Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Chang Pyo Hong | + 2441 word(s) | 2441 | 2021-08-18 04:35:13 |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Hong, C.P. Gibberellin. Encyclopedia. Available online: https://encyclopedia.pub/entry/13591 (accessed on 26 June 2026).
Hong CP. Gibberellin. Encyclopedia. Available at: https://encyclopedia.pub/entry/13591. Accessed June 26, 2026.
Hong, Chang Pyo. "Gibberellin" Encyclopedia, https://encyclopedia.pub/entry/13591 (accessed June 26, 2026).
Hong, C.P. (2021, August 26). Gibberellin. In Encyclopedia. https://encyclopedia.pub/entry/13591
Hong, Chang Pyo. "Gibberellin." Encyclopedia. Web. 26 August, 2021.
Copy Citation
Gibberellins (GAs) are an important group of phytohormones associated with diverse growth and developmental processes, including cell elongation, seed germination, and secondary growth.
gibberellins
Panax ginseng
GID1s
phytohormones
storage root secondary growth
1. Introduction
Korean ginseng (Panax ginseng C.A. Meyer) has been used as an important medicinal plant species for thousands of years in Asia, especially in Korea, Japan, and China [1]. Ginseng has the ability to increase immunity and vitality and to prevent aging, and its pharmacological effects have been proven through clinical trials and animal experiments [2]. Studies on ginseng have focused primarily on its pharmacological effects in humans; however, its unique physiological and developmental characteristics have rarely been explored thoroughly due to its perennial heterozygous nature [3]. The application of functional genomic approaches in ginseng has been challenging because of its unique physiological and ecological characteristics and the lack of genomic information [4,5], which could be attributed to its relatively large genome (3.2 Gbp), heterozygous allotetraploid nature (2n = 4x = 48), and significant amounts of repetitive DNA [6,7].
Plants, as sessile organisms, induce a variety of physiological changes in response to adverse environmental conditions. Plant hormones function as master regulators of growth and development under unsuitable external environments [8]. Gibberellins (GAs) are tetracyclic diterpenoid hormones that play essential roles in seed germination, flowering, pollen maturation, stress tolerance, secondary growth, and development [9,10,11]. Recent studies have shown that there are complicated internal and external signaling interactions of GAs and various hormone pathways, including auxin, abscisic acid (ABA), jasmonic acid (JA), ethylene, and cytokinin. These interactions are highly relevant to plant growth, development, and stress tolerance [12,13,14]. Major GA signaling components have been identified through genetic studies on model plant species, including rice (Oryza sativa) and Arabidopsis thaliana. The nucleocytoplasmic GA receptor gibberellin-insensitive dwarf1 (GID1) was first identified in rice through studies on gibberellin-insensitive dwarf mutants [15]. OsGID1 and AtGID1s have been characterized as water-soluble GA receptors that exhibit strong binding affinity for bio-active GAs [15,16]. At the C-terminus, GID1 harbors a well-conserved α/β-hydrolase domain, which belongs to the carboxylesterase family of plant proteins [17,18]. The canonical GA signaling pathway is initiated by direct binding of active GAs to GID1 in the nucleus. The GA–GID1 complex induces rapid degradation of the major growth-inhibitory factor DELLA through direct protein–protein interactions [12,19]. DELLAs are key transcriptional repressors that inhibit GA signaling and plant growth. Degradation of DELLA proteins requires both GID1 and the F-box protein SLY1 (SLEEPY)/GID2 [15,20,21,22]. In the presence of GA, DELLA is recognized by the SCFSLY1/GID2 ubiquitin E3 ligase complex for subsequent 26S proteasome-meditated degradation [21,23,24]. This canonical GA signaling pathway is responsible for various physiological responses during plant growth and development.
The secondary growth of roots is particularly important for production of various root crops and for accumulation of useful compounds and nutrients in storage roots [25,26,27]. Studies on root crops such as radish and cassava show that the secondary growth of storage roots is regulated by the activity of cambium stem cells. Secondary growth of plants involves formation of cambium stem cells and differentiation of phloem and xylem tissues, which are mainly regulated by the plant hormones auxin and cytokinin [25,26,27,28,29]. In addition, the role of GA in secondary xylem formation and lignification in storage roots has also been confirmed in several plant species, including poplar, carrot, and cotton [30,31,32,33]. Overproduction of GA promotes elongation and division of xylem and fiber cells in the vascular bundle and increases cambium activity [26,32,34]. However, in carrot and sweet potato, exogenous GA treatment inhibits root growth by affecting cell division and vascular lignin synthesis [35,36]. These results suggest that the physiological response to GA, a growth-promoting hormone, varies with the plant species. Although Korean ginseng is one of the most important medicinal root crops with a 6-year cultivation period, genetic and physiological factors affecting the growth and development of ginseng roots have rarely been investigated.
Owing to recent advances in sequencing technology, the draft genome sequence of P. ginseng has been released by two research groups [37,38]. In addition, we also previously reported the precise transcript sequences of P. ginseng using PacBio single-molecule real-time (SMRT) isoform sequencing (Iso-seq) analysis [39].
2. GA Enhances Shoot Primary Growth and Root Secondary Growth in P. ginseng
The secondary growth of the tap root, as a storage organ, is one of the main factors affecting ginseng yield. To investigate the physiological effects of GA in P. ginseng, active GA3 and paclobutrazol (PCZ), a GA biosynthetic inhibitor, were applied exogenously to the roots of 1-year-old P. ginseng plants. Exogenous application of GA significantly promoted stem growth compared with mock treatment (Figure 1A,B), which is consistent with the role of GA as a growth-promoting hormone [40]. Interestingly, exogenous GA treatment also increased the diameter of the tap root; however, PCZ application reduced both shoot and root growth in P. ginseng (Figure 1A,B). To further analyze the GA-mediated promotion of shoot and root growth, GA- and PCZ-treated P. ginseng samples were applied to histological paraffin-embedded sections with Safranine-Astra blue combination staining. Compared with the control, plants in the GA treatment group showed longer epidermal cells, consistent with their elongated stem phenotype (Figure 1C). Conversely, the stems of plants in the PCZ treatment group showed slightly shorter cells compared with the control (Figure 1C). However, there was no significant difference in the size of the divided cells surrounding the cambium layer of the storage tap root, although the number of divided starch-deposited storage parenchyma cells located between xylem vessels and resin duct cells was greatly increased in GA-treated P. ginseng roots (Figure 1D). Consistently, the number of cambium-derived storage parenchyma and vascular cells in PCZ-treated roots was significantly reduced (Figure 1D). These results indicate that GA facilitates root secondary growth in P. ginseng by promoting storage parenchyma cell development.

Figure 1. Exogenous gibberellin (GA) treatment promotes primary growth of stems and secondary growth of roots in Panax ginseng. (A) Phenotype of 1-year-old P. ginseng plants treated with DMSO (control [Con]), 10 μM GA3, and 100 μM paclobutrazol (PCZ) once a week for 8 weeks. Scale bar = 2 cm. (B) Measurements of shoot length and root diameter. (C) Representative images of stained stem cross-sections of P. ginseng plants treated with DMSO (Con), GA3 and PCZ. Scale bar = 100 μm. (D) Quantification of cell length in the indicated treatments. (E) Representative images of stained root cross-sections of P. ginseng plants treated with DMSO (Con), GA3, and PCZ. XV: xylem vessel, CZ: cambial cell layer zone, RD: resin duct cells. Scale bar = 100 μm (F) Quantification of cambium-derived cells in the XV and RD of each ray. In (B,D,F), dots, squares and triangles represent individual values. Error bars represent standard error; n = 16 (B), 20 (D), 10 (F). Different lowercase letters indicate statistically significant differences (p < 0.05; one-way analysis of variance [ANOVA], followed by Tukey’s multiple range test).
3. Identification of Putative GA Receptor Genes in P. ginseng Genome
The growth-promoting effect of GA on the stem and root growth of P. ginseng suggests that the canonical GA signaling pathways are likely evolutionarily conserved in P. ginseng. Since GA signal transduction initiates with its perception, we searched for GA receptor-encoding genes in the P. ginseng genome by analyzing the two genome sequence drafts of P. ginseng and PacBio Iso-seq data [21,22,23]. A BLAST search of the AtGID1A amino acid sequence led to identification of eight putative GID1 DNA sequences in P. ginseng, designated as PgGID1A–H (Figure 2A). Phylogenetic analysis of PgGID1s and AtGID1s showed that five PgGID1s (A, B, C, E, and F) clustered with AtGID1A and AtGID1C, while the remaining three PgGID1s (D, G, and H) clustered with AtGID1B (Figure 2A). Next, we examined the secondary structure of OsGID1 [41] (Figure 2B) and compared the amino acid sequences of PgGID1s with those of AtGID1s and OsGID1 (Figure 2C). A topology diagram based on the predicted secondary structure of OsGID1 provided the structure information of PgGID1s (Figure 1B). The N-terminal region and two α-helices (α8 and α9; located between the β6 and β7 sheets) of OsGID1 corresponded to the left and right sides of the lid structure, respectively (Figure 2B). It is well characterized that OsGID1, one of the hormone-sensitive lipases (HSLs), contains an evolutionarily conserved HGG sequence and a catalytic triad (S, D, and H), both of which are essential for its enzymatic activity; H in the SDH catalytic triad is replaced by V. In PgGID1s, the V residue is replaced by I, as in AtGID1B and AtGID1C (Figure 2C). Furthermore, PgGID1s showed high sequence similarity with AtGID1s and OsGID1 (Figure 2C). Additionally, important amino acid residues and motifs (Figure 2B) were well-conserved among the PgGIDs, suggesting that PgGID1s function as GA receptors. It is well-known that GID1 proteins act as molecular glue to facilitate the interactions of DELLA repressors with SCFSLY1/GID2 in a GA-dependent manner [41]. To confirm subcellular localization of PgGID1s, we co-expressed the PgGID1-GFP fusions and AtARR2-RFP (as a nuclear marker) in Arabidopsis protoplasts. Similar to the nucleocytoplasmic localization of AtGID1s and OsGID1 [42], all four PgGIDs (A–D) localized to the nucleus and cytoplasm (Figure 2D).

Figure 2. Phylogenetic analysis and amino acid sequence alignment of GID1 proteins. (A) Phylogenetic analysis of PgGID1A–H, AtGID1s, and OsGID1. The phylogenetic tree was constructed using the MEGA7 program. Horizontal branch lengths are proportional to the estimated number of amino acid substitutions per residue. Bootstrap values were obtained by 1000 bootstrap replicates. Pg, Panax ginseng; At, Arabidopsis thaliana; Os, Oryza sativa. (B) Topology diagram based on the predicted secondary structure of the OsGID1 protein [15]. (Blue circles indicate important residues involved in the GID1–SLR1 interaction, and red dots indicate important residues involved in OsGID1–GA and GID1–SLR1 interactions. Red stars indicate residues essential for enzymatic activity. Colored zones indicate the lid (yellow) and binding pocket (red). (C) Amino acid sequence alignment of the GID1 proteins of Arabidopsis, rice, and P. ginseng constructed using SMS (https://www.Bioinformatics.org (accessed on 1 May 2021)). (D) Subcellular localization analysis of PgGID1A–D proteins in Arabidopsis protoplasts. Full-length coding sequences of PgGID1A–D were fused to the GFP reporter gene. The nucleus was visualized using the AtARR2-RFP nuclear marker. GFP and RFP fluorescence images were merged. Scale bar = 50 μm.
4. Complementation Analysis of the Atgid1a/c Doble Mutant
To evaluate the biological roles of PgGID1s in the GA signaling pathway, we performed complementation analysis of the Arabidopsis Atgid1a/c double knockout mutant, which exhibits a GA-deficient semidwarf phenotype [20]. PgGID1A–D were individually overexpressed in the atgid1a/c double mutant under the control of the constitutively active 35S promoter (Figure 3A–C). All complementation lines expressing PgGID1s showed a wild-type shoot phenotype (Figure 3A) and partially enhanced silique length compared with Atgid1a/c plants (Figure 3B). These results indicate that PgGID1s can replace the function of AtGID1s during plant growth and development. Next, we investigated whether exogenous GA3 enhances the interaction of PgGIDs with PgDELLA proteins (Figure 3D). We cloned five DELLA-encoding genes of P. ginseng (named as PgRGA1–5), and ca) (Supplementary Table S2), contributing to the essential role for secondary cell wall biosynthesis. We carried out yeast two-hybrid assays in the presence or absence of exogenous GA3. In the absence of active GA3, rare physical protein interactions between all PgGID1s and PgRGA1, 2 and 3 proteins occurred, but the protein interactions between PgGIDs and PgRGAs (except between PgGID1D and PgRGA1-3) were enhanced in the presence of GA3. However, PgGID1D interacted only with PgRGA4 and PgRGA5 in a GA3-independent manner (Figure 3D). These results reveal interaction specificity between PgGIDs and PgDELLAs.
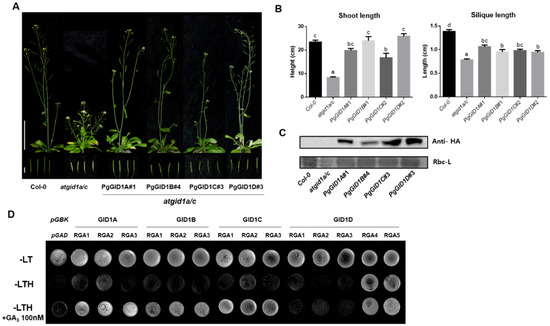
Figure 3. Complementation of the Arabidopsis gid1 loss-of-function mutant via PgGID1 overexpression. (A) Rescue of the GA-insensitive dwarf phenotype of the atgid1a/c double mutant by overexpression of PgGID1A–D genes. Scale bar = 5 cm (top panel) and 0.5 cm (bottom panel). (B) Measurement of the length of shoots and siliques shown in (A). Error bars represent the standard error (n > 12). Different lowercase letters indicate statistically significant differences (p < 0.05; one-way ANOVA, followed by Tukey’s multiple range test). (C) Western blot analysis of PgGID1A–D protein levels in atgid1a/c plants. (D) Yeast two-hybrid assay. To test the interaction between PgGID1s and PgRGAs, PgGID1A–D-carrying pGBKT7 constructs (bait) were co-expressed with PgRGA1–5-harboring pGADT7 constructs (prey) in yeast cells, which were grown on media (-LTH) supplemented with or without 100 nM GA3.
5. Transcriptome Analysis of Root Secondary Growth in Response to GA in P. ginseng
To investigate the root secondary growth of P. ginseng in response to GA, we analyzed a total of 5721 genes differentially expressed between DMSO- and GA-treated roots using RNA-seq data (Supplementary Figure S1). Gene ontology (GO) enrichment analysis of differentially expressed genes (DEGs) revealed significant enrichment of the following functional categories: ‘cell cycle and/or division’ (p-values of sub-GO terms in the representative category: p = 4.1 × 10−3 − 1.6 × 10−4), ‘developmental process’ (p = 4.7 × 10−3 − 3.1 × 10−8), ‘cell growth’ (p = 1.7 × 10−3) and ‘cell wall biogenesis’ (p = 9.2 × 10−3 − 7.0 × 10−6) (Figure 4A). This suggests that GA regulates root secondary growth by controlling root elongation via cell division. Interestingly, the functional enrichment of ‘response to nitrogen’ (p = 3.8 × 10−3 − 8.6 × 10−8), related to nitrate transport and assimilation, was also identified (Figure 4A). Expression of nitrate transporter genes (NRTs) was increased, whereas that of nitrate reductase genes (NIRs) was decreased, suggesting that GA signaling influences the nitrate metabolic process through the transcriptional regulation of NRTs [43]. The results of gene set enrichment analysis (GSEA) further supported the antagonistic transcriptional responses of ABA-related signaling pathway in GA-treated ginseng roots (Figure 4B). Interestingly, JA-responsive genes were significantly enriched among the up-regulated genes in GA-treated P. ginseng roots (Figure 4B). These results suggest that JA-mediated downstream signaling pathways are also closely associated with the GA-mediated activation of cambium stem cells.
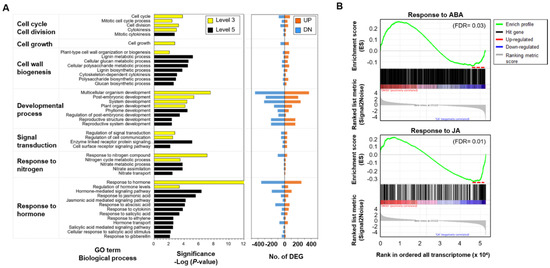
Figure 4. Transcriptome profiling of P. ginseng roots treated with or without GA. (A) Gene ontology (GO) enrichment analysis of differentially expressed genes (DEGs) identified by comparison of GA- and DMSO-treated root samples. GO terms of level 3 (yellow bars) and level 5 (black bars), with EASE score < 0.01, were selected (left panel). The number of up-regulated genes (red) and down-regulated genes (blue) categorized under the enriched GO terms are shown in the right panel. (B) Enrichment plot for the responses to ABA (GO: 0009737) and JA (GO: 0009753) in the transcriptome data of DMSO- and GA-treated root samples. In the enrichment plot, the red dotted line represents the gene subset that made the largest contribution to the enrichment score (ES) (false discovery rate [FDR] < 0.05). The ranking list metric in the plot measures the correlation between a gene and the plant phenotype. In the ranking list, positive values indicate genes up-regulated in DMSO-treated root samples with red color gradient, and negative values indicate genes down-regulated in DMSO-treated root samples.
Information
Subjects:
Plant Sciences
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revision:
1 time
(View History)
Update Date:
26 Aug 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No