 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ainhoa Alberro | + 2667 word(s) | 2667 | 2021-08-23 11:06:35 | | | |
| 2 | Catherine Yang | -10 word(s) | 2657 | 2021-08-26 04:15:19 | | |
Video Upload Options
Extracellular vesicles (EVs) are important players for intercellular communication. EVs are secreted by almost all cell types; they can transfer information between nearby or distant cells, and they are highly abundant in body fluids. The present work provides an overview of the components, effects, and applications of EVs in blood.
1. Introduction
Extracellular vesicles (EVs) are membrane-coated particles secreted by almost all cell types. Their first identification was already reported in 1946 as procoagulant platelet-derived particles in normal plasma [1], and more than 20 years later, in 1967, they were referred as “platelet-dust” [2]. Since then, several publications started to report novel particle sources and functions and, by the end of the 20th century, they were already known to play a role in relevant processes, such as antigen presentation [3]. Importantly, at the beginning of the present century, research on EVs gained interest among the scientific community, as they were implicated in other central issues, such as the immune system mediated antitumour response [4], and also due to the discovery that EVs transfer mRNAs and microRNAs (miRNAs) from the donor to recipient cells inducing functional changes [5]. In the last two decades, thousands of works have continued to describe the characteristics, functions, and implications of EVs and their cargo in intercellular communication. Thanks to all of them, we can now state that EVs are important players in most biological processes.
EVs are present in diverse tissues and biofluids, and one of the most studied sources is blood. Circulating EVs are relatively easy to obtain with minimally invasive samplings, and more importantly, they have been found to be essential mediators of cell communication between different tissues and to be implicated in diverse cellular processes [6][7][8][9][10]. EVs originated from blood cells and from many other tissues compose the complex pool of EVs circulating in blood, and similarly, they can reach most of the tissues of the organism. Furthermore, due to the possibility of directing EVs in circulation to other tissues, the potential use of EV-based treatments through administration in blood has also been proposed and investigated. The aim of this review is to gather and present the studies of EVs that have been carried out in EVs isolated from blood or with EVs introduced in circulation, and to gain the whole perspective of EVs in circulation and their potential uses.
2. Biological Characteristics of Extracellular Vesicles
The term EVs is used to refer to all the particles that cells secrete to the extracellular media. There are two main categories of EVs: exosomes and microvesicles. Besides, apoptotic bodies are also considered EVs. Indeed, apoptotic bodies play an essential role in the proper clearance of the dying cell, as well as for the signalling of this programmed cell death to surrounding cells and for the regeneration of the tissue [11]. However, most of the works studying EVs are focused on exosomes and microvesicles, due to their multiple functions and implications. Exosomes are secreted particles originated from the fusion of a multivesicular body and the plasma membrane, while microvesicles are formed by the direct budding and fission of the plasma membrane.
With regard to the molecules carried by EVs, we have to consider both their membrane and inner cargo. The membrane of EVs consists mainly of proteins and lipids, but each EV has distinct types of proteins and lipids depending on their origin and function. Furthermore, the composition of the EV membrane influences the fate and internalization by recipient cells [12]. The components of the EV lumen are even more diverse and include proteins and many different nucleic acids. Apart from the above-mentioned mRNA and miRNAs, EVs carry other types of small and long non-coding RNAs, circular RNAs, and double-stranded DNA fragments [13][14][15]. Importantly, investigations about EV secretion and their cargo have revealed that the sorting of components into a forming particle is a controlled mechanism and not a random packaging of the available molecules in the secreting cell [5][16].
Similarly, the uptake of EVs is a controlled process. Many authors have studied the binding and internalization of EVs by recipient cells and multiple molecules, such as tetraspanins, integrins, lipids, and lectins, which have been identified to mediate the uptake [17]. The integration of EVs by recipient cells can be performed by the fusion of the EV and cellular membranes, or by distinct endocytic pathways. An extensive and complete review on the biogenesis, release, and targeting mechanisms of EVs was recently published, and it is a recommended read to go into this subject in depth [12].
3. Extracellular Vesicles Circulating in Blood
EVs secreted by blood cells, but also EVs originated from many other tissues, can be found in the circulatory system. As a consequence, the studies performed in EVs isolated from blood samples have shown the complex pool of particles present in this fluid (Figure 1).
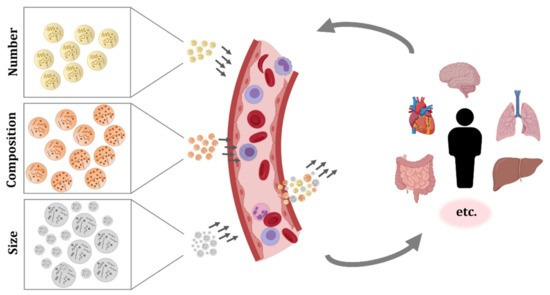
The most abundant EVs in blood are the ones from platelets. They were also the first ones to be described and were referred to as “platelet-dust” [1][2]. Investigations of EVs in plasma revealed the abundance of EVs from platelets as well as from erythrocytes, but also the influence of preanalytical factors such as blood sample storage time, temperature, and anticoagulants on their secretion [18][19][20].
Apart from platelets and erythrocytes, other blood cells also produce EVs. For instance, it has been demonstrated that cells such as mast cells, neutrophils, and eosinophils secrete EVs, but these reports are from in vitro experiments and they do not verify the presence of EVs from those cell types in circulation [21][22][23]. In the case of basophils, to our knowledge, there is no evidence of EV secretion. Under normal conditions, there are few basophils circulating in blood, and when activated, they secrete the contents of the high amount of granules they bear, releasing the inner content, but they do not produce EVs [24]. The production and secretion of EVs by other cell populations circulating in blood, including monocytes, macrophages, dendritic cells, NK cells, and B and T lymphocytes has been extensively reported, but similarly, most works were performed with cultured cells [3][25][26][27][28] and only few publications identified EVs in circulation with characteristic membrane molecules that could confirm the in vivo secretion of EVs from these immune cells [29][30][31][32]. Recently, a comprehensive work further evaluated the membrane molecules of EVs from plasma and serum, and pointed to the importance of the EV source and detection method on the identification of the markers [33].
One of the first works that identified EVs secreted by other cell types in blood was published more than 20 years ago. They detected vitronectin receptor (αvβ3) and other endothelial markers in human umbilical vein endothelial cells (HUVECs) and EVs secreted by these cells in vitro, and importantly, they also confirmed the presence of endothelial EVs in human plasma [34]. Similarly, EVs positive for adipocyte-specific markers such as adiponectin and resistin were first found in mice serum, and later, the presence of adipocyte-derived EVs in human plasma was confirmed by several adipocyte markers and adipokines [35][36][37][38]. The connection between the liver and the circulatory system suggested the presence of EVs from the liver in blood; this was confirmed in mice and human samples, in which hepatocyte-derived EVs were detected [39][40]. In addition, the muscle tissue also secretes EVs and, indeed, striated muscle-specific miRNAs or myo-miRNAs have been found in human plasma EVs [41][42]. Moreover, EVs produced by cardiomyocytes have been identified in mice and humans, as measured by cardiac bridging integrator 1 (cBIN1)-containing particles in circulation [43]. A systematic review on EVs secreted by six cardiac cell types, their cargo, and functions was published elsewhere, summarizing in vitro investigations and some studies performed in blood [44]. Another relevant communication system mediated by EVs was found in pregnant women, as indicated by the presence of placental-derived EVs (bearing placental-type alkaline phosphatase) in blood [45].
Notably, cancer cells from different origins also produce EVs, and these can enter the circulatory system [46][47][48]. Another type of EVs with great interest and potential uses was found in blood some years ago: EVs coming from the central nervous system (CNS). The ability of EVs to cross the blood–brain barrier (BBB) was confirmed and, thus, it is now known that particles produced by CNS cells can circulate in blood, and that EVs from diverse origins can enter the CNS [49][50][51].
Moreover, EVs are secreted in physiological and pathological conditions, and depending on their cargo and on the conditions of receptor cells, they can have beneficial or detrimental effects [52].
4. Blood Extracellular Vesicles in Physiological and Pathological Processes
As introduced before, it is long known that EVs are functional particles. More than 60 years ago, EVs were described to play a role in blood coagulation [1][2]. Since then, EVs have been found to be secreted by almost all cell types and to play a role in diverse physiological processes.
For instance, one of the first studies that characterized the miRNA signature of circulating EVs in healthy donors found the angiogenesis-related miRNA-126 enriched in EVs [53]. In addition, circulating EVs have been found to influence immune response. A work carried out with human samples also found MCH-II, FasL, and other immune markers in plasma EVs, and besides, they reported an effect of circulating EVs on CD4 T cell response [54]. Another study showed that plasma EVs influence monocyte and B cell activation [29], and a work carried out in our group showed their influence on CD4 and CD8 T cell activation [55]. Therefore, it becomes evident that blood EVs have a role in immune system responses, but we are still far from being able to understand the complex mixture of circulating particles and their effects in the face of the numerous stimuli and insults that can occur to a single organism throughout time.
Regarding other physiological processes, EVs have been shown to be implicated in feto-maternal communication [45][56][57][58][59][60], in aging [55][61][62][63][64] and also to be affected by physical activity [65][66][67][68][69]. Similarly, EVs have been studied in many pathological processes, including neurological [70][71][72][73][74][75][76] and inflammatory diseases [77][78], vascular pathologies [79][80][81], infectious diseases [82][83][84][85], preganancy complications [86][87][88] or cancer [89][90][91][92][93][94][95] (Figure 2). Aiming to contribute to disease diagnosis, monitoring, and treatment, many researchers have investigated circulating EVs. Certainly, blood is a relatively easily accessible biofluid and can reflect modifications from different tissues and organs. However, much work is still required to validate the proposed candidates.
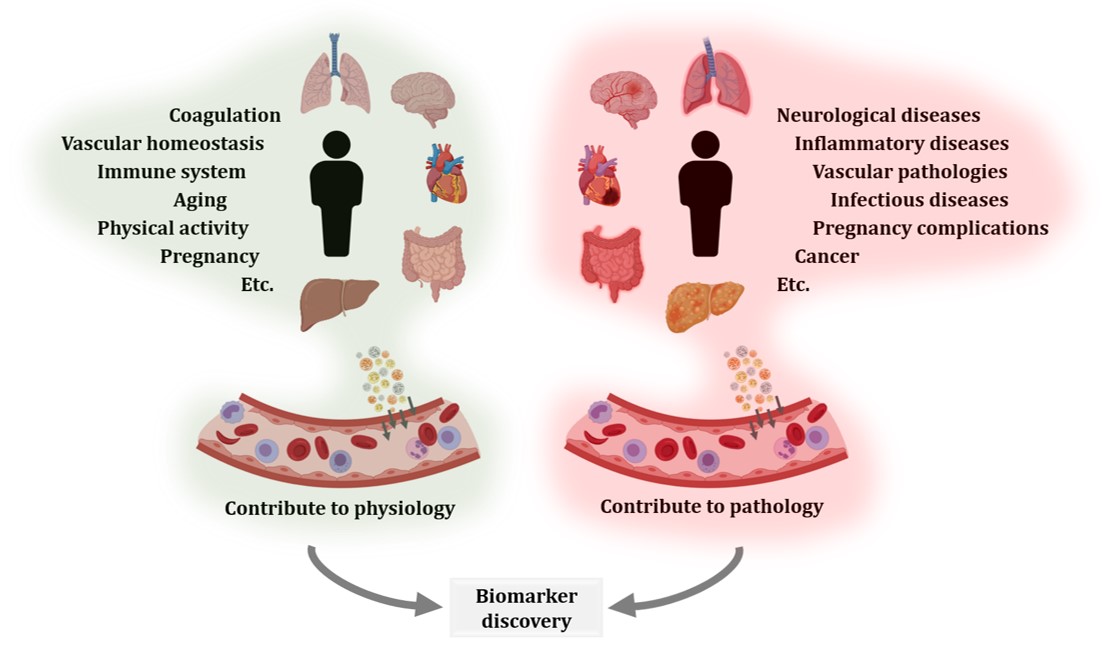
Figure 2. Extracellular vesicles (EVs) contribute to physiological and pathological processes. EVs secreted by different organs, tissues, and cell types reach systemic circulation. EV production and circulation occurs both in physiological and pathological states. Therefore, comparison of blood EVs from healthy and diseased donors, including different disease stages or treatments, is an interesting approach for the characterization of biological processes and biomarker discovery.
5. Potential Applications of Extracellular Vesicles Introduced in Circulation
The use of cell therapies, particularly the ones based on stem cells, was proposed to have great potential. Additionally, in recent years, the possibility of administering EVs have gained interest, as they can have the same beneficial effects as progenitor cells while reducing their risks, such as uncontrolled proliferation or transplant rejection [96]. Furthermore, the use of EVs has additional benefits, including the easier storage and distribution, as well as the numerous possible routes of administration and modes of application [97]. Another advantage of EVs is that they are formed from cells and thus, biologically designed to be taken up by recipient cells. In contrast, other constructs, such as liposomes or nanoparticles, are easily loaded with the molecule or drug of interest, but they could face biodistribution or targeting problems, reducing their efficacy [98]. Moreover, there are several techniques for obtaining EVs enriched in a particular compound. On the one side, we can modify the EV-producing cell by transfection or transduction by culturing them under a particular stress or condition, or by incubating them with the molecule of interest. On the other side, EVs can be modified after their production, by electroporation, sonication, extrusion, or other methods [98][99][100] (Figure 3).
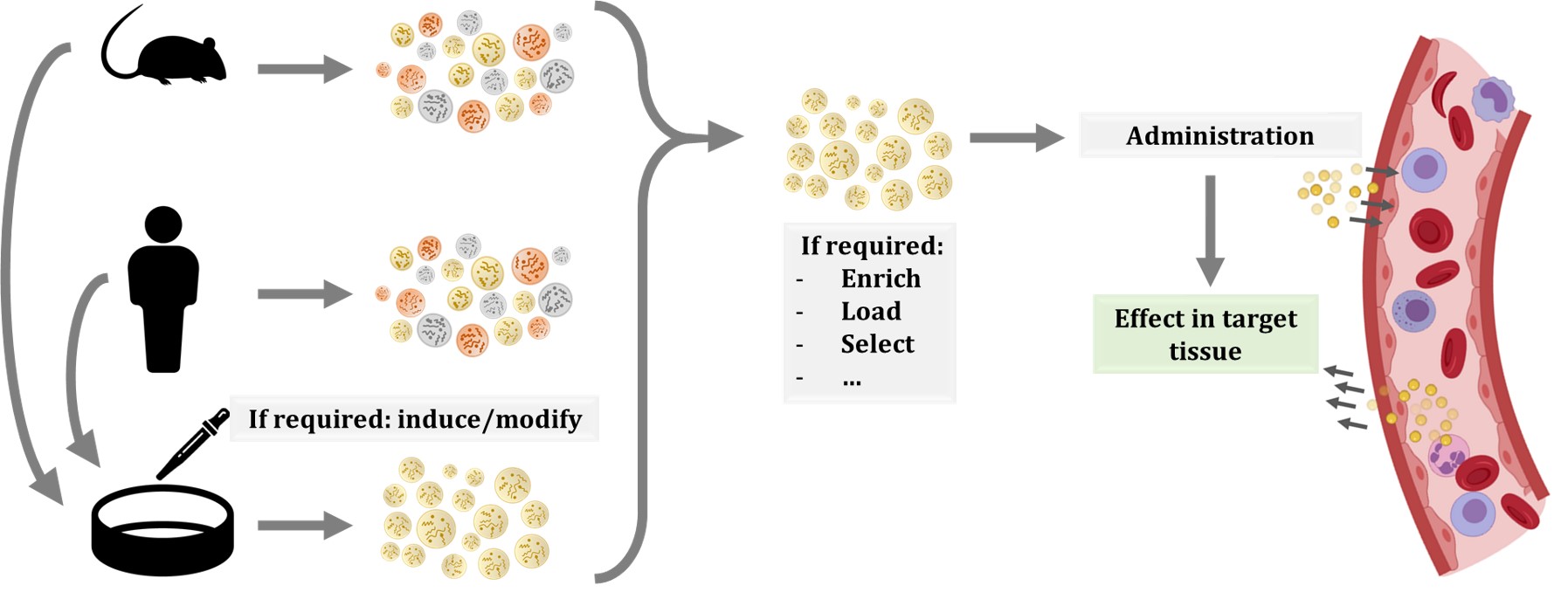
Figure 3. Schematic representation of possible workflows for extracellular vesicle (EV) administration. EVs or producing cells can be isolated from human donors or other organisms. In the case of cells, specific culture protocols are applied to induce the production of EVs with the desired features or cargo. Therefore, EVs produced in culture are more homogeneous than EVs isolated from whole organisms. In any case, once EVs are isolated, other procedures can be applied to EVs, including enrichment, or loading of molecules of interest. Then, EVs are injected in circulation and, thanks to their membrane composition and cargo, they will migrate to the target tissue and exert their effect.
Many studies have been performed with EVs introduced in circulation, most of them in mice. Some investigations focused on the effect of EVs from patients with diverse pathologies, while others evaluated the impact of murine EVs. There are also some works that evaluated parasite-derived EVs [101][102][103]. A pioneering work in the field was published already 10 years ago, with dendritic cell-derived EVs. Primary cells from murine bone marrow were cultured and transfected to produce EVs that bear the CNS-targeting rabies viral glycoprotein (RVG) peptide at their membrane. Then, EVs were isolated and electroporated with the siRNA of interest. The intravenous injection of these EVs resulted in brain targeting and importantly, downregulation of BACE1—a protease implicated in amyloid aggregation in Alzheimer’s disease—was achieved [104]. Besides, this system was effectively applied to target Parkinson’s disease related to α-synuclein [105]. The effect of systemically administered EVs in the CNS has also been demonstrated in acute problems, for instance in traumatic brain injury. A single dose of miRNA-124-enriched EVs 24 h after traumatic brain injury in rats resulted in a significant improvement [106].
Moreover, the effect of systemic injection of EVs has been reported to also affect metabolic processes, such as insulin sensitivity [107][108]. The potential applications of EVs are also being investigated with regard to other pathologies and complications. In the case of transplantations, EVs enhanced allograft tolerance and long-term survival in animal models [109][110] and experiments performed with human EVs obtained promising results in a wound-healing mouse model [111]. In addition, a recent study points to the theranostic potential of autologous circulating EVs. The authors demonstrated that fluorescently labelled EVs from colorectal cancer patients, when injected in mice, selectively target patient-derived xenografts. Furthermore, their preliminary experiments performed in mast cells and mammary tumour-bearing dogs also showed tumour tropism of autologous plasma EVs that were fluorescently labelled [112].
These and other studies demonstrate the promising applicability of EVs in blood, but they also highlight the multiple approaches that are being tested and the need for standardization. Before routine clinical application of EVs could be reached, aspects such as the cell of origin, EV production and isolation methods, possible EV loading or modifications, storage, administration, and dosing should be further investigated [113][114][115].
Despite current knowledge limitations, there are already hundreds of clinical trials registered (clinicaltrials.gov) involving EVs, also termed exosomes or microparticles/microvesicles. It should be noted that most of them focus on the analysis of EVs from body fluids as biomarkers for disease diagnosis or monitoring. However, there are also clinical trials evaluating EV administration, and a high proportion of them test EVs introduced in circulation [116][117]. To date, there are few finished clinical trials that reported their results, but the available information indicates that EV administration is safe and well tolerated by patients with diverse pathologies. However, the efficacy of tested EV therapies is moderated. Results from ongoing and future clinical trials are needed to be able to critically evaluate the impact of EV-based treatments.
References
- Chargaff, E.; West, R. The biological significance of the thromboplastic protein of blood. J. Biol. Chem. 1946, 166, 189–197.
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 13, 269–288.
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B Lymphocytes Secrete Antigen-presenting Vesicles. J. Exp. Med. 1996, 183, 1161–1172.
- Wolfers, J.; Lozier, A.; Raposo, G.; Regnault, A.; Théry, C.; Masurier, C.; Flament, C.; Pouzieux, S.; Faure, F.; Tursz, T.; et al. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat. Med. 2001, 7, 297–303.
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659.
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208.
- Urbanelli, L.; Buratta, S.; Sagini, K.; Tancini, B.; Emiliani, C. Extracellular vesicles as new players in cellular senescence. Int. J. Mol. Sci. 2016, 17, 1408.
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317.
- Dickhout, A.; Koenen, R.R. Extracellular Vesicles as Biomarkers in Cardiovascular Disease; Chances and Risks. Front. Cardiovasc. Med. 2018, 5, 113.
- Kikuchi, S.; Yoshioka, Y.; Prieto-Vila, M.; Ochiya, T. Involvement of extracellular vesicles in vascular-related functions in cancer progression and metastasis. Int. J. Mol. Sci. 2019, 20, 2584.
- Kalra, H.; Drummen, G.P.C.; Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. Int. J. Mol. Sci. 2016, 17, 170.
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228.
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769.
- Li, Y.; Zhao, J.; Yu, S.; Wang, Z.; He, X.; Su, Y.; Guo, T.; Sheng, H.; Chen, J.; Zheng, Q.; et al. Extracellular Vesicles Long RNA Sequencing Reveals Abundant mRNA, circRNA, and lncRNA in Human Blood as Potential Biomarkers for Cancer Diagnosis. Clin. Chem. 2019, 65, 798–808.
- Li, S.; Li, Y.; Chen, B.; Zhao, J.; Yu, S.; Tang, Y.; Zheng, Q.; Li, Y.; Wang, P.; He, X.; et al. ExoRBase: A database of circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids Res. 2018, 46, D106–D112.
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980.
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641.
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: Determination of their morphology, size, phenotype and concentration. J. Thromb. Haemost. 2014, 12, 614–627.
- Arraud, N.; Gounou, C.; Turpin, D.; Brisson, A.R. Fluorescence triggering: A general strategy for enumerating and phenotyping extracellular vesicles by flow cytometry. Cytom. Part. A 2016, 89, 184–195.
- Wisgrill, L.; Lamm, C.; Hartmann, J.; Preißing, F.; Dragosits, K.; Bee, A.; Hell, L.; Thaler, J.; Ay, C.; Pabinger, I.; et al. Peripheral blood microvesicles secretion is influenced by storage time, temperature, and anticoagulants. Cytom. Part. A 2016, 89, 663–672.
- Skokos, D.; Le Panse, S.; Villa, I.; Rousselle, J.-C.; Peronet, R.; David, B.; Namane, A.; Mécheri, S. Mast Cell-Dependent B and T Lymphocyte Activation Is Mediated by the Secretion of Immunologically Active Exosomes. J. Immunol. 2001, 166, 868–876.
- Hess, C.; Sadallah, S.; Hefti, A.; Landmann, R.; Schifferdi, J.A. Ectosomes released by human neutrophils are specialized functional units. J. Immunol. 1999, 163, 4564–4573.
- Mazzeo, C.; Cañas, J.A.; Zafra, M.P.; Marco, A.R.; Fernández-Nieto, M.; Sanz, V.; Mittelbrunn, M.; Izquierdo, M.; Baixaulli, F.; Sastre, J.; et al. Exosome secretion by eosinophils: A possible role in asthma pathogenesis. J. Allergy Clin. Immunol. 2015, 135, 1603–1613.
- Merluzzi, S.; Betto, E.; Ceccaroni, A.A.; Magris, R.; Giunta, M.; Mion, F. Mast cells, basophils and B cell connection network. Mol. Immunol. 2015, 63, 94–103.
- Esser, J.; Gehrmann, U.; D’Alexandri, F.L.; Hidalgo-Estévez, A.M.; Wheelock, C.E.; Scheynius, A.; Gabrielsson, S.; Radmark, O. Exosomes from human macrophages and dendritic cells contain enzymes for leukotriene biosynthesis and promote granulocyte migration. J. Allergy Clin. Immunol. 2010, 126, 1032–1040.
- Cypryk, W.; Öhman, T.; Eskelinen, E.L.; Matikainen, S.; Nyman, T.A. Quantitative proteomics of extracellular vesicles released from human monocyte-derived macrophages upon β-glucan stimulation. J. Proteome Res. 2014, 13, 2468–2477.
- Federici, C.; Shahaj, E.; Cecchetti, S.; Camerini, S.; Casella, M.; Iessi, E.; Camisaschi, C.; Paolino, G.; Calvieri, S.; Ferro, S.; et al. Natural-Killer-Derived Extracellular Vesicles: Immune Sensors and Interactors. Front. Immunol. 2020, 11, 262.
- Kormelink, T.G.; Mol, S.; de Jong, E.C.; Wauben, M.H.M. The role of extracellular vesicles when innate meets adaptive. Semin. Immunopathol. 2018, 40, 439–452.
- Ren, Y.; Yang, J.; Xie, R.; Gao, L.; Yang, Y.; Fan, H.; Qian, K. Exosomal-like vesicles with immune-modulatory features are present in human plasma and can induce CD4+ T-cell apoptosis in vitro. Transfusion 2011, 51, 1002–1011.
- Sáenz-Cuesta, M.; Irizar, H.; Castillo-Triviño, T.; Muñoz-Culla, M.; Osorio-Querejeta, I.; Prada, A.; Sepúlveda, L.; López-Mato, M.P.; López de Munain, A.; Comabella, M.; et al. Circulating microparticles reflect treatment effects and clinical status in multiple sclerosis. Biomark. Med. 2014, 8, 653–661.
- Zinger, A.; Latham, S.L.; Combes, V.; Byrne, S.; Barnett, M.H.; Hawke, S.; Grau, G.E. Plasma levels of endothelial and B-cell-derived microparticles are restored by fingolimod treatment in multiple sclerosis patients. Mult. Scler. 2016, 22, 1883–1887.
- Alberro, A.; Osorio-Querejeta, I.; Sepúlveda, L.; Fernández-Eulate, G.; Mateo-Abad, M.; Muñoz-Culla, M.; Carregal-Romero, S.; Matheu, A.; Vergara, I.; de Munain, A.L.; et al. T cells and immune functions of plasma extracellular vesicles are differentially modulated from adults to centenarians. Aging 2019, 11, 10723–10741.
- Wiklander, O.P.B.; Bostancioglu, R.B.; Welsh, J.A.; Zickler, A.M.; Murke, F.; Corso, G.; Felldin, U.; Hagey, D.W.; Evertsson, B.; Liang, X.M.; et al. Systematic methodological evaluation of a multiplex bead-based flow cytometry assay for detection of extracellular vesicle surface signatures. Front. Immunol. 2018, 9, 1326.
- Combes, V.; Simon, A.C.; Grau, G.E.; Arnoux, D.; Camoin, L.; Sabatier, F.; Mutin, M.; Sanmarco, M.; Sampol, J.; Dignat-George, F. In vitro generation of endothelial microparticles and possible prothrombotic activity in patients with lupus anticoagulant. J. Clin. Invest. 1999, 104, 93–102.
- Ogawa, R.; Tanaka, C.; Sato, M.; Nagasaki, H.; Sugimura, K.; Okumura, K.; Nakagawa, Y.; Aoki, N. Adipocyte-derived microvesicles contain RNA that is transported into macrophages and might be secreted into blood circulation. Biochem. Biophys. Res. Commun. 2010, 398, 723–729.
- Phoonsawat, W.; Aoki-Yoshida, A.; Tsuruta, T.; Sonoyama, K. Adiponectin is partially associated with exosomes in mouse serum. Biochem. Biophys. Res. Commun. 2014, 448, 261–266.
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253.
- Connolly, K.D.; Wadey, R.M.; Mathew, D.; Johnson, E.; Rees, D.A.; James, P.E. Evidence for adipocyte-derived extracellular vesicles in the human circulation. Endocrinology 2018, 159, 3259–3267.
- Povero, D.; Eguchi, A.; Li, H.; Johnson, C.D.; Papouchado, B.G.; Wree, A.; Messer, K.; Feldstein, A.E. Circulating extracellular vesicles with specific proteome and liver microRNAs are potential biomarkers for liver injury in experimental fatty liver disease. PLoS ONE 2014, 9, e113651.
- Garcia-Martinez, I.; Santoro, N.; Chen, Y.; Hoque, R.; Ouyang, X.; Caprio, S.; Shlomchik, M.; Coffman, R.L.; Candia, A.; Mehal, W.Z. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR9. J. Clin. Invest. 2016, 126, 859–864.
- Guescini, M.; Canonico, B.; Lucertini, F.; Maggio, S.; Annibalini, G.; Barbieri, E.; Luchetti, F.; Papa, S.; Stocchi, V. Muscle releases alpha-sarcoglycan positive extracellular vesicles carrying miRNAs in the bloodstream. PLoS ONE 2015, 10, e0125094.
- Carpi, S.; Polini, B.; Nieri, D.; Dubbini, N.; Celi, A.; Nieri, P.; Neri, T. Expression analysis of muscle-specific miRNAs in plasma-derived extracellular vesicles from patients with chronic obstructive pulmonary disease. Diagnostics 2020, 10, 502.
- Xu, B.; Fu, Y.; Liu, Y.; Agvanian, S.; Wirka, R.C.; Baum, R.; Zhou, K.; Shaw, R.M.; Hong, T.T. The ESCRT-III pathway facilitates cardiomyocyte release of cBIN1-containing microparticles. PLoS Biol. 2017, 15, e2002354.
- Xu, M.Y.; Ye, Z.S.; Song, X.T.; Huang, R.C. Differences in the cargos and functions of exosomes derived from six cardiac cell types: A systematic review. Stem Cell Res. Ther. 2019, 10, 194.
- Sabapatha, A.; Gercel-taylor, C.; Taylor, D.D. Specific isolation of placenta-derived exosomes from the circulation of pregnant women and their immunoregulatory consequences. Am. J. Reprod. Immunol. 2006, 56, 345–355.
- Vagner, T.; Spinelli, C.; Minciacchi, V.R.; Balaj, L.; Zandian, M.; Conley, A.; Zijlstra, A.; Freeman, M.R.; Demichelis, F.; De, S.; et al. Large extracellular vesicles carry most of the tumour DNA circulating in prostate cancer patient plasma. J. Extracell. Vesicles 2018, 7, 1505403.
- Peinado, H.; Alečković, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; García-Santos, G.; Ghajar, C.M.; et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891.
- Li, J.; Chen, J.; Wang, S.; Li, P.; Zheng, C.; Zhou, X.; Tao, Y.; Chen, X.; Sun, L.; Wang, A.; et al. Blockage of transferred exosome-shuttled miR-494 inhibits melanoma growth and metastasis. J. Cell. Physiol. 2019, 234, 15763–15774.
- Shi, M.; Sheng, L.; Stewart, T.; Zabetian, C.P.; Zhang, J. New windows into the brain: Central nervous system-derived extracellular vesicles in blood. Prog. Neurobiol. 2019, 175, 96–106.
- Shi, M.; Liu, C.; Cook, T.J.; Bullock, K.M.; Zhao, Y.; Ginghina, C.; Li, Y.; Aro, P.; Dator, R.; He, C.; et al. Plasma exosomal α-synuclein is likely CNS-derived and increased in Parkinson’s disease. Acta Neuropathol. 2014, 128, 639–650.
- Fiandaca, M.S.; Kapogiannis, D.; Mapstone, M.; Boxer, A.; Eitan, E.; Schwartz, J.B.; Abner, E.L.; Petersen, R.C.; Federoff, H.J.; Miller, B.L.; et al. Identification of pre-clinical Alzheimer’s disease by a profile of pathogenic proteins in neurally-derived blood exosomes: A case-control study. Alzheimer’s Dement. 2015, 11, 600–607.
- Goetzl, E.J.; Mustapic, M.; Kapogiannis, D.; Eitan, E.; Lobach, I.V.; Goetzl, L.; Schwartz, J.B.; Miller, B.L. Cargo proteins of plasma astrocyte-derived exosomes in Alzheimer’s disease. FASEB J. 2016, 30, 3853–3859.
- Lo Cicero, A.; Stahl, P.D.; Raposo, G. Extracellular vesicles shuffling intercellular messages: For good or for bad. Curr. Opin. Cell Biol. 2015, 35, 69–77.
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.L.T.; Schmittgen, T.D.; et al. Detection of microRNA expression in human peripheral blood microvesicles. PLoS ONE 2008, 3, e3694.
- Eitan, E.; Green, J.; Bodogai, M.; Mode, N.A.; Bæk, R.; Jørgensen, M.M.; Freeman, D.W.; Witwer, K.W.; Zonderman, A.B.; Biragyn, A.; et al. Age-Related Changes in Plasma Extracellular Vesicle Characteristics and Internalization by Leukocytes. Sci. Rep. 2017, 7, 1342.
- Pap, E.; Pállinger, É.; Falus, A.; Kiss, A.A.; Kittel, Á.; Kovács, P.; Buzás, E.I. T Lymphocytes are Targets for Platelet- and Trophoblast-Derived Microvesicles During Pregnancy. Placenta 2008, 29, 826–832.
- Nardi, F.D.S.; Michelon, T.F.; Neumann, J.; Manvailer, L.F.S.; Wagner, B.; Horn, P.A.; Bicalho, M.D.G.; Rebmann, V. High levels of circulating extracellular vesicles with altered expression and function during pregnancy. Immunobiology 2016, 221, 753–760.
- Sarker, S.; Scholz-Romero, K.; Perez, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E.; Salomon, C. Placenta-derived exosomes continuously increase in maternal circulation over the first trimester of pregnancy. J. Transl. Med. 2014, 12, 204.
- Salomon, C.; Torres, M.J.; Kobayashi, M.; Scholz-Romero, K.; Sobrevia, L.; Dobierzewska, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E. A gestational profile of placental exosomes in maternal plasma and their effects on endothelial cell migration. PLoS ONE 2014, 9, e98667.
- Howe, C.G.; Foley, H.B.; Kennedy, E.M.; Eckel, S.P.; Chavez, T.A.; Faham, D.; Grubbs, B.H.; Al-Marayati, L.; Lerner, D.; Suglia, S.; et al. Extracellular vesicle microRNA in early versus late pregnancy with birth outcomes in the MADRES study. Epigenetics 2021.
- Xie, Y.; Gao, Y.; Zhang, L.; Chen, Y.; Ge, W.; Tang, P. Involvement of serum-derived exosomes of elderly patients with bone loss in failure of bone remodeling via alteration of exosomal bone-related proteins. Aging Cell 2018, 17, e12758.
- Alberro, A.; Sáenz-Cuesta, M.; Muñoz-Culla, M.; Mateo-Abad, M.; Gonzalez, E.; Carrasco-Garcia, E.; Araúzo-Bravo, M.J.; Matheu, A.; Vergara, I.; Otaegui, D. Inflammaging and frailty status do not result in an increased extracellular vesicle concentration in circulation. Int. J. Mol. Sci. 2016, 17, 1168.
- Gustafson, C.M.; Shepherd, A.J.; Miller, V.M.; Jayachandran, M. Age- and sex-specific differences in blood-borne microvesicles from apparently healthy humans. Biol. Sex. Differ. 2015, 6, 10.
- Weilner, S.; Keider, V.; Winter, M.; Harreither, E.; Salzer, B.; Weiss, F.; Schraml, E.; Messner, P.; Pietschmann, P.; Hildner, F.; et al. Vesicular Galectin-3 levels decrease with donor age and contribute to the reduced osteo-inductive potential of human plasma derived extracellular vesicles. Aging 2016, 8, 16–33.
- Frühbeis, C.; Helmig, S.; Tug, S.; Simon, P.; Krämer-Albers, E.M. Physical exercise induces rapid release of small extracellular vesicles into the circulation. J. Extracell. Vesicles 2015, 4, 28239.
- Bei, Y.; Xu, T.; Lv, D.; Yu, P.; Xu, J.; Che, L.; Das, A.; Tigges, J.; Toxavidis, V.; Ghiran, I.; et al. Exercise-induced circulating extracellular vesicles protect against cardiac ischemia–reperfusion injury. Basic Res. Cardiol. 2017, 112, 38.
- Hou, Z.; Qin, X.; Hu, Y.; Zhang, X.; Li, G.; Wu, J.; Li, J.; Sha, J.; Chen, J.; Xia, J.; et al. Longterm Exercise-Derived Exosomal miR-342-5p: A Novel Exerkine for Cardioprotection. Circ. Res. 2019, 124, 1386–1400.
- Whitham, M.; Parker, B.L.; Friedrichsen, M.; Hingst, J.R.; Hjorth, M.; Hughes, W.E.; Egan, C.L.; Cron, L.; Watt, K.I.; Kuchel, R.P.; et al. Extracellular Vesicles Provide a Means for Tissue Crosstalk during Exercise. Cell Metab. 2018, 27, 237–251.
- Nederveen, J.P.; Warnier, G.; Di Carlo, A.; Nilsson, M.I.; Tarnopolsky, M.A. Extracellular Vesicles and Exosomes: Insights From Exercise Science. Front. Physiol. 2021, 11, 604274.
- Picca, A.; Guerra, F.; Calvani, R.; Marini, F.; Biancolillo, A.; Landi, G.; Beli, R.; Landi, F.; Bernabei, R.; Bentivoglio, A.R.; et al. Mitochondrial Signatures in Circulating Extracellular Vesicles of Older Adults with Parkinson’s Disease: Results from the EXosomes in PArkiNson’s Disease (EXPAND) Study. J. Clin. Med. 2020, 9, 504.
- Lim, C.Z.J.; Zhang, Y.; Chen, Y.; Zhao, H.; Stephenson, M.C.; Ho, N.R.Y.; Chen, Y.; Chung, J.; Reilhac, A.; Loh, T.P.; et al. Subtyping of circulating exosome-bound amyloid β reflects brain plaque deposition. Nat. Commun. 2019, 10, 1144.
- Lee, S.; Mankhong, S.; Kang, J.H. Extracellular vesicle as a source of alzheimer’s biomarkers: Opportunities and challenges. Int. J. Mol. Sci. 2019, 20, 1728.
- Li, J.J.; Wang, B.; Kodali, M.C.; Chen, C.; Kim, E.; Patters, B.J.; Lan, L.; Kumar, S.; Wang, X.; Yue, J.; et al. In vivo evidence for the contribution of peripheral circulating inflammatory exosomes to neuroinflammation. J. Neuroinflammation 2018, 15, 8.
- Jimenez, J.J.; Minagar, A.; Mauro, L.M.; Jy, W.; Mao, W.W.; Horstman, L.L.; Ahn, Y.S.; Sheremata, W.A. Elevated plasma endothelial microparticles in multiple sclerosis. Neurology 2001, 56, 1319–1324.
- Bhargava, P.; Nogueras-Ortiz, C.; Kim, S.; Delgado-Peraza, F.; Calabresi, P.A.; Kapogiannis, D. Synaptic and complement markers in extracellular vesicles in multiple sclerosis. Mult. Scler. J. 2020, 27, 509–518.
- Sáenz-Cuesta, M.; Alberro, A.; Muñoz-Culla, M.; Osorio-Querejeta, I.; Fernandez-Mercado, M.; Lopetegui, I.; Tainta, M.; Prada, Á.; Castillo-Triviño, T.; Falcón-Pérez, J.M.; et al. The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients. Int. J. Mol. Sci. 2018, 19, 2448.
- Arntz, O.J.; Pieters, B.C.H.; Thurlings, R.M.; Wenink, M.H.; van Lent, P.L.E.M.; Koenders, M.I.; van den Hoogen, F.H.J.; van der Kraan, P.M.; van de Loo, F.A.J. Rheumatoid Arthritis Patients With Circulating Extracellular Vesicles Positive for IgM Rheumatoid Factor Have Higher Disease Activity. Front. Immunol. 2018, 9, 2388.
- Tziatzios, G.; Polymeros, D.; Spathis, A.; Triantafyllou, M.; Gkolfakis, P.; Karakitsos, P.; Dimitriadis, G.; Triantafyllou, K. Increased levels of circulating platelet derived microparticles in Crohn’s disease patients. Scand. J. Gastroenterol. 2016, 51, 1184–1192.
- Simak, J.; Gelderman, M.P.; Yu, H.; Wright, V.; Baird, A.E. Circulating endothelial microparticles in acute ischemic stroke: A link to severity, lesion volume and outcome. J. Thromb. Haemost. 2006, 4, 1296–1302.
- Sinning, J.M.; Losch, J.; Walenta, K.; Böhm, M.; Nickenig, G.; Werner, N. Circulating CD31 +/Annexin V + microparticles correlate with cardiovascular outcomes. Eur. Heart J. 2011, 32, 2034–2041.
- Ye, W.; Tang, X.; Yang, Z.; Liu, C.; Zhang, X.; Jin, J.; Lyu, J. Plasma-derived exosomes contribute to inflammation via the TLR9-NF-κB pathway in chronic heart failure patients. Mol. Immunol. 2017, 87, 114–121.
- Mehaffy, C.; Dobos, K.M.; Nahid, P.; Kruh-Garcia, N.A. Second generation multiple reaction monitoring assays for enhanced detection of ultra-low abundance Mycobacterium tuberculosis peptides in human serum. Clin. Proteom. 2017, 14, 21.
- Arya, R.; Dabral, D.; Faruquee, H.M.; Mazumdar, H.; Patgiri, S.J.; Deka, T.; Basumatary, R.; Kupa, R.U.; Semy, C.; Kapfo, W.; et al. Serum Small Extracellular Vesicles Proteome of Tuberculosis Patients Demonstrated Deregulated Immune Response. Proteom. Clin. Appl. 2020, 14, e1900062.
- Hubert, A.; Subra, C.; Jenabian, M.-A.; Labrecque, P.-F.T.; Tremblay, C.; Laffont, B.; Provost, P.; Routy, J.-P.; Gilbert, C. Elevated Abundance, Size, and MicroRNA Content of Plasma Extracellular Vesicles in Viremic HIV-1+ Patients: Correlations With Known Markers of Disease Progression. J. Acquir. Immune Defic. Syndr. 2015, 70, 219–227.
- Chettimada, S.; Lorenz, D.R.; Misra, V.; Dillon, S.T.; Reeves, R.K.; Manickam, C.; Morgello, S.; Kirk, G.D.; Mehta, S.H.; Gabuzda, D. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci. Rep. 2018, 8, 7227.
- Menon, R.; Debnath, C.; Lai, A.; Guanzon, D.; Bhatnagar, S.; Kshetrapal, P.; Sheller-Miller, S.; Salomon, C. Protein profile changes in circulating placental extracellular vesicles in term and preterm births: A longitudinal study. Endocrinology 2020, 161, bqaa009.
- Li, H.; Ouyang, Y.; Sadovsky, E.; Parks, W.T.; Chu, T.; Sadovsky, Y. Unique microRNA Signals in Plasma Exosomes from Pregnancies Complicated by Preeclampsia. Hypertension 2020, 75, 762–771.
- Buca, D.; Bologna, G.; D’Amico, A.; Cugini, S.; Musca, F.; Febbo, M.; D’Arcangelo, D.; Buca, D.; Simeone, P.; Liberati, M.; et al. Extracellular Vesicles in Feto–Maternal Crosstalk and Pregnancy Disorders. Int. J. Mol. Sci. 2020, 21, 2120.
- Kim, H.K.; Song, K.S.; Park, Y.S.; Kang, Y.H.; Lee, Y.J.; Lee, K.R.; Kim, H.K.; Ryu, K.W.; Bae, J.M.; Kim, S. Elevated levels of circulating platelet microparticles, VEGF, IL-6 and RANTES in patients with gastric cancer: Possible role of a metastasis predictor. Eur. J. Cancer 2003, 39, 184–191.
- Nazari, M.; Javandoost, E.; Talebi, M.; Movassaghpour, A.; Soleimani, M. Platelet microparticle controversial role in cancer. Adv. Pharm. Bull. 2021, 11, 39–55.
- Cappello, F.; Logozzi, M.; Campanella, C.; Bavisotto, C.C.; Marcilla, A.; Properzi, F.; Fais, S. Exosome levels in human body fluids: A tumor marker by themselves? Eur. J. Pharm. Sci. 2017, 96, 93–98.
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N.; et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015, 523, 177–182.
- Liang, K.; Liu, F.; Fan, J.; Sun, D.; Liu, C.; Lyon, C.J.; Bernard, D.W.; Li, Y.; Yokoi, K.; Katz, M.H.; et al. Nanoplasmonic Quantification of Tumor-derived Extracellular Vesicles in Plasma Microsamples for Diagnosis and Treatment Monitoring. Nat. Biomed. Eng. 2017, 1, 0021.
- Yang, K.S.; Im, H.; Hong, S.; Pergolini, I.; del Castillo, A.F.; Wang, R.; Clardy, S.; Huang, C.-H.; Pille, C.; Ferrone, S.; et al. Multi-parametric plasma EV profiling facilitates diagnosis of pancreatic malignancy. Sci. Transl. Med. 2017, 9, eaal3226.
- Skog, J.; Wurdinger, T.; van Rijn, S.; Meijer, D.; Gainche, L.; Sena-Esteves, M.; Curry, W.T., Jr.; Carter, R.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and protein that promote tumor growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476.
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New molecular targets of diseases. Acta Pharmacol. Sin. 2018, 39, 501–513.
- Gimona, M.; Pachler, K.; Laner-Plamberger, S.; Schallmoser, K.; Rohde, E. Manufacturing of human extracellular vesicle-based therapeutics for clinical use. Int. J. Mol. Sci. 2017, 18, 1190.
- Rufino-Ramos, D.; Albuquerque, P.R.; Carmona, V.; Perfeito, R.; Nobre, R.J.; de Almeida, L.P. Extracellular vesicles: Novel promising delivery systems for therapy of brain diseases. J. Control. Release 2017, 262, 247–258.
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156.
- Osorio-Querejeta, I.; Alberro, A.; Muñoz-Culla, M.; Mäger, I.; Otaegui, D. Therapeutic potential of extracellular vesicles for demyelinating diseases; Challenges and opportunities. Front. Mol. Neurosci. 2018, 11, 434.
- Trelis, M.; Galiano, A.; Bolado, A.; Toledo, R.; Marcilla, A.; Bernal, D. Subcutaneous injection of exosomes reduces symptom severity and mortality induced by Echinostoma caproni infection in BALB/c mice. Int. J. Parasitol. 2016, 46, 799–808.
- Shears, R.K.; Bancroft, A.J.; Hughes, G.W.; Grencis, R.K.; Thornton, D.J. Extracellular vesicles induce protective immunity against Trichuris muris. Parasite Immunol. 2018, 40, e12536.
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate immune response in Th1- and Th2-dominant mouse strains. Shock 2004, 22, 460–466.
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345.
- Cooper, J.M.; Wiklander, P.B.O.; Nordin, J.Z.; Al-Shawi, R.; Wood, M.J.; Vithlani, M.; Schapira, A.H.V.; Simons, J.P.; El-Andaloussi, S.; Alvarez-Erviti, L. Systemic exosomal siRNA delivery reduced alpha-synuclein aggregates in brains of transgenic mice. Mov. Disord. 2014, 29, 1476–1485.
- Yang, Y.; Ye, Y.; Kong, C.; Su, X.; Zhang, X.; Bai, W.; He, X. MiR-124 Enriched Exosomes Promoted the M2 Polarization of Microglia and Enhanced Hippocampus Neurogenesis After Traumatic Brain Injury by Inhibiting TLR4 Pathway. Neurochem. Res. 2019, 44, 811–828.
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose Tissue Macrophage-Derived Exosomal miRNAs Can Modulate in Vivo and in Vitro Insulin Sensitivity. Cell 2017, 171, 372–384.
- Castaño, C.; Mirasierra, M.; Vallejo, M.; Novials, A.; Párrizas, M. Delivery of muscle-derived exosomal miRNAs induced by HIIT improves insulin sensitivity through down-regulation of hepatic FoxO1 in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 30335–30343.
- Pêche, H.; Renaudin, K.; Beriou, G.; Merieau, E.; Amigorena, S.; Cuturi, M.C. Induction of tolerance by exosomes and short-term immunosuppression in a fully MHC-mismatched rat cardiac allograft model. Am. J. Transplant. 2006, 6, 1541–1550.
- Ma, B.; Yang, J.Y.; Song, W.J.; Ding, R.; Zhang, Z.C.; Ji, H.C.; Zhang, X.; Wang, J.L.; Yang, X.S.; Tao, K.S.; et al. Combining Exosomes Derived from Immature DCs with Donor Antigen-Specific Treg Cells Induces Tolerance in a Rat Liver Allograft Model. Sci. Rep. 2016, 6, 32971.
- Ren, S.; Chen, J.; Duscher, D.; Liu, Y.; Guo, G.; Kang, Y.; Xiong, H.; Zhan, P.; Wang, Y.; Wang, C.; et al. Microvesicles from human adipose stem cells promote wound healing by optimizing cellular functions via AKT and ERK signaling pathways. Stem Cell Res. Ther. 2019, 10, 47.
- Villa, A.; Garofalo, M.; Crescenti, D.; Rizzi, N.; Brunialti, E.; Vingiani, A.; Belotti, P.; Sposito, C.; Franzè, S.; Cilurzo, F.; et al. Transplantation of autologous extracellular vesicles for cancer-specific targeting. Theranostics 2021, 11, 2034–2047.
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; del Portillo, H.A.; et al. Applying extracellular vesicles based therapeutics in clinical trials—An ISEV position paper. J. Extracell. Vesicles 2015, 4, 30087.
- Mendt, M.; Kamerkar, S.; Sugimoto, H.; McAndrews, K.M.; Wu, C.C.; Gagea, M.; Yang, S.; Blanko, E.V.R.; Peng, Q.; Ma, X.; et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight 2018, 3, e99263.
- Rankin-Turner, S.; Vader, P.; O’Driscoll, L.; Giebel, B.; Heaney, L.M.; Davies, O.G. A call for the standardised reporting of factors affecting the exogenous loading of extracellular vesicles with therapeutic cargos. Adv. Drug Deliv. Rev. 2021, 173, 479–491.
- Wiklander, O.P.B.; Brennan, M.; Lötvall, J.; Breakefield, X.O.; Andaloussi, S.E.L. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, eaav8521.
- Chen, Y.S.; Lin, E.Y.; Chiou, T.W.; Harn, H.J. Exosomes in clinical trial and their production in compliance with good manufacturing practice. Tzu Chi Med. J. 2020, 32, 113–120.

