Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hiroshi Takagi | + 1920 word(s) | 1920 | 2021-08-09 11:28:43 | | | |
| 2 | Amina Yu | Meta information modification | 1920 | 2021-08-18 08:08:04 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Takagi, H. Longevity Regulation in Yeast. Encyclopedia. Available online: https://encyclopedia.pub/entry/13289 (accessed on 29 June 2026).
Takagi H. Longevity Regulation in Yeast. Encyclopedia. Available at: https://encyclopedia.pub/entry/13289. Accessed June 29, 2026.
Takagi, Hiroshi. "Longevity Regulation in Yeast" Encyclopedia, https://encyclopedia.pub/entry/13289 (accessed June 29, 2026).
Takagi, H. (2021, August 18). Longevity Regulation in Yeast. In Encyclopedia. https://encyclopedia.pub/entry/13289
Takagi, Hiroshi. "Longevity Regulation in Yeast." Encyclopedia. Web. 18 August, 2021.
Copy Citation
Mitochondrial energy metabolism is maintained through oxidative degradation of proline and that this process is important in regulating the longevity of yeast cells.
chronological lifespan
energy metabolism
homeostasis
longevity
proline
proline oxidase
Saccharomyces cerevisiae
1. Introduction
Aging is a progressive decline in biological functions occurring in all living organisms, ultimately leading to cell death [1]. Organisms are exposed to various environmental stresses (e.g., nutrient starvation, oxidative stress, temperature shift) during their lifespan and aging is associated with homeostatic imbalances in the form of reduced capacity to respond to various stresses [2][3]. Molecules that work to maintain cellular homeostasis are essential for longevity.
Proline is a multifunctional amino acid in organisms. In addition to being a proteogenic amino acid, proline functions as a stress protectant, namely an osmolyte, oxidative stress protectant, protein-folding chaperone, membrane stabilizer and scavenger of reactive oxygen species [4][5][6][7]. In short, proline is known to be important in maintaining cellular homeostasis by protecting cells from various environmental stresses. Thus, the metabolic regulation of proline, including biosynthesis, degradation and transport, has been of great interest. In the budding yeast Saccharomyces cerevisiae (S. cerevisiae), proline is synthesized from glutamate by three cytoplasmic enzymes, the γ-glutamyl kinase Pro1, the γ-glutamyl phosphate reductase Pro2, and the Δ1-pyrroline-5-carboxylate (P5C) reductase Pro3 [8][9]. In most eukaryotes, including yeast, proline is degraded predominantly in mitochondria. The mitochondrial proline oxidase Put1 converts proline into P5C, which is then converted to glutamate by the P5C dehydrogenase Put2 [10][11]. Proline oxidation by Put1 on the mitochondrial inner membrane may result in the generation of electrons that are then donated to the mitochondrial electron transport chain (ETC) through flavine adenine dinucleotide (FAD) to generate ATP [12][13]. PUT1 and PUT2 are regulated by the transcriptional activator Put3. Although Put3 is constitutively bound to the PUT1 and PUT2 promoters, Put3 is maximally activated for upregulation of PUT1 and PUT2 only in the presence of proline and in the absence of preferred nitrogen sources [14]. In addition to the transcriptional regulation of proline metabolisms, excess proline also inhibits the enzymatic activity in a concentration-dependent manner [15]. Unlike bacteria and plants, yeasts do not increase their intracellular proline level under various stress conditions. However, our previous study revealed that the Ile150Thr variant of Pro1 was less sensitive to negative feedback inhibition by proline [15]. Thus, yeast cells accumulate proline by expressing the mutant PRO1 gene encoding the Ile150Thr variant. Yeast cells with proline accumulation have been shown to exhibit an increase in stress tolerance to freezing, desiccation, oxidation, and ethanol [16].
The aging of yeast cells is measured in two different ways: the replicative lifespan and the chronological lifespan [17]. The replicative lifespan is defined as the number of mitotic divisions before senescence and is used as a model to study the aging of proliferative cells (stem cells, etc.). During the process of mitotic division, yeast cells are exposed to various stresses such as osmotic shock. Therefore, stress protectants in cells are important for replicative lifespan regulation. In fact, we recently reported that intracellular proline levels regulate the replicative lifespan in yeast [18]. In contrast, chronological lifespan is the time that yeast cells survive under non-proliferative conditions (namely, after the stationary phase) and this period models the aging of postmitotic mammalian cells (neuron cells, etc.). After the stationary phase, yeast cells are exposed to nutrient starvation; thus, energy homeostasis in cells is known to be critical for longevity. Basically, the quality and quantity of nutrients in the environment are key factors of longevity regulation. It is common knowledge that calorie restriction extends longevity [19]. The cellular response to calorie restriction occurs partly through nutrient-sensing mechanisms containing the target of rapamycin, cAMP-PKA, and/or the Snf1 pathways [20]. These signals maintain cellular homeostasis by controlling energy metabolism, stress responses and so on, leading to regulation of the lifespan.
In this study, we discovered that Put1 is important for the chronological lifespan of yeast. During the aging period, PUT1 expression was positively regulated by Put3 and was required for longevity. More importantly, the oxidation of proline by Put1 helped maintain mitochondrial membrane potential and ATP production, leading to an increase in lifespan. Our results suggest a possible mechanism for longevity regulation via proline metabolism in yeast.
2. Discussion
In this study, we discovered that the proline oxidase Put1 contributes to the chronological lifespan of yeast. Upon yeast cells entering the aging period, PUT1 expression was positively regulated by Put3 and was associated with longevity. More intriguingly, Put1 helped sustain mitochondrial membrane potential and ATP production, leading to maintenance of the energy metabolism. Our present data suggest a possible mechanism for the longevity regulation via proline metabolism (Figure 1). Yeast cells normally produce ATP from glucose and ethanol via glycolysis in the cytosol and/or ETC in mitochondria, respectively. When all glucose and ethanol are consumed, yeast cells probably uptake proline from the environment and start to utilize proline as an energy source via its oxidation by Put1. This conversion of proline into P5C results in the generation of electrons that are donated to ETC through FAD to generate ATP. The mechanism functions for the homeostasis of energy metabolism, which contributes to longevity.
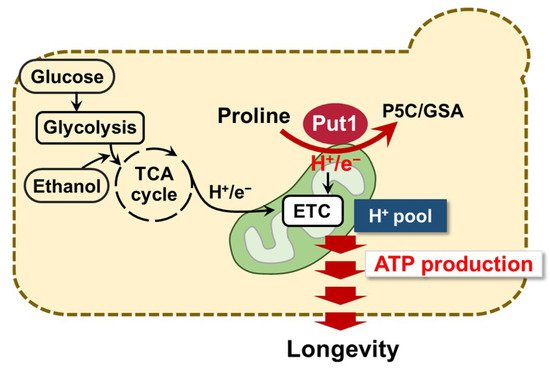
Figure 1. Schematic model of longevity regulation by proline oxidation. In the absence of principal energy sources, such as glucose and ethanol, yeast cells start to utilize proline as an energy source via the Put1-catylatic reaction. This reaction results in the generation of electrons which are donated to ETC, which generates ATP. Such a mechanism may contribute to longevity.
Proline is a multifunctional amino acid in many organisms. In addition to being a proteogenic amino acid, proline functions as a stress protectant, namely an osmolyte, oxidative stress protectant, protein-folding chaperone, membrane stabilizer and scavenger of reactive oxygen species [4][5][6][7]. Therefore, proline itself may protect yeast cells against various stresses caused by aging and regulate the longevity. However, our present study showed that the amount of proline in the cell does not affect the lifespan. In addition, proline addition had no effect on the lifespan of put1Δ cells. These results indicated that the use of proline for ATP synthesis is more important for longevity than as a protective molecule. Aged cells are known to accumulate misfolded proteins or protein aggregates inside the cell due to downregulation of protein quality control systems [21]. Molecular chaperones are central components of such systems and require ATP for protein refolding. Thus, ATP synthesized via proline oxidation catalyzed by Put1 may be used to support the activity of molecular chaperones during aging period, leading to removal of unfolded proteins and prolongation of the longevity. Further studies are needed to reveal that the proline catabolism helps the chaperone activity under aging.
Since yeast cells can utilize various nitrogen sources, the sensing of nitrogen sources is important for optimizing cellular metabolism. Generally, the presence of a preferred nitrogen source, such as ammonium ions or glutamine, represses the expression of catabolic genes compared to nonpreferred nitrogen sources, such as proline and leucine, through nitrogen catabolite repression (NCR) [22][23][24]. All of the proline-metabolizing genes are NCR-regulated genes; that is, their transcription is repressed by the NCR repressors Ure2 and Dal80 when a preferred nitrogen source is present [25]. In addition, the proline transporters Gap1 and Put4 are well-known to be subjected to post-translational regulation, which suppresses their transport activity by the ubiquitin ligase Rsp5 [26][27]. Thus, proline appears to be consumed, similar to other nonpreferred nitrogen sources. In fact, however, yeast cells rarely uptake and consume extracellular proline, even though they do use other nonpreferred nitrogen sources, such as leucine and 4-aminobutanoic acid [28][29][30]. In short, both the uptake and metabolism of proline are strongly inhibited in cells compared with other nonpreferred nitrogen sources. In light of our present study, yeast cells may adopt a strategy of leaving proline outside the cell as a last resort for survival. The presence of energy sources such as glucose strongly inhibits the uptake of proline. In the absence of other energy sources, yeast cells may uptake proline via Gap1 and Put4 and oxidize proline by Put1, which produces ATP to assist in cellular survival. This suggests that proline metabolism is not only regulated by nitrogen sources but also by energy sources such as glucose. Our previous study showed that PUT1 induction is positively regulated by the general stress transcription factors Msn2/4 [18]. The important fact is that Msn2/4 is activated by the low-glucose sensor Snf1 [31]. Hence, Snf1 may sense energy source depletion in the environment and subsequently Msn2/4 and Put3 may work cooperatively to activate proline metabolism, leading to maintenance of energy homeostasis. Further studies will be needed for understanding the detailed mechanism and physiological significance of proline-mediated energy homeostasis.
In this study, we found that proline taken up from outside the cell, rather than proline synthesized inside the cell, contributes to the longevity of yeast. On the other hand, a previous study indicated that the intracellular proline regulates the replicative lifespan of yeast [18]. Generally, proline is present in the cytosol, but most of excess proline is stored in the vacuoles [32]. Since a mechanism for active transports of proline from the vacuoles to the cytosol has not been identified, proline in the vacuoles may remain there indefinitely. The most severe stress under replicative events is considered to be osmotic stress, and vacuoles are important organelles for the adaptation to osmotic pressure [33]. Thus, proline maintained in vacuoles may be functional during the replicative lifespan. In the case of regulating the chronological lifespan, however, proline must be transported from the cytosol to the mitochondria to serve as a substrate for Put1. Therefore, it is likely to be important for yeast cells to uptake extracellular proline upon entering the aging period.
Proline metabolisms is highly conserved across various organisms. In terms of proline degradation, the localization of this process and the properties of the enzymes involved are very similar between yeasts and mammals. It is known that the mitochondrial proline oxidase is implicated in supporting ATP production, protein synthesis, and redox homeostasis in cancer cells [34]. One of the most lethal capacities of cancer cells is their ability to metastasize to distant organs. Recently, Elia et al. suggested that proline metabolism supports the metastatic cascade via the production of ATP [35]. Importantly, cancer cells in metastatic tissue exhibit higher expression of proline oxidase than normal cells. Accordingly, inhibition of proline oxidase activity impairs metastasis formation in different kinds of metastatic breast cancer [35]. However, the details of how ATP production is involved in metastatic progression are still unknown. Considering our present data, proline oxidase may function as an enzyme to prevent cellular senescence mediated by ATP production. Likewise, inhibition of senescence is well-known to induce cancer formation and metastasis [36]. Thus, an inhibitor of proline oxidase would be a promising candidate as a drug that interferes with the metastatic cascade. Yeast cells can grow on the medium containing proline as a sole nitrogen source, but Put1, an orthologue of proline oxidase, is essential for such growth. Therefore, inhibitors of proline oxidase may be screened by a chemical biology-based study utilizing yeast.
The aging process is a part of the life cycle of all organisms. Yeasts are very important microorganisms for various industrial fields, such as the production of fermented foods and useful compounds. A high fermentative capacity is a key factor for their biotechnological application. Since the yeast performance in biotechnological applications is dependent on levels of cell viability and vitality, the extension of the lifespan is important to maximize these processes. Hence, isolating and/or engineering yeast strains with the appropriate localization and concentration of proline could contribute to improvements in fermentation and other kinds of biochemical productivity.
References
- Jones, O.R.; Scheuerlein, A.; Salguero-Gómez, R.; Camarda, C.G.; Schaible, R.; Casper, B.B.; Dahlgren, J.P.; Ehrlén, J.; García, M.B.; Menges, E.S.; et al. Diversity of ageing across the tree of life. Nature 2014, 505, 169–173.
- Pomatto, L.C.D.; Davies, K.J.A. The role of declining adaptive homeostasis in ageing. J. Physiol. 2017, 595, 7275–7309.
- Mouton, S.N.; Thaller, D.J.; Crane, M.M.; Rempel, I.L.; Terpstra, O.T.; Steen, A.; Kaeberlein, M.; Lusk, C.P.; Boersma, A.J.; Veenhoff, L.M. A physicochemical perspective of aging from single-cell analysis of pH, macromolecular and organellar crowding in yeast. eLife 2020, 9, e54707.
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011.
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466.
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11.
- Mat Nanyan, N.S.B.; Takagi, H. Proline homeostasis in Saccharomyces cerevisiae: How does the stress-responsive transcription factor Msn2 play a role? Front. Genet. 2020, 11, 438.
- Tomenchok, D.M.; Brandriss, M.C. Gene-enzyme relationships in the proline biosynthetic pathway of Saccharomyces cerevisiae. J. Bacteriol. 1987, 169, 5364–5372.
- Brandriss, M.C.; Falvey, D.A. Proline biosynthesis in Saccharomyces cerevisiae: Analysis of the PRO3 gene, which encodes delta 1-pyrroline-5-carboxylate reductase. J. Bacteriol. 1992, 174, 3782–3788.
- Wang, S.S.; Brandriss, M.C. Proline utilization in Saccharomyces cerevisiae: Analysis of the cloned PUT1 gene. Mol. Cell. Biol. 1986, 6, 2638–2645.
- Brandriss, M.C. Proline utilization in Saccharomyces cerevisiae: Analysis of the cloned PUT2 gene. Mol. Cell. Biol. 1983, 3, 1846–1856.
- Pandhare, J.; Donald, S.P.; Cooper, S.K.; Phang, J.M. Regulation and function of proline oxidase under nutrient stress. J. Cell. Biochem. 2009, 107, 759–768.
- Hancock, C.N.; Liu, W.; Alvord, W.G.; Phang, J.M. Co-regulation of mitochondrial respiration by proline dehydrogenase/oxidase and succinate. Amino Acids 2016, 48, 859–872.
- Huang, H.L.; Brandriss, M.C. The regulator of the yeast proline utilization pathway is differentially phosphorylated in response to the quality of the nitrogen source. Mol. Cell. Biol. 2000, 20, 892–899.
- Sekine, T.; Kawaguchi, A.; Hamano, Y.; Takagi, H. Desensitization of feedback inhibition of the Saccharomyces cerevisiae gamma-glutamyl kinase enhances proline accumulation and freezing tolerance. Appl. Environ. Microbiol. 2007, 73, 4011–4019.
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223.
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab. 2012, 16, 18–31.
- Mukai, Y.; Kamei, Y.; Liu, X.; Jiang, S.; Sugimoto, Y.; Mat Nanyan, N.S.B.; Watanabe, D.; Takagi, H. Proline metabolism regulates replicative lifespan in the yeast Saccharomyces cerevisiae. Microb. Cell 2019, 6, 482–490.
- Lin, S.J.; Defossez, P.A.; Guarente, L. Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science 2000, 289, 2126–2128.
- Hwangbo, D.-S.; Lee, H.-Y.; Abozaid, L.S.; Min, K.-J. Mechanisms of lifespan regulation by calorie restriction and intermittent fasting in model organisms. Nutrients 2020, 12, 1194.
- Krisko, A.; Radman, M. Protein damage, ageing and age-related diseases. Open Biol. 2019, 9, 180249.
- Dubois, E.; Vissers, S.; Grenson, M.; Wiame, J.M. Glutamine and ammonia in nitrogen catabolite repression of Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 1977, 75, 233–239.
- Hofman-Bang, J. Nitrogen catabolite repression in Saccharomyces cerevisiae. Mol. Biotechnol. 1999, 12, 35–73.
- Zhang, W.; Du, G.; Zhou, J.; Chen, J. Regulation of sensing, transportation, and catabolism of nitrogen sources in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2018, 82.
- Coffman, J.A.; Rai, R.; Cooper, T.G. Genetic evidence for Gln3p-independent, nitrogen catabolite repression-sensitive gene expression in Saccharomyces cerevisiae. J. Bacteriol. 1995, 177, 6910–6918.
- Villers, J.; Savocco, J.; Szopinska, A.; Degand, H.; Nootens, S.; Morsomme, P. Study of the plasma membrane proteome dynamics reveals novel targets of the nitrogen regulation in yeast. Mol. Cell. Proteom. 2017, 16, 1652–1668.
- Nishimura, A.; Tanahashi, R.; Takagi, H. The yeast α-arrestin Art3 is a key regulator for arginine-induced endocytosis of the high-affinity proline transporter Put4. Biochem. Biophys. Res. Commun. 2020, 531, 416–421.
- Hill, A.E.; Stewart, G.G. Free amino nitrogen in brewing. Fermentation 2019, 5, 22.
- Valero, E.; Millán, C.; Ortega, J.M.; Mauricio, J.C. Concentration of amino acids in wine after the end of fermentation by Saccharomyces cerevisiae strains. J. Sci. Food Agric. 2003, 83, 830–835.
- Nishimura, A.; Tanikawa, T.; Takagi, H. Inhibitory effect of arginine on proline utilization in Saccharomyces cerevisiae. Yeast 2020, 37, 531–540.
- Li, L.; Kaplan, J.; Ward, D.M. The glucose sensor Snf1 and the transcription factors Msn2 and Msn4 regulate transcription of the vacuolar iron importer gene CCC1 and iron resistance in yeast. J. Biol. Chem. 2017, 292, 15577–15586.
- Nishida, I.; Watanabe, D.; Tsolmonbaatar, A.; Kaino, T.; Ohtsu, I.; Takagi, H. Vacuolar amino acid transporters upregulated by exogenous proline and involved in cellular localization of proline in Saccharomyces cerevisiae. J. Gen. Appl. Microbiol. 2016, 62, 132–139.
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372.
- Tanner, J.J.; Fendt, S.-M.; Becker, D.F. The proline cycle as a potential cancer therapy target. Biochemistry 2018, 57, 3433–3444.
- Elia, I.; Broekaert, D.; Christen, S.; Boon, R.; Radaelli, E.; Orth, M.F.; Verfaillie, C.; Grünewald, T.G.P.; Fendt, S.M. Proline metabolism supports metastasis formation and could be inhibited to selectively target metastasizing cancer cells. Nat. Commun. 2017, 8, 15267.
- Ito, T.; Igaki, T. Yorkie drives ras-induced tumor progression by microrna-mediated inhibition of cellular senescence. Sci. Signal. 2021, 14, eaaz3578.
More
Information
Subjects:
Microbiology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
856
Revisions:
2 times
(View History)
Update Date:
18 Aug 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No