maximum reaction velocity -Vmax- values of about 1.6–2.5 µmol min−1 mg−1 and Michaelis—Mentent constant -Km- ranging between 263 and 310 nM) and a vacuolar subcellular localization [16].
As summarized in Table 1 and Figure S2, other PA-MRP genes and the corresponding mutants/transgenics have been hereafter characterized in rice, Glycine max (L.) Merr. (soybean), Phaseolus vulgaris L. (common bean) and Triticum aestivum L. (soft wheat) [28][29][30][31][32][33]. Moreover, the putative Pennisetum glaucum (L.) R.Br. (pearl millet) PA-MRP gene has recently been described [34].
Table 1. Described cereal and legume transporters involved in PA accumulation and corresponding mutations affecting PA seed/grain content. EMS stands for ethyl methanesulfonate. * Indicates incomplete sequence.
| Class of Transporters | Species | Gene | Phytozome/Genbank/Ensembl Accession Number | Origin of Mutation | Mutation | Reference |
|---|---|---|---|---|---|---|
| MRP | Zea mays | ZmMRP4/ZmABCC4 | EF586878 | EMS | lpa1-1 | [35] |
| lpa1-241 | [36] | |||||
| lpa1-7 | [37] | |||||
| T-DNA insertion | lpa1-mum1 | [24] | ||||
| Embryo specific:RNAi | Ole::MRP4 Glb::MRP4 | [24] | ||||
| Oryza sativa | OsMRP5/OsABCC13 | LOC_Os03g04920 | γ rays + sodium azide | Os-lpa-XS110-2 | [38] | |
| Os-lpa-XS110-3 | [39] | |||||
| T-DNA insertion | 4A-02500 | [39] | ||||
| Embryo specific amiRNA | Ami-MRP5 | [40] | ||||
| Triticum aestivum | TaABCC13-4B | TraesCS4B02G343800 | Constitutive RNAi | TaABCC13 RNAi | [33] | |
| TaABCC13-4D | TraesCS4D02G339000 | |||||
| TaABCC13-5A* | TraesCS5A02G512500 | |||||
| Glycine max | GmMRP3/GmABCC1 | Glyma.03G167800 | EMS | CX1834 | [29][30][31][41] | |
| GmMRP19/GmABCC2 | Glyma.19G169000 | |||||
| GmMRP13/GmABCC3 | Glyma.13G127500 | no reported mutant | no reported mutant | [32] | ||
| Phaseolus vulgaris | PvMRP1/PvABCC1 | Phvul.001G165500 | EMS | lpa1 | [32][42] | |
| lpa12 | [43] | |||||
| PvMRP2/PvABCC2 | Phvul.007G153800 | no reported mutant | no reported mutant | [32] | ||
| SULTR | Oryza sativa | OsSULTR3;3 | LOC_Os04g55800 | γ rays | Oslpa-MH86-1 Os-lpa-Z9B-1 |
[28] [19] |
| OsSULTR3;4 | LOC_Os06g05160 | retrotransposon Tos-17 insertion | spdt-1, spdt-2, spdt-3 | [20] | ||
| Hordeum vulgare | Hvst | HORVU2Hr1G113050 | sodium azide | lpa1-1(M422) | [44] [19] |
|
| Pht | Oryza sativa | OsPht1;4 | LOC_Os04g10750 | retrotransposon Tos-17 insertion | ospt4-1 (NE1260) ospt4-2 (SHIP_ZSF6267) RNAi |
[21] [22] |
The main difference between cereals and legumes for which PA-MRPs have been characterized so far is the gene number: While only one gene is present in diploid maize, pearl millet, and rice genomes, and three copies in the hexaploid Triticum aestivum, two and three paralogues are present in common bean and soybean, respectively [17][32][33]. The presence of more than one member of the PA-MRP genes seems to be a common feature of legumes, for example also in Medicago truncatula it is possible to predict two PA-MRP genes [43], unlike the situation in other dicotyledons, such as Arabidopsis and Solanum lycopersicum L. (tomato) in which only one PA-MRP gene was described [16][45]. As discussed below, the gene copy number has a significant influence on the lpa mutant phenotypes.
The gene structure of PA-MRP transporters is very highly conserved: All analyzed genes in the present study have 11 exons and 10 introns with very similar lengths, only some differences can be found in the lengths of some introns between cereals and legumes. In Figure 1a, the rice OsMRP5 and the soybean GmABCC1 gene schematic representations are given as examples.
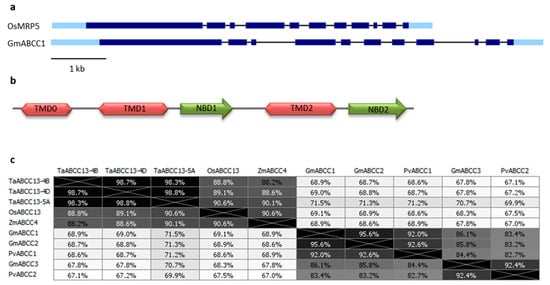
Figure 1. (a) Gene structure of OsMRP5 and GmABCC1 genes, as an example of a cereal and a legume PA-MRP gene, respectively. Light and dark blue rectangles represent UTRs and coding exons, respectively, the black bars correspond to introns. Gene Structure Display Server [46] was used; (b) Predicted domains of the PA-MRP protein. The transmembrane domains (TMD) and the nucleotide-binding domains (NBD) are represented in red and green, respectively. The structure of the PA-MRP proteins was previously described [17]; (c) Distances between PA-MRP proteins, expressed as a percentage of identity. Phylogenies were constructed with the Geneious Tree Builder tool, using the Jukes—Cantor distance model, neighbor-joining tree build method.
PA-MRP proteins are full-length ABC transporters (length from 1501 aa of TaABCC13–4B to 1539 aa of GmABCC1) with three membrane-spanning domains (TMD0, containing five transmembrane α-helices and TMD1, TMD2, each with six α-helices) and two cytosolic nucleotide-binding domains (NBD1 and NBD2, containing the Walker A and B motifs), arranged in the TMD0-TMD1-NBD1_TMD2-NBD2 so-called forward orientation (Figure 1b), as previously described [17]. Although it is not known which specific amino acids are involved in PA transport, a conserved lysine residues stretch, located in the cytosolic loop linking NBD1 and TMD2 and a number of charged amino acid residues (mostly lysine and arginine) found in other conserved stretches in TMD1 and TMD2 have been suggested to be involved in PA transport [17].
PA-MRP protein sequences are phylogenetically very highly conserved among different species, mainly in the TMD and NBD domains, but also outside, particularly among cereals or legumes (Figure 1 and Figure S1). As shown in Figure 1c, the degree of aminoacid identity among different PA-MRP proteins belonging to different species is very high, also between cereals and legumes where it ranges from 67 to 71.5% (similarity between cereals and legumes ranges from 86 to 89%, data not shown).
In Figure 2, pictographic representations of the different organs’ expression patterns of the OsMRP5 and the soybean PA-MRP genes, taken as examples for cereals and legumes, are reported from the rice and soybean eFP Browsers [47]. The rice gene is expressed at high levels in different organs including the caryopsis, as previously reported [39]. The maize ortholog shows a similar expression pattern [24]. The TaABCC13 genes are expressed in different plant organs, preferentially during grain developmental stages, with the transcript accumulation derived from the B genome the highest one, mainly at 14 days after anthesis [33][48].
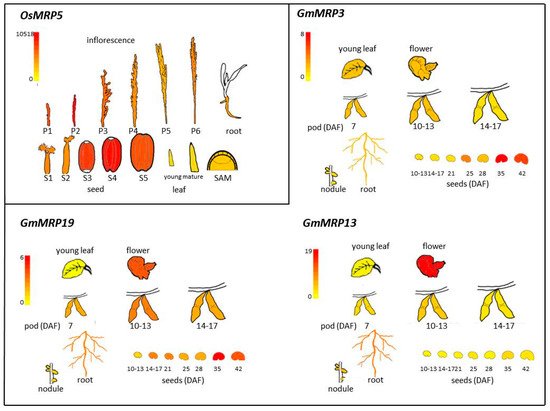
Figure 2. PA-MRP gene expression data in various rice and soybean organs and tissues were obtained from the rice and soybean eFP Browsers [47]. Rice MAS and soybean Severin data sources were used. For rice, the default signal threshold was used, while for the three soybean genes the signal threshold was arbitrarily put to the same value (8.00) in order to compare expression data between different genes.
The expression pattern of the different legume genes varies. As shown in Figure 2, GmMRP3 and GmMRP19 genes are expressed in different organs and highly expressed in seed, particularly at the late stage of development, while the GmMRP13 gene is mainly expressed in root and flower and at a very low level in seed. As discussed below, when both GmMRP3 and GmMRP19 are mutated an lpa seed phenotype occurs [30]. It suggests that GmMRP3 and GmMRP19 transporters have an important role in PA accumulation and their function is redundant, while GmMRP13 is not active in the seed.
A similar diversified expression pattern was observed in common bean, where the PvMRP1 gene, coding for a protein more similar to GmMRP3 and GmMRP19 (Figure 1c and Figure S1), is highly expressed in cotyledons, where its transcript levels continue to increase during seed development, reaching the highest levels at 28 days after flowering (DAF) with a similar kinetics to that reported for the accumulation of PA in the same organ. The PvMRP2 gene, ortholog of GmMRP13, is expressed similarly to PvMRP1 in vegetative organs, but at no appreciable level in cotyledons. Interestingly, both genes are expressed in root nodules, organs specialized in symbiosis with nitrogen-fixing bacteria, in which the role of PA is still unknown [43]. Recently, a detailed analysis was reported of GUS activity in Arabidopsis thaliana and Medicago truncatula plants, harboring a promoter sequence of PvMRP1 and PvMRP2 genes, fused upstream of the GUS reporter gene. The strongest GUS activity, driven by both constructs, in organs other than the seeds was present in the vascular tissues [43]. Similar patterns of reporter gene activity were previously shown in transgenic plants harboring the AtMRP5 promoter [25] and promoters of different Arabidopsis genes coding for enzymes involved in different steps in PA pathway [49][50][51][52][53][54][55][56]. These data suggest that vascular tissues are an important site for synthesis and transport of PA involved in the regulation of different cellular processes, the so-called “signaling PA” [57].

3. SULTR3.3 and SULTR3.4 Transporters Involved in PA Metabolism
Two lpa mutants isolated in barley and rice are affected in HvST and OsSULTR3;3 genes, respectively [19][28][44][58], coding for two putative sulfate transporters, belonging to the SULTR3;3 class [18]. Recently, another rice lpa mutant, affected in the OsSULTR3;4 putative sulfate transporter, also called SULTR-like Phosphorus Distribution Transporter (SPDT), was isolated [20].
Here, we present an in silico analysis of SULTR3;3 and SULTR3;4 genes, including the ones already described (Table 1) and also putative SULTR3;3 and SULTR3;4 orthologs from other cereal and legume crops for which interest in the isolation of lpa mutants is considered an important challenge, such as maize, barley, common bean and soybean (Table 2). A phylogenetic tree with all the SULTR3;3 and SULTR3;4 proteins of cereals and legumes analyzed in the present work is shown in Figure S3.
Table 2. Putative orthologous genes of OsSULTR3;3/HvSULTR3;3 and OsSULTR3;4, identified in maize, common bean, and soybean by in silico analysis.
| SULTR Group. | Species | Gene Name | Phytozome Accession Number |
|---|---|---|---|
| SULTR3;3 | Zea mays | ZmSULTR3;3 | GRMZM2G395114 |
| Phaseolus vulgaris | PvSULTR3;3 | Phvul.002G095300 | |
| Glycine max | GmSULTR3;3a | Glyma.20G017100 | |
| GmSULTR3;3b | Glyma.07G218900 | ||
| SULTR3;4 | Zea mays | ZmSULTR3;4 | GRMZM2G444801 |
| Phaseolus vulgaris | PvSULTR3;4a | Phvul.005G171800 | |
| PvSULTR3;4b | Phvul.010G151000 | ||
| Glycine max | GmSULTR3;4a | Glyma.07G006500 | |
| GmSULTR3;4b | Glyma.08G207100 | ||
| GmSULTR3;4c | Glyma.13G360000 | ||
| GmSULTR3;4d | Glyma.15G014000 |
3.1. SULTR3;3
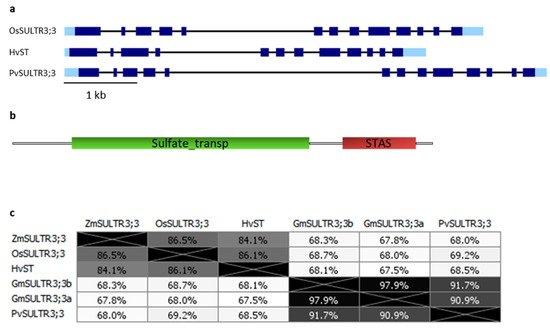
In all analyzed species, one putative ortholog belonging to the SULTR3;3 group was found by BLAST analysis of HvST or OsSULTR3;3 proteins against the different genomes, except for soybean, in which two different genes have been identified (Table 2, Figures S3 and S4). Indeed, this is not unexpected, as soybean underwent an ancient event of genome duplication [59]. The SULTR3;3 gene structure is quite conserved among species and consists of 13 exons in the majority of the genes, with the exceptions of barley and maize with only 12 exons. All genomic sequences are characterized by the presence of a long fourth or fifth intron, as reported in Figure 3a, where the structure of the characterized OsSULTR3;3 and HvST and of PvSULTR3;3 is shown as an example.
Figure 3. (a) Gene structure of OsSULTR3;3, HvST and putative PvSULTR3;3 genes. Light and dark blue rectangles represent UTRs and coding exons, respectively, the black bars correspond to introns. Table 1. a legend. (b) Predicted domains of the SULTR3;3 protein. The sulfate transporter and the anti-sigma factor antagonist (STAS) domains are represented in red and green, respectively. Picture reproduced from [58]. (c) Distances between SULTR3;3 proteins, expressed as the percentage of identity. Phylogenies were constructed as described in Figure 1c.
Predicted domains of SULTR3;3 proteins are represented in Figure 3b and correspond to a sulfate transporter domain and an anti-sigma factor antagonist (STAS) domain, as previously reported [58]. Protein length varies from 647 aa of PvSULTR3;3 to 661 aa of OsSULTR3;3.
Protein identity is generally very high among different species, ranging from 84.1% to 86.5% among the considered cereals and from 90.9% to 91.7% among legumes, and at 97.9% in the two paralogs of soybean, as shown in the Figure 3c diagram.
In the case of the OsSULTR3;3 gene detailed expression analysis was reported: Transgenic lines harboring the promoter of this gene fused to the GUS reporter gene revealed that a strong GUS activity was present in vascular bundles of shoots, leaves, flowers, and grains, where it was mainly detected in the scutellum. Moreover, the subcellular localization was defined to be in the endoplasmic reticulum [58]. Interestingly, both GmSULTR3;3a and GmSULTR3;3b are expressed in leaves and flowers, while only GmSULTR3;3a was expressed in the seed, with an increasing expression during seed development with a peak at 35 DAF (data not shown, in silico analysis performed using the soybean eFP Browser [47]).
The exact function of this family of proteins is still unknown and in the case of OsSULTR3;3, which was the only one analyzed in detail, no activity was revealed for the transport of phosphate, sulfate, inositol or inositol 1,4,5 triphosphate by heterologous expression in either yeast or Xenopus oocytes [58].
3.2. SULTR3;4
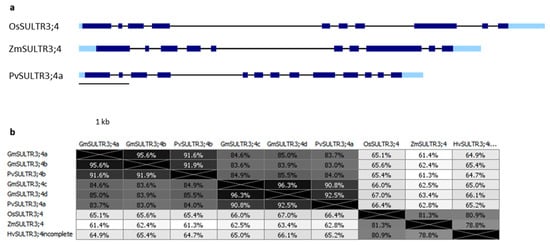
In the case of the SULTR3;4 group of transporters a similar situation to the one previously described for MRP proteins is present: In cereals, only one protein for each species can be found by BLAST analysis of OsSULTR3;4 against the different genomes, while in legumes, two or four paralogous proteins are present in common bean and soybean, respectively (Table 2 and Figures S3 and S5). The gene structure differs between cereals with 10 exons (the barley sequence present in the Phytozome database is incomplete with only eight exons) and legumes with 13 exons and also in this case, the fourth (the fifth in maize) intron is quite long. In Figure 4a the structures of the characterized OsSULTR3;4 and of putative ZmSULTR3;4 and PvSULTR3;4a genes are given as examples.
Figure 4. (a) Gene structure of OsSULTR3;4 and putative ZmSULTR3;4 and PvSULTR3;4a genes. Light and dark blue rectangles represent UTRs and coding exons, respectively, the black bars correspond to introns. The gene structure was obtained as described in Figure 1a legend. (b) Distances between SULTR3;4 proteins, expressed as the percentage of identity. Phylogenies were constructed as described in Figure 1c.
Predicted domains are the same as those already described for the SULTR3;3 protein, represented in Figure 3b. Protein length varies from 648 aa of the soybean protein to 670 aa of the rice one (Figure S5).
Also for SULTR3;4 proteins, identity is quite high ranging from 78.8% to 81.3% among cereals, from 83% to 91.9% among legumes and from 61.3% to 67% between cereals and legumes, as shown in the diagram in Figure 4b.
Analysis of the phylogenetic tree (Figure S3) clearly shows a separation between monocotyledons and dicotyledons. Furthermore, in the two legume species, the gene is duplicated, with soybean carrying four genes arising from an ancient event of genome duplication [59].
qRT-PCR expression analysis of OsSULTR3;4 gene revealed that during grain filling it was mainly expressed in node I, a very important hub for mineral distribution to upper node and panicle in Poaceae [60]. Moreover, immunostaining against GFP in lines harboring OsSULTR3;4 promoter fused to GFP, showed the highest staining in the xylem region of both enlarged- and diffuse-vascular bundles of the basal node and in node I, as well as in the parenchyma tissues between them, but not in the phloem region [20]. The activity of OsSULTR3;4 as an influx plasma-membrane localized H+/Pi symporter was shown in proteoliposomes as well as in Xenopus oocytes. Particularly, it was found that OsSULTR3;4 is involved in the intervascular transfer of P at the nodes, unloading P from xylem towards phloem [20].
4. OsPht1;4 Phosphate Transporter
The OsPht1;4 (or OsPT4, corresponding to LOC_Os04g10750) phosphate transporter, belonging to the Pht1 family, was described as influencing grain PA content, as the corresponding mutant produces lpa grains [21]. As the identification of putative orthologues of this protein in other species is not so obvious, due to the high number of Pht1 genes and to their sequence similarity, (in rice there are 26 [61]), we limit our consideration to OsPT4.
The genomic sequence is characterized by the presence of a single exon (Figure 5a) and the protein, 538 aa long, by a major facilitator superfamily domain, characteristic of different transporters, including phosphate transporters (Figure 5b).The OsPT4 gene is mainly expressed in roots, flag leaves and embryos, and its expression is increased in response to prolonged P starvation conditions in shoots and roots, where the signal is specifically localized to the exodermis. The protein is localized to the plasma membrane, as shown in the protoplast system and it is a functional Pi influx transporter, able to complement a yeast mutant defective in Pi uptake and to facilitate the increased accumulation of Pi in Xenopus oocytes.
The genomic sequence is characterized by the presence of a single exon (Figure 5a) and the protein, 538 aa long, by a major facilitator superfamily domain, characteristic of different transporters, including phosphate transporters (Figure 5b).The OsPT4 gene is mainly expressed in roots, flag leaves and embryos, and its expression is increased in response to prolonged P starvation conditions in shoots and roots, where the signal is specifically localized to the exodermis. The protein is localized to the plasma membrane, as shown in the protoplast system and it is a functional Pi influx transporter, able to complement a yeast mutant defective in Pi uptake and to facilitate the increased accumulation of Pi in Xenopus oocytes.
Figure 5. (a) Gene structure of OsPHT1;4. Light and dark blue rectangles represent UTRs and coding exons, respectively, the black bars correspond to introns. The gene structure was obtained as described in Figure 1a legend. (b) Predicted domain of the OsPHT1;4 protein by PFAM software. The major facilitator superfamily (MFS) domain is represented.
References
- Sparvoli, F.; Cominelli, E. Seed biofortification and phytic acid reduction: A conflict of interest for the plant? Plants 2015, 4, 728–755.
- Coelho, C.; Tsai, S.; Vitorello, V. Dynamics of inositol phosphate pools (tris-, tetrakis- and pentakisphosphate) in relation to the rate of phytate synthesis during seed development in common bean (Phaseolus vulgaris). J. Plant Physiol. 2005, 162, 1–9.
- Hatzack, F.; Johansen, K.; Rasmussen, S. Nutritionally relevant parameters in low-phytate barley (Hordeum vulgare L.) grain mutants. J. Agric. Food Chem. 2000, 48, 6074–6080.
- Lin, L.; Ockenden, I.; Lott, J. The concentrations and distribution of phytic acid-phosphorus and other mineral nutrients in wild-type and low phytic acid1-1 (lpa1-1) corn (Zea mays L.) grains and grain parts. Can. J. Bot. 2005, 83, 131–141.
- Ockenden, I.; Dorsch, J.; Reid, M.; Lin, L.; Grant, L.; Raboy, V.; Lott, J. Characterization of the storage of phosphorus, inositol phosphate and cations in grain tissues of four barley (Hordeum vulgare L.) low phytic acid genotypes. Plant Sci. 2004, 167, 1131–1142.
- Regvar, M.; Eichert, D.; Kaulich, B.; Gianoncelli, A.; Pongrac, P.; Vogel-Mikus, K.; Kreft, I. New insights into globoids of protein storage vacuoles in wheat aleurone using synchrotron soft X-ray microscopy. J. Exp. Bot. 2011, 62, 3929–3939.
- Krishnan, H. Preparative procedures markedly influence the appearance and structural integrity of protein storage vacuoles in soybean seeds. J. Agric. Food Chem. 2008, 56, 2907–2912.
- O’Dell, B.L.; de Boland, A.R.; Koirtyohann, S.T. Distribution of phytate and nutritionally important elements among the morphological components of cereal grains. J. Agric. Food Chem. 1972, 20, 718–721.
- Ariza-Nieto, M.; Blair, M.; Welch, R.; Glahn, R. Screening of iron bioavailability patterns in eight bean (Phaseolus vulgaris L.) genotypes using the caco-2 cell in vitro model. J. Agric. Food Chem. 2007, 55, 7950–7956.
- Raboy, V. myo-Inositol-1,2,3,4,5,6-hexakisphosphate. Phytochemistry 2003, 64, 1033–1043.
- Raboy, V. Seeds for a better future: ‘low phytate’, grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 2001, 6, 458–462.
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53 (Suppl. 2), S330–S375.
- Leytem, A.B.; Maguire, R.O. Environmental implications of inositol phosphates in animal manures. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CAB International: Wallingford, CT, USA; Oxfordshire, UK, 2007; pp. 150–168.
- Raboy, V. Approaches and challenges to engineering seed phytate and total phosphorus. Plant Sci. 2009, 177, 281–296.
- Martinoia, E. Vacuolar transporters—Companions on a longtime journey. Plant Physiol. 2018, 176, 1384–1407.
- Nagy, R.; Grob, H.; Weder, B.; Green, P.; Klein, M.; Frelet-Barrand, A.; Schjoerring, J.; Brearley, C.; Martinoia, E. The Arabidopsis ATP-binding cassette protein AtMRP5/AtABCC5 is a high affinity inositol hexakisphosphate transporter involved in guard cell signaling and phytate storage. J. Biol. Chem. 2009, 284, 33614–33622.
- Colombo, F.; Paolo, D.; Cominelli, E.; Sparvoli, F.; Nielsen, E.; Pilu R. MRP transporters and low phytic acid mutants in major crops: main pleiotropic effects and future perspectives. Front Plant Sci. 2020, 11,1301. doi: 10.3389/fpls.2020.01301. eCollection 2020.
- Takahashi, H.; Buchner, P.; Yoshimoto, N.; Hawkesford, M.J.; Shiu, S.H. Evolutionary relationships and functional diversity of plant sulfate transporters. Front. Plant Sci. 2011, 2, 119.
- Ye, H.; Zhang, X.; Broughton, S.; Westcott, S.; Wu, D.; Lance, R.; Li, C. A nonsense mutation in a putative sulphate transporter gene results in low phytic acid in barley. Funct. Integr. Genom. 2011, 11, 103–110.
- Yamaji, N.; Takemoto, Y.; Miyaji, T.; Mitani-Ueno, N.; Yoshida, K.T.; Ma, J.F. Reducing phosphorus accumulation in rice grains with an impaired transporter in the node. Nature 2017, 541, 92–95.
- Zhang, F.; Sun, Y.; Pei, W.; Jain, A.; Sun, R.; Cao, Y.; Wu, X.; Jiang, T.; Zhang, L.; Fan, X.; et al. Involvement of OsPht1;4 in phosphate acquisition and mobilization facilitates embryo development in rice. Plant J. 2015, 82, 556–569.
- Ye, Y.; Yuan, J.; Chang, X.; Yang, M.; Zhang, L.; Lu, K.; Lian, X. The phosphate transporter gene OsPht1;4 is involved in phosphate homeostasis in rice. PLoS ONE 2015, 10, e0126186.
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 338–355.
- Shi, J.; Wang, H.; Schellin, K.; Li, B.; Faller, M.; Stoop, J.; Meeley, R.; Ertl, D.; Ranch, J.; Glassman, K. Embryo-specific silencing of a transporter reduces phytic acid content of maize and soybean seeds. Nat. Biotechnol. 2007, 25, 930–937.
- Gaedeke, N.; Klein, M.; Kolukisaoglu, U.; Forestier, C.; Muller, A.; Ansorge, M.; Becker, D.; Mamnun, Y.; Kuchler, K.; Schulz, B.; et al. The Arabidopsis thaliana ABC transporter AtMRP5 controls root development and stomata movement. EMBO J. 2001, 20, 1875–1887.
- Klein, M.; Perfus-Barbeoch, L.; Frelet, A.; Gaedeke, N.; Reinhardt, D.; Mueller-Roeber, B.; Martinoia, E.; Forestier, C. The plant multidrug resistance ABC transporter AtMRP5 is involved in guard cell hormonal signalling and water use. Plant J. 2003, 33, 119–129.
- Suh, S.J.; Wang, Y.F.; Frelet, A.; Leonhardt, N.; Klein, M.; Forestier, C.; Mueller-Roeber, B.; Cho, M.H.; Martinoia, E.; Schroeder, J.I. The ATP binding cassette transporter AtMRP5 modulates anion and calcium channel activities in Arabidopsis guard cells. J. Biol. Chem. 2007, 282, 1916–1924.
- Liu, Q.; Xu, X.; Ren, X.; Fu, H.; Wu, D.; Shu, Q. Generation and characterization of low phytic acid germplasm in rice (Oryza sativa L.). Theor. Appl. Genet. 2007, 114, 803–814.
- Maroof, M.; Glover, N.; Biyashev, R.; Buss, G.; Grabau, E. Genetic basis of the low-phytate trait in the soybean line CX1834. Crop Sci. 2009, 49, 69–76.
- Gillman, J.; Pantalone, V.; Bilyeu, K. The low phytic acid phenotype in soybean line CX1834 is due to mutations in two homologs of the maize low phytic acid gene. Plant Genome 2009, 2, 179–190.
- Gillman, J.; Baxter, I.; Bilyeu, K. Phosphorus partitioning of soybean lines containing different mutant alleles of two soybean seed-specific adenosine triphosphate-binding cassette phytic acid transporter paralogs. Plant Genome 2013, 6.
- Panzeri, D.; Cassani, E.; Doria, E.; Tagliabue, G.; Forti, L.; Campion, B.; Bollini, R.; Brearley, C.A.; Pilu, R.; Nielsen, E.; et al. A defective ABC transporter of the MRP family, responsible for the bean lpa1 mutation, affects the regulation of the phytic acid pathway, reduces seed myo-inositol and alters ABA sensitivity. New Phytol. 2011, 191, 70–83.
- Bhati, K.K.; Alok, A.; Kumar, A.; Kaur, J.; Tiwari, S.; Pandey, A.K. Silencing of ABCC13 transporter in wheat reveals its involvement in grain development, phytic acid accumulation and lateral root formation. J. Exp. Bot. 2016, 67, 4379–4389.
- Boncompagni, E.; Orozco-Arroyo, G.; Cominelli, E.; Gangashetty, P.I.; Grando, S.; Kwaku Zu, T.T.; Daminati, M.G.; Nielsen, E.; Sparvoli, F. Antinutritional factors in pearl millet grains: Phytate and goitrogens content variability and molecular characterization of genes involved in their pathways. PLoS ONE 2018, 13, e0198394.
- Raboy, V.; Gerbasi, P.F.; Young, K.A.; Stoneberg, S.D.; Pickett, S.G.; Bauman, A.T.; Murthy, P.P.; Sheridan, W.F.; Ertl, D.S. Origin and seed phenotype of maize low phytic acid 1-1 and low phytic acid 2-1. Plant Physiol. 2000, 124, 355–368.
- Pilu, R.; Panzeri, D.; Gavazzi, G.; Rasmussen, S.K.; Consonni, G.; Nielsen, E. Phenotypic, genetic and molecular characterization of a maize low phytic acid mutant (lpa241). Theor. Appl. Genet. 2003, 107, 980–987.
- Cerino Badone, F.; Amelotti, M.; Cassani, E.; Pilu, R. Study of Low Phytic Acid1-7 (lpa1-7), a New ZmMRP4 Mutation in Maize. J. Hered. 2012, 103, 598–605.
- Liu, K.; Peterson, K.; Raboy, V. Comparison of the phosphorus and mineral concentrations in bran and abraded kernel fractions of a normal barley (Hordeum vulgare) cultivar versus four low phytic acid isolines. J. Agric. Food Chem. 2007, 55, 4453–4460.
- Xu, X.; Zhao, H.; Liu, Q.; Frank, T.; Engel, K.; An, G.; Shu, Q. Mutations of the multi-drug resistance-associated protein ABC transporter gene 5 result in reduction of phytic acid in rice seeds. Theor. Appl. Genet. 2009, 119, 75–83.
- Li, W.; Zhao, H.; Pang, W.; Cui, H.; Poirier, Y.; Shu, Q. Seed-specific silencing of OsMRP5 reduces seed phytic acid and weight in rice. Transgenic Res. 2014, 23, 585–599.
- Wilcox, J.; Premachandra, G.; Young, K.; Raboy, V. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop Sci. 2000, 40, 1601–1605.
- Campion, B.; Sparvoli, F.; Doria, E.; Tagliabue, G.; Galasso, I.; Fileppi, M.; Bollini, R.; Nielsen, E. Isolation and characterisation of an lpa (low phytic acid) mutant in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2009, 118, 1211–1221.
- Cominelli, E.; Confalonieri, M.; Carlessi, M.; Cortinovis, G.; Daminati, M.G.; Porch, T.G.; Losa, A.; Sparvoli, F. Phytic acid transport in Phaseolus vulgaris: A new low phytic acid mutant in the PvMRP1 gene and study of the PvMRPs promoters in two different plant systems. Plant Sci. 2018, 270, 1–12.
- Zhao, H.; Liu, Q.; Ren, X.; Wu, D.; Shu, Q. Gene identification and allele-specific marker development for two allelic low phytic acid mutations in rice (Oryza sativa L.). Mol. Breed. 2008, 22, 603–612.
- Ofori, P.A.; Mizuno, A.; Suzuki, M.; Martinoia, E.; Reuscher, S.; Aoki, K.; Shibata, D.; Otagaki, S.; Matsumoto, S.; Shiratake, K. Genome-wide analysis of ATP binding cassette (ABC) transporters in tomato. PLoS ONE 2018, 13, e0200854.
- Gene Structure Display Server. Available online: http://gsds.cbi.pku.edu.cn/ (accessed on 20 November 2019).
- Patel, R.; Nahal, H.; Breit, R.; Provart, N. BAR expressolog identification: Expression profile similarity ranking of homologous genes in plant species. Plant J. 2012, 71, 1038–1050.
- Bhati, K.; Aggarwal, S.; Sharma, S.; Mantri, S.; Singh, S.; Bhalla, S.; Kaur, J.; Tiwari, S.; Roy, J.; Tuli, R.; et al. Differential expression of structural genes for the late phase of phytic acid biosynthesis in developing seeds of wheat (Triticum aestivum L.). Plant Sci. 2014, 224, 74–85.
- Latrasse, D.; Jegu, T.; Meng, P.; Mazubert, C.; Hudik, E.; Delarue, M.; Charon, C.; Crespi, M.; Hirt, H.; Raynaud, C.; et al. Dual function of MIPS1 as a metabolic enzyme and transcriptional regulator. Nucleic Acids Res. 2013, 41, 2907–2917.
- Donahue, J.; Alford, S.; Torabinejad, J.; Kerwin, R.; Nourbakhsh, A.; Ray, W.; Hernick, M.; Huang, X.; Lyons, B.; Hein, P.; et al. The Arabidopsis thaliana myo-inositol 1-phosphate synthase1 gene is required for myo-inositol synthesis and suppression of cell death. Plant Cell 2010, 22, 888–903.
- Chen, H.; Xiong, L. myo-inositol-1-phosphate synthase is required for polar auxin transport and organ development. J. Biol. Chem. 2010, 285, 24238–24247.
- Sato, Y.; Yazawa, K.; Yoshida, S.; Tamaoki, M.; Nakajima, N.; Iwai, H.; Ishii, T.; Satoh, S. Expression and functions of myo-inositol monophosphatase family genes in seed development of Arabidopsis. J. Plant Res. 2011, 124, 385–394.
- Nourbakhsh, A.; Collakova, E.; Gillaspy, G.E. Characterization of the inositol monophosphatase gene family in Arabidopsis. Front. Plant Sci. 2014, 5, 725.
- Sweetman, D.; Stavridou, I.; Johnson, S.; Green, P.; Caddick, S.; Brearley, C. Arabidopsis thaliana inositol 1,3,4-trisphosphate 5/6-kinase 4 (AtITPK4) is an outlier to a family of ATP-grasp fold proteins from Arabidopsis. FEBS Lett. 2007, 581, 4165–4171.
- Xia, H.; Brearley, C.; Elge, S.; Kaplan, B.; Fromm, H.; Mueller-Roeber, B. Arabidopsis inositol polyphosphate 6-/3-kinase is a nuclear protein that complements a yeast mutant lacking a functional ArgR-Mcm1 transcription complex. Plant Cell 2003, 15, 449–463.
- Zhang, Z.B.; Yang, G.; Arana, F.; Chen, Z.; Li, Y.; Xia, H.J. Arabidopsis inositol polyphosphate 6-/3-kinase (AtIpk2beta) is involved in axillary shoot branching via auxin signaling. Plant Physiol. 2007, 144, 942–951.
- Munnik, T.; Vermeer, J. Osmotic stress-induced phosphoinositide and inositol phosphate signalling in plants. Plant Cell Environ. 2010, 33, 655–669.
- Zhao, H.; Frank, T.; Tan, Y.; Zhou, C.; Jabnoune, M.; Arpat, A.B.; Cui, H.; Huang, J.; He, Z.; Poirier, Y.; et al. Disruption of OsSULTR3;3 reduces phytate and phosphorus concentrations and alters the metabolite profile in rice grains. New Phytol. 2016, 211, 926–939.
- Shoemaker, R.C.; Polzin, K.; Labate, J.; Specht, J.; Brummer, E.C.; Olson, T.; Young, N.; Concibido, V.; Wilcox, J.; Tamulonis, J.P.; et al. Genome duplication in soybean (Glycine subgenus soja). Genetics 1996, 144, 329–338.
- Yamaji, N.; Ma, J.F. Node-controlled allocation of mineral elements in Poaceae. Curr. Opin. Plant Biol. 2017, 39, 18–24.
- Liu, F.; Chang, X.; Ye, Y.; Xie, W.; Wu, P.; Lian, X. Comprehensive sequence and whole-life-cycle expression profile analysis of the phosphate transporter gene family in rice. Mol. Plant 2011, 4, 1105–1122.
More
Information
Subjects:
Plant Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
901
Revision:
1 time
(View History)
Update Date:
08 Oct 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No