 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Yosuke Kurashima | + 3039 word(s) | 3039 | 2021-07-09 08:10:23 | | | |
| 2 | Lindsay Dong | Meta information modification | 3039 | 2021-08-03 08:35:16 | | | | |
| 3 | Conner Chen | Meta information modification | 3039 | 2021-10-12 10:58:27 | | |
Video Upload Options
It is well known that mast cells (MCs) initiate type I allergic reactions and inflammation in a quick response to the various stimulants, including—but not limited to—allergens, pathogen-associated molecular patterns (PAMPs), and damage-associated molecular patterns (DAMPs). MCs highly express receptors of these ligands and proteases (e.g., tryptase, chymase) and cytokines (TNF), and other granular components (e.g., histamine and serotonin) and aggravate the allergic reaction and inflammation. On the other hand, accumulated evidence has revealed that MCs also possess immune-regulatory functions, suppressing chronic inflammation and allergic reactions on some occasions. IL-2 and IL-10 released from MCs inhibit excessive immune responses.
1. Introduction
Mast cells (MCs) are a type of innate immune cell that belongs to the myeloid lineage. It is generally believed that MCs from both humans and rodents are derived from hematopoietic stem cells (HSCs). Mast cell progenitors (MCPs) leave the bone marrow as immature cells and enter the blood circulation, with the help of surface molecules, such as α4β7 integrin, MAdCAM-1 and VCAM1, to migrate to various target tissues [1][2]. MCs have high plasticity and heterogeneity due to their unique process of maturation.
MCs are also regulators of inflammatory disorders and fibrosis occurred in various organs. Associations between MCs recruitment/infiltration and fibrosis have been found in various tissues [3]. Current studies have found that many MCs products, including—but not limited to—tryptase, chymase, histamine, TGF- β1, IL-13, IL-9, CCL2, platelet-derived growth factor (PDGF), glycosaminoglycan and fibroblast growth factor-2 (FGF-2) can promote fibrosis [3]. MCs have long been regarded as the initiators of allergy and inflammation, as well as the promoters of fibrotic diseases, which are pathogenic. However, as every coin has two sides, in addition to some harmful effects, MCs possess anti-allergic and inflammatory effects. MCs appear to play an immunomodulatory role in allergic, acute, and chronic inflammation (e.g., fibrosis [4][5]). There is growing evidence that MCs play an enormous role in allergic responses, inflammatory responses, and wound healing [4][6][7].
2. Origin and Heterogeneity of MCs
It is generally believed that MCs from both humans and rodents are derived from HSCs. In the bone marrow, HSCs first grow into myeloid progenitors and then differentiate into MCPs [8]. Next, MCPs transfer from the bone marrow as immature cells into the bloodstream. Eventually, with the help of surface molecules (e.g., α4β7 integrin, MAdCAM-1, and VCAM1) [2], MCPs migrate to various target tissues, such as the serous cavity, in close contact with the micro-environment (e.g., the skin, gastrointestinal tract, upper airways and lungs [9]) and some vascularized organs (e.g., the liver and kidneys) [10][11]. These tissues contain various tissue-specific factors (e.g., cytokines, growth factors and extracellular matrix [ECM]), which help MCPs finally become phenotypically mature and perform different functions [12].
However, the exact origin of MCs remains a matter of debate. It is generally believed that MCPs in mice are derived from bone marrow. It has also been reported that MCs are developed from the common myeloid progenitor cells (CMPs) [13]. However, Dahlin et al., who demonstrated that MCPs were derived from multipotential progenitor cells (MMPs) rather than CMPs, disagreed [14]. Recent studies have found that mouse MCs have dual developmental origins. A portion of the MCs have a primitive origin. They are derived from the yolk sac during embryogenesis. MCPs derived from the yolk sac migrate to different connective tissues, such as the skin. Another part of adult definitive MCs comes from the HSCs of the aortic-gonad-mesonephros vascular endothelium [15]. Studies have also shown that MCs in adult tissues are supplemented by the proliferation/differentiation of resident precursors in long-lived tissues [16]. Thus, in mouse-based studies, MCs have at least two maturation pathways; however, whether these pathways apply to other mammals, such as humans, has not been fully clarified.
MCs also have high plasticity and heterogeneity due to their unique process of maturation. They show subtype-dependent differences in cell morphology, histochemical characteristics, granular protease expression, function, and survival according to the microenvironment, activating factors and the cytokine milieu [17].
According to the traditional classification system, MCs are divided into two subsets in mice: MMCs and CTMCs. The two subsets have different anatomical localization and protease expression patterns [8]. CTMCs, as the name suggests, are mainly located in the connective tissue of the intestinal submucosa and muscularis propria, the peritoneal cavity, and skin. This subset expresses both chymase and tryptase [14]. Their cytoplasm also contains heparin proteoglycan and a high level of histamine [18]. In contrast, MMCs are a chymase-expressing type [14]. They are usually present in the mucosal tissues of the lung and gastrointestinal tract, with little or no heparin proteoglycans in their granules, and have lower amounts of histamine [18].
Similarly, in humans, the MCs subtypes are classified according to whether they secrete both tryptase and chymase (MCTC) or tryptase alone (MCT). The former is comparable with mouse MMCs, while the latter corresponds to CTMCs [17][18]. The MCTCs are mainly distributed in the skin, the gastrointestinal tract, and conjunctiva and the MCTs can be found in the lungs, nose and sinuses [19].
3. Pro-Allergic and Inflammatory Actions of MCs
3.1. Regulation of Receptor-Mediated MCs Activation
As one of the most common immune cells, the main characteristics of MCs are their pro-allergy and pro-inflammatory effects. MCs can express a variety of receptors, which, when combined with corresponding ligands, can induce the activation of MCs, thereby triggering various pathways. MCs activation mainly occurs by IgE-dependent and IgE-independent pathways [20].
MCs degranulation mediated by the IgE high-affinity receptor (FcεRI) is the most classical mechanism of MCs activation. Immunoglobulin E (IgE) is a class of antibodies produced by plasma cells that shows high affinity to MCs, and which can mediate Type I hypersensitivity reactions, such as food allergy and asthma. The first pathogenic step of IgE-mediated type I hypersensitivity reactions is sensitization (Figure 1A). When the body’s first contact with allergens, antigen-presenting cells like monocytes-macrophages and dendritic cells (DCs) present antigen information to T helper lymphocytes, causing them to secrete cytokines. B lymphocytes then transform into plasma cells and secrete various allergen-specific IgE in response to the cytokines derived from T helper lymphocytes [21]. Specific IgE can bind to FcεRI on the surface of MCs. FcεRI is a type of high-affinity receptor of IgE that exists in the form of a trimer or tetramer. FcεRI contains one α chain, two identical γ chains and one β chain which is sometimes missing. In humans, FcεRI can be expressed as both αβγ2 and αγ2. In rodents, however, FcεRI is only expressed in the form of αβγ2 [21][22]. The extracellular domain of the α chain can bind to the Fc segment of IgE, which is a critical site for triggering allergic reactions. The primary role of the β subunit is to enhance the tyrosine kinase activity and calcium influx and then to amplify the expression of FcεRI on the surface of MCs [14]. When the allergens enter again, FcεRI/IgE complexes are cross-linked with high-affinity antigens on the surface of sensitive MCs, the FcεRI receptor will be activated, causing signal transduction in MCs and promoting the degranulation of MCs and the subsequent release of inflammatory mediators, like histamine, serotonin and leukotriene, which are involved allergic reactions or inflammation [21].
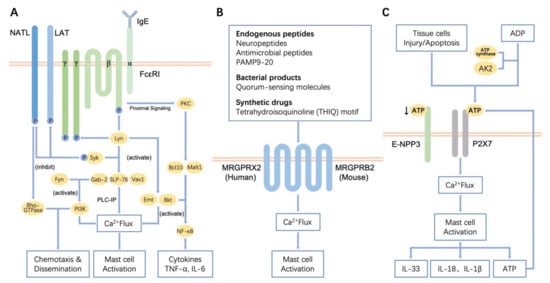
3.2. Fibrogenic Actions of MCs
4. Regulatory-Type Actions of MCs in Allergy and Inflammation
MCs have long been regarded as the initiators of immunity and inflammation, which is pathogenic. For example, specific MCs proteases can promote inflammation, such as tryptase, chymase and carboxypeptidase A3 [46][47][48][49]. However, there are two sides to every coin. In addition to some harmful effects, the benefits of MCs to the human body cannot be ignored. MCs appear to play an immunomodulatory role in inflammation, allergy and fibrosis.
4.1. Regulation of Chronic Inflammation/Fibrosis/Wound Healing
Many results show that while the depletion of MCs cannot prevent tissue repair and remodeling, it can delay wound healing [7][50]. Hence, MCs can promote wound healing. It has been pointed out that MCs are involved in wound healing at several stages [7]. Under the influence of SCF released by keratinocytes as well as CCL2 (MCP-1) and IL-33, MCs gather at the edge of the wound in the first few days [51]. The TNF secreted by them can enhance the expression of XIIIa factor in dermal dendritic cells and then promote hemostasis and clot formation, which help reduce injury [52]. Through the secretion of histamine, lipid mediators, and VEGF, they increase vascular permeability and recruit other cells, such as neutrophils, to help heal the wound (Figure 2) [3]. In the proliferation stage of wound healing, the migration and proliferation of fibroblasts and the formation of collagen fibers are the key steps of wound healing [3]. MCs can release a variety of substances that interact with fibroblasts to promote wound healing (Figure 2). As mentioned before, proteases released by MCs can chemotaxis fibroblasts and promote their mitosis [39][53]. In addition, VEGF, IL-4 and basic fibroblast growth factor (bFGF) derived from MCs can stimulate the proliferation of fibroblasts [54].
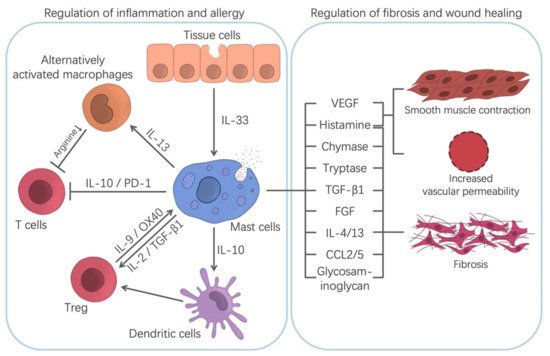
4.2. Regulation of Allergic and Inflammatory Diseases
There is growing evidence that MCs play an important role in inhibiting allergic reactions and inflammation. It has been found that oral immunotherapy (OIT)-induced desensitized MCs have a robust regulatory function and can cooperate with Tregs to form a regulatory network, which helps control food allergy [6]. It has been reported that the anti-allergic part of MCs is mainly realized through the production of regulatory cytokines, such as IL-2 and IL-10 [6]. Several studies have found that the inhibition of the immune responses in the skin, intestines and bladder by MCs depends on IL-10 [19]. MCs-derived IL-10 can induce the production of Tregs by DCs (Figure 2). However, how the immunosuppressive effect of IL-10 is regulated at the molecular level remains to be determined. Thymic-derived Tregs are a type of suppressive T cell, which accounts for 5–10% of circulating CD4+T cells [58]. Studies have shown that Tregs can reduce the production of allergen-specific IgE and pathogenic Th2 and inhibit the degranulation of MCs and basophils, which push forward an immense influence on the control of allergic symptoms [6][59]. Tregs impairment is related to a loss of tolerance, autoimmunity and allergy [59]. In a similar manner, IL-2 is also involved in MCs-mediated immunosuppression and is critical for the development, amplification, activity and survival of Tregs [60]. IL-2 secreted by MCs can effectively expand Tregs populations and then inhibit the immune response. Another cytokine, IL-33, can indirectly participate in the immunosuppressive response by stimulating MCs to secrete IL-2, promoting ST2-independent immunosuppression in Tregs (Figure 2) [61][62]. Meanwhile, some studies have shown that IL-33 can promote the secretion of IL-13 by MCs, which inhibits the production of IL-12 by DCs in the skin. This can hinder the Th1 cell response to cutaneous antigen exposure [63].
References
- Collington, S.J.; Williams, T.J.; Weller, C.L. Mechanisms underlying the localization of mast cells in tissues. Trends Immunol. 2011, 32, 478–485.
- Gurish, M.F.; Tao, H.; Abonia, J.P.; Arya, A.; Friend, D.S.; Parker, C.M.; Austen, K.F. Intestinal mast cell progenitors require CD49dbeta7 (alpha4beta7 integrin) for tissue-specific homing. J. Exp. Med. 2001, 194, 1243–1252.
- Overed-Sayer, C.; Rapley, L.; Mustelin, T.; Clarke, D.L. Are mast cells instrumental for fibrotic diseases? Front. Pharmacol. 2013, 4, 174.
- Lu, L.F.; Lind, E.F.; Gondek, D.C.; Bennett, K.A.; Gleeson, M.W.; Pino-Lagos, K.; Scott, Z.A.; Coyle, A.J.; Reed, J.L.; Van Snick, J.; et al. Mast cells are essential intermediaries in regulatory T-cell tolerance. Nature 2006, 442, 997–1002.
- He, Z.; Song, J.; Hua, J.; Yang, M.; Ma, Y.; Yu, T.; Feng, J.; Liu, B.; Wang, X.; Li, Y.; et al. Mast cells are essential intermediaries in regulating IL-33/ST2 signaling for an immune network favorable to mucosal healing in experimentally inflamed colons. Cell Death Dis. 2018, 9, 1173.
- Takasato, Y.; Kurashima, Y.; Kiuchi, M.; Hirahara, K.; Murasaki, S.; Arai, F.; Izawa, K.; Kaitani, A.; Shimada, K.; Saito, Y.; et al. Orally desensitized mast cells form a regulatory network with Treg cells for the control of food allergy. Mucosal Immunol. 2021, 14, 640–651.
- Wulff, B.C.; Wilgus, T.A. Mast cell activity in the healing wound: More than meets the eye? Exp. Dermatol. 2013, 22, 507–510.
- Hallgren, J.; Gurish, M.F. Pathways of murine mast cell development and trafficking: Tracking the roots and routes of the mast cell. Immunol. Rev. 2007, 217, 8–18.
- Metz, M.; Maurer, M. Mast cells–key effector cells in immune responses. Trends Immunol. 2007, 28, 234–241.
- Ehara, T.; Shigematsu, H. Mast cells in the kidney. Nephrology 2003, 8, 130–138.
- Farrell, D.J.; Hines, J.E.; Walls, A.F.; Kelly, P.J.; Burt, A.D. Intrahepatic mast cells in chronic liver diseases. Hepatology 2010, 22, 1175–1181.
- Weiskirchen, R.; Meurer, S.K.; Liedtke, C.; Huber, M. Mast cells in liver fibrogenesis. Cells 2019, 8, 1429.
- Voehringer, D. Protective and pathological roles of mast cells and basophils. Nat. Rev. Immunol. 2013, 13, 362–375.
- Elieh Ali Komi, D.; Wohrl, S.; Bielory, L. Mast cell biology at molecular level: A comprehensive review. Clin. Rev. Allergy. Immunol. 2020, 58, 342–365.
- Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajenoff, M. Hemogenic endothelial fate mapping reveals dual developmental origin of mast cells. Immunity 2018, 48, 1160–1171.
- Nilsson, G.; Dahlin, J.S. New insights into the origin of mast cells. Allergy 2019, 74, 844–845.
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat. Immunol. 2011, 12, 1035–1044.
- Gurish, M.F.; Austen, K.F. Developmental origin and functional specialization of mast cell subsets. Immunity 2012, 37, 25–33.
- Galli, S.J.; Gaudenzio, N.; Tsai, M. Mast cells in inflammation and disease: Recent progress and ongoing concerns. Annu. Rev. Immunol. 2020, 38, 49–77.
- Bulfone-Paus, S.; Nilsson, G.; Draber, P.; Blank, U.; Levi-Schaffer, F. Positive and negative signals in mast cell activation. Trends Immunol. 2017, 38, 657–667.
- Pritchard, D.I.; Falcone, F.H.; Mitchell, P.D. The evolution of IgE-mediated type I hypersensitivity and its immunological value. Allergy 2021, 76, 1024–1040.
- Cauvi, D.M.; Tian, X.; von Loehneysen, K.; Robertson, M.W. Transport of the IgE receptor alpha-chain is controlled by a multicomponent intracellular retention signal. J. Biol. Chem. 2006, 281, 10448–10460.
- Saitoh, S.; Arudchandran, R.; Manetz, T.S.; Zhang, W.; Sommers, C.L.; Love, P.E.; Rivera, J.; Samelson, L.E. LAT is essential for FcεRI-mediated mast cell activation. Immunity 2000, 12, 525–535.
- Parravicini, V.; Gadina, M.; Kovarova, M.; Odom, S.; Gonzalez-Espinosa, C.; Furumoto, Y.; Saitoh, S.; Samelson, L.E.; O’Shea, J.J.; Rivera, J. Fyn kinase initiates complementary signals required for IgE-dependent mast cell degranulation. Nat. Immunol. 2002, 3, 741–748.
- Moretti, S.; Renga, G.; Oikonomou, V.; Galosi, C.; Pariano, M.; Iannitti, R.G.; Borghi, M.; Puccetti, M.; De Zuani, M.; Pucillo, C.E.; et al. A mast cell-ILC2-Th9 pathway promotes lung inflammation in cystic fibrosis. Nat. Commun 2017, 8, 14017.
- Lipitsa, T.; Naukkarinen, A.; Laitala, J.; Harvima, I.T. Complement C3 is expressed by mast cells in cutaneous vasculitis and is degraded by chymase. Arch. Dermatol. Res. 2016, 308, 575–584.
- Ali, H. Regulation of human mast cell and basophil function by anaphylatoxins C3a and C5a. Immunol. Lett. 2010, 128, 36–45.
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A mast-cell-specific receptor mediates neurogenic inflammation and pain. Neuron 2019, 101, 412–420.e3.
- Zhang, Y.; Wang, J.; Ge, S.; Zeng, Y.; Wang, N.; Wu, Y. Roxithromycin inhibits compound 48/80-induced pseudo-allergy via the MrgprX2 pathway both in vitro and in vivo. Cell. Immunol. 2020, 358, 104239.
- Wolf, K.; Kuhn, H.; Boehm, F.; Gebhardt, L.; Glaudo, M.; Agelopoulos, K.; Stander, S.; Ectors, P.; Zahn, D.; Riedel, Y.K.; et al. A group of cationic amphiphilic drugs activates MRGPRX2 and induces scratching behavior in mice. J. Allergy Clin. Immunol. 2021, in press.
- Wilhelm, K.; Ganesan, J.; Muller, T.; Durr, C.; Grimm, M.; Beilhack, A.; Krempl, C.D.; Sorichter, S.; Gerlach, U.V.; Juttner, E.; et al. Graft-versus-host disease is enhanced by extracellular ATP activating P2X7R. Nat. Med. 2010, 16, 1434–1438.
- Kurashima, Y.; Amiya, T.; Fujisawa, K.; Shibata, N.; Suzuki, Y.; Kogure, Y.; Hashimoto, E.; Otsuka, A.; Kabashima, K.; Sato, S.; et al. The enzyme Cyp26b1 mediates inhibition of mast cell activation by fibroblasts to maintain skin-barrier homeostasis. Immunity 2014, 40, 530–541.
- Kurashima, Y.; Amiya, T.; Nochi, T.; Fujisawa, K.; Haraguchi, T.; Iba, H.; Tsutsui, H.; Sato, S.; Nakajima, S.; Iijima, H.; et al. Extracellular ATP mediates mast cell-dependent intestinal inflammation through P2X7 purinoceptors. Nat. Commun. 2012, 3, 1034.
- Liu, B.; Yang, M.Q.; Yu, T.Y.; Yin, Y.Y.; Liu, Y.; Wang, X.D.; He, Z.G.; Yin, L.; Chen, C.Q.; Li, J.Y. Mast cell tryptase promotes inflammatory bowel disease-induced intestinal fibrosis. Inflamm. Bowel Dis. 2021, 27, 242–255.
- Miyazawa, S.; Hotta, O.; Doi, N.; Natori, Y.; Nishikawa, K.; Natori, Y. Role of mast cells in the development of renal fibrosis: Use of mast cell-deficient rats. Kidney Int. 2004, 65, 2228–2237.
- Kong, P.; Christia, P.; Frangogiannis, N.G. The pathogenesis of cardiac fibrosis. Cell. Mol. Life Sci 2014, 71, 549–574.
- Shimbori, C.; Upagupta, C.; Bellaye, P.S.; Ayaub, E.A.; Sato, S.; Yanagihara, T.; Zhou, Q.; Ognjanovic, A.; Ask, K.; Gauldie, J.; et al. Mechanical stress-induced mast cell degranulation activates TGF-beta1 signalling pathway in pulmonary fibrosis. Thorax 2019, 74, 455–465.
- Hargrove, L.; Kennedy, L.; Demieville, J.; Jones, H.; Meng, F.; DeMorrow, S.; Karstens, W.; Madeka, T.; Greene, J., Jr.; Francis, H. Bile duct ligation-induced biliary hyperplasia, hepatic injury, and fibrosis are reduced in mast cell-deficient Kit(W-sh) mice. Hepatology 2017, 65, 1991–2004.
- Atiakshin, D.; Buchwalow, I.; Tiemann, M. Mast cells and collagen fibrillogenesis. Histochem. Cell Biol. 2020, 154, 21–40.
- Moyer, K.E.; Saggers, G.C.; Ehrlich, H.P. Mast cells promote fibroblast populated collagen lattice contraction through gap junction intercellular communication. Wound Repair Regen. 2004, 12, 269–275.
- Chen, H.; Xu, Y.; Yang, G.; Zhang, Q.; Huang, X.; Yu, L.; Dong, X. Mast cell chymase promotes hypertrophic scar fibroblast proliferation and collagen synthesis by activating TGF-beta1/Smads signaling pathway. Exp. Ther. Med. 2017, 14, 4438–4442.
- Wolak, M.; Bojanowska, E.; Staszewska, T.; Piera, L.; Szymanski, J.; Drobnik, J. Histamine augments collagen content via H1 receptor stimulation in cultures of myofibroblasts taken from wound granulation tissue. Mol. Cell. Biochem. 2021, 476, 1083–1092.
- Kennedy, L.; Meadows, V.; Demieville, J.; Hargrove, L.; Virani, S.; Glaser, S.; Zhou, T.; Rinehart, E.; Jaeger, V.; Kyritsi, K.; et al. Biliary damage and liver fibrosis are ameliorated in a novel mouse model lacking l-histidine decarboxylase/histamine signaling. Lab. Investig. 2020, 100, 837–848.
- Kulke, M.; Geist, N.; Friedrichs, W.; Langel, W. Molecular dynamics simulations on networks of heparin and collagen. Proteins 2017, 85, 1119–1130.
- Roy, A.; Ganesh, G.; Sippola, H.; Bolin, S.; Sawesi, O.; Dagalv, A.; Schlenner, S.M.; Feyerabend, T.; Rodewald, H.R.; Kjellen, L.; et al. Mast cell chymase degrades the alarmins heat shock protein 70, biglycan, HMGB1, and interleukin-33 (IL-33) and limits danger-induced inflammation. J. Biol. Chem. 2014, 289, 237–250.
- Atiakshin, D.; Buchwalow, I.; Horny, P.; Tiemann, M. Protease profile of normal and neoplastic mast cells in the human bone marrow with special emphasis on systemic mastocytosis. Histochem. Cell Biol. 2021, 155, 561–580.
- Atiakshin, D.; Buchwalow, I.; Tiemann, M. Mast cell chymase: Morphofunctional characteristics. Histochem. Cell Biol. 2019, 152, 253–269.
- Atiakshin, D.; Buchwalow, I.; Samoilova, V.; Tiemann, M. Tryptase as a polyfunctional component of mast cells. Histochem. Cell Biol. 2018, 149, 461–477.
- Bayat, M.; Chien, S.; Chehelcheraghi, F. Co- localization of Flt1 and tryptase of mast cells in skin wound of rats with type I diabetes: Initial studies. Acta Histochem. 2021, 123, 151680.
- Younan, G.J.; Heit, Y.I.; Dastouri, P.; Kekhia, H.; Xing, W.; Gurish, M.F.; Orgill, D.P. Mast cells are required in the proliferation and remodeling phases of microdeformational wound therapy. Plast. Reconstr. Surg. 2011, 128, 649e–658e.
- Trautmann, A.; Toksoy, A.; Engelhardt, E.; Gillitzer, R. Mast cell involvement in normal human skin wound healing: Expression of monocyte chemoattractant protein-1 is correlated with recruitment of mast cells which synthesize interleukin-4 in vivo. J. Pathol. 2000, 190, 100–106.
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A review of the contribution of mast cells in wound healing: Involved molecular and cellular mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312.
- Blank, U.; Madera-Salcedo, I.K.; Danelli, L.; Claver, J.; Tiwari, N.; Sanchez-Miranda, E.; Vazquez-Victorio, G.; Ramirez-Valadez, K.A.; Macias-Silva, M.; Gonzalez-Espinosa, C. Vesicular trafficking and signaling for cytokine and chemokine secretion in mast cells. Front. Immunol. 2014, 5, 453.
- Tellechea, A.; Leal, E.C.; Kafanas, A.; Auster, M.E.; Kuchibhotla, S.; Ostrovsky, Y.; Tecilazich, F.; Baltzis, D.; Zheng, Y.; Carvalho, E.; et al. Mast cells regulate wound healing in diabetes. Diabetes 2016, 65, 2006–2019.
- Kashyap, M.; Thornton, A.M.; Norton, S.K.; Barnstein, B.; Macey, M.; Brenzovich, J.; Shevach, E.; Leonard, W.J.; Ryan, J.J. Cutting edge: CD4 T cell-mast cell interactions alter IgE receptor expression and signaling. J. Immunol. 2008, 180, 2039–2043.
- Gorczynski, R.M.; Chen, Z.; Khatri, I.; Yu, K. Graft-infiltrating cells expressing a CD200 transgene prolong allogeneic skin graft survival in association with local increases in Foxp3(+)Treg and mast cells. Transpl. Immunol. 2011, 25, 187–193.
- Gri, G.; Piconese, S.; Frossi, B.; Manfroi, V.; Merluzzi, S.; Tripodo, C.; Viola, A.; Odom, S.; Rivera, J.; Colombo, M.P.; et al. CD4+CD25+ regulatory T cells suppress mast cell degranulation and allergic responses through OX40-OX40L interaction. Immunity 2008, 29, 771–781.
- Ohkura, N.; Kitagawa, Y.; Sakaguchi, S. Development and maintenance of regulatory T cells. Immunity 2013, 38, 414–423.
- Bonnet, B.; Vigneron, J.; Levacher, B.; Vazquez, T.; Pitoiset, F.; Brimaud, F.; Churlaud, G.; Klatzmann, D.; Bellier, B. Low-Dose IL-2 Induces Regulatory T Cell-Mediated Control of Experimental Food Allergy. J. Immunol. 2016, 197, 188–198.
- Shevach, E.M. Application of IL-2 therapy to target T regulatory cell function. Trends. Immunol. 2012, 33, 626–632.
- Morita, H.; Arae, K.; Unno, H.; Miyauchi, K.; Toyama, S.; Nambu, A.; Oboki, K.; Ohno, T.; Motomura, K.; Matsuda, A.; et al. An interleukin-33-mast cell-interleukin-2 axis suppresses papain-induced allergic inflammation by promoting regulatory T cell numbers. Immunity 2015, 43, 175–186.
- Hatzioannou, A.; Banos, A.; Sakelaropoulos, T.; Fedonidis, C.; Vidali, M.S.; Kohne, M.; Handler, K.; Boon, L.; Henriques, A.; Koliaraki, V.; et al. An intrinsic role of IL-33 in Treg cell-mediated tumor immunoevasion. Nat. Immunol. 2020, 21, 75–85.
- Leyva-Castillo, J.M.; Das, M.; Artru, E.; Yoon, J.; Galand, C.; Geha, R.S. Mast cell-derived IL-13 downregulates IL-12 production by skin dendritic cells to inhibit the TH1 cell response to cutaneous antigen exposure. J. Allergy Clin. Immunol. 2021, 147, 2305–2315.

