Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Yoshikazu Nakamura | + 947 word(s) | 947 | 2021-07-06 12:18:41 | | | |
| 2 | Vicky Zhou | Meta information modification | 947 | 2021-07-08 05:16:35 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Nakamura, Y. RBM-007. Encyclopedia. Available online: https://encyclopedia.pub/entry/11805 (accessed on 26 June 2026).
Nakamura Y. RBM-007. Encyclopedia. Available at: https://encyclopedia.pub/entry/11805. Accessed June 26, 2026.
Nakamura, Yoshikazu. "RBM-007" Encyclopedia, https://encyclopedia.pub/entry/11805 (accessed June 26, 2026).
Nakamura, Y. (2021, July 08). RBM-007. In Encyclopedia. https://encyclopedia.pub/entry/11805
Nakamura, Yoshikazu. "RBM-007." Encyclopedia. Web. 08 July, 2021.
Copy Citation
RBM-007 is an anti-FGF2 aptamer composed of 37 nucleotides, whose ribose 2′ positions are modified to resist ribonucleases, in addition to being 5′-PEGylated and 3′-conjugated with an inverted dT to confer an advantageous pharmacokinetic profile. RBM-007 binds strongly and specifically to FGF2 and does not cross-react with other FGF family proteins or heparin-binding proteins, blocking the interaction between human FGF2 and its receptors FGFR1 through FGFR4. The dissociation constant (KD) of the non-PEGylated form of RBM-007 to human FGF2 protein is 2 pM, compared to 5, 7, and 27 pM in rat, mouse, and rabbit protein, respectively, showing the high affinity of RBM-007 for different FGF2s regardless of the species difference.
fibroblast growth factor 2
RNA aptamer
age-related macular degeneration
achondroplasia
lung cancer
cancer pain
1. Introduction
In mammals, fibroblast growth factors (FGF) have 22 known members that exert important functions in regulating cell proliferation, differentiation, and migration [1][2]. Upon binding to tyrosine kinase FGF receptors FGFR1–FGFR4, FGFs activate essential signaling pathways, such as the mitogen-activated protein kinase (MAPK)/ERK and JNK pathways, that are centrally involved in angiogenesis, tissue remodeling, and regeneration, including the repair of neuronal damage, skin wound healing, joint protection, and the control of hypertension. Among the FGFs, human FGF2 is an 18-kDa non-glycosylated polypeptide composed of 146 amino acids [3].
FGF2 generally plays a positive role in bone health; however, several in vitro studies have demonstrated the adverse role of FGF2 in the progression of bone disease [4][5][6][7]. Recent advances in understanding the role of FGF2 in bone formation alternatively posit that the pharmaceutical manipulation of FGF2 signaling may be a promising approach for bone disease therapy. In the literature, a number of anti-FGF2 neutralizing monoclonal antibodies (mAbs) have been reported [8][9][10][11][12], while no anti-FGF2 mAbs have been investigated for the clinical potency.
Focusing on the therapeutic potential in inhibiting FGF2, we developed the inhibitory RNA aptamer, RBM-007 [13]. In the literature, a few aptamers (DNA & RNA-based) against FGF2 (or bFGF) were described in the 1990s, but there appeared no subsequent studies on these aptamers. Aptamers are single-stranded short oligonucleotides selected in vitro from a large random sequence library, and are applicable to therapies because of several pharmaceutical advantages, such as a medium size between small molecules and antibodies, chemical synthesis, production cost, and low antigenicity [14]. Aptamers are built on the foundation of targeted molecules using the systematic evolution of ligands by exponential enrichment (SELEX) [15][16]. The concept relies on the potential of short oligonucleotides to fold, in the presence of a target, unique tertiary structures that bind with high specificity and affinity. We highlight the broad therapeutic potential of RBM-007 and the multifunctionality of FGF2 (Figure 1) in the treatments of wet AMD, achondroplasia, cancer pain and lung cancer, respectively.
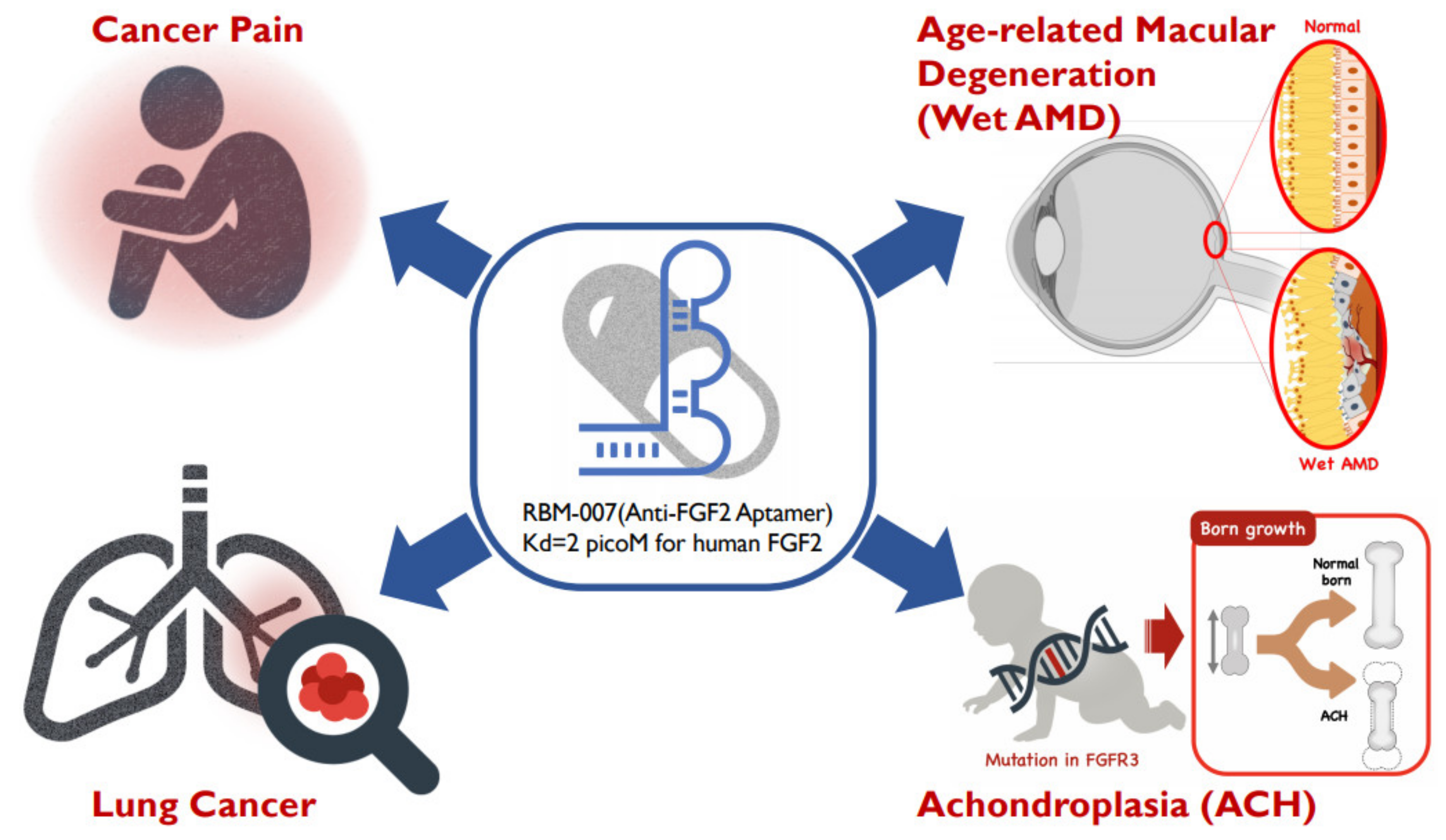 Figure 1. Diverse therapeutic applications of RBM-007 (anti-FGF2 aptamer).
Figure 1. Diverse therapeutic applications of RBM-007 (anti-FGF2 aptamer).2. Facts about RBM-007
Aptamers prefer to interact with positively charged surfaces of the target proteins due to the negatively charged nature of backbone linkages. We have shown previously that a 23-nucleotide (nt) RNA aptamer interacting with the Fc domain of human IgG1 (hFc1), which lacks positive charges on the surface, can achieve strong and specific affinity to hFc1 through multiple non-electrostatic forces, such as hydrogen bonds and hydrophobic interactions [17]. This strong affinity relies on the shape complementarity between aptamers and targets [18]. Therefore, the aptamer technology is applicable not only for positively charged molecules, but also for relatively neutral molecules.
In our laboratory, we have raised therapeutic RNA aptamers to a variety of human proteins and forwarded some of them to preclinical or clinical studies. Although aptamers share many properties with antibodies, the aptamers exhibited some superior features, such as higher affinity, medium size between small molecules and antibodies, chemical synthesis, production cost, and low antigenicity. This review summarized the properties of one such aptamer, RBM-007, against FGF2, with pre-clinical and clinical development. Reflecting the multifunctional properties of FGF2, excess FGF2 activity is involved in disease progression. Four therapeutic applications have been described here, including the treatment of wet AMD, achondroplasia, bone cancer pain, and lung cancer—all of which show RBM-007 as an expected, overall candidate for the therapeutic treatment of diseases affected by excess FGF2.
Regarding wet AMD, we have shown in vitro and in vivo evidence that excess FGF2 plays a vital role in wet AMD by promoting angiogenesis, and that RBM-007 blocks both choroidal neovascularization and subretinal fibrosis. Moreover, combined treatment of RBM-007 and ranibizumab (Lucentis®) showed a synergistic effect in preventing CNV [19] although the action mechanisms remain to be clarified. This therapeutic array is further supported by the finding that FGF receptor double-conditional knockout (Fgfr1/2) mice showed a marked reduction in CNV accompanied by a decrease in the FGF2 level upon laser injury [20]. Additionally, FGF2 was the only essential ligand in the in vivo models of CNV, showing that FGF2 regulates pathogenic angiogenesis via the STAT3 pathway [21].
The therapeutic applicability of RBM-007 for achondroplasia was first suggested in an ovariectomized (OVX) rat model, a well-established model for osteoporosis; severe disruption of the epiphyseal growth plate occurred in OVX rats and RBM-007 sharply blocked the disruption of the epiphyseal growth plate in a dose-dependent manner [16]. This finding suggests that RBM-007 might ameliorate the epiphyseal growth plate and prevent skeletal dysplasias, including achondroplasia, which is related to excessive FGFR3 activation.
In future, a long-lasting modification of RBM-007 will offer potential for extended therapeutic duration that could be beneficial for wet AMD patients who in “real-world” clinical management receive less frequent intravitreal injections, as well as for ACH young patients who receive subcutaneous injections for the entire medication period from childhood for 12–14 years. Ongoing and further clinical studies along these lines should provide us with a novel therapy for unmet medical needs and other applications, such as to diagnostics or for immune modulation.
References
- Krejci, P.; Prochazkova, J.; Bryja, V.; Kozubik, A.; Wilcox, W.R. Molecular pathology of the fibroblast growth factor family. Hum. Mutat. 2011, 30, 1245–1255.
- Marie, P.J.; Miraoui, H.; Severe, N. FGF/FGFR signaling in bone formation: Progress and perspectives. Growth Factors 2012, 30, 117–123.
- Okada-Ban, M.; Thiery, J.P.; Jouanneau, J. Fibroblast growth factor-2. Int. J. Biochem. Cell Biol. 2000, 32, 263–267.
- Eda, H.; Aoki, K.; Marumo, K.; Fujii, K.; Ohkawa, K. FGF-2 signaling induces downregulation of TAZ protein in osteoblastic MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 2008, 366, 471–475.
- Kawaguchi, H.; Nakamura, K.; Tabata, Y.; Ikada, Y.; Aoyama, I.; Anzai, J.; Nakamura, T.; Hiyama, Y.; Tamura, M. Acceleration of fracture healing in nonhuman primates by fibroblast growth factor-2. J. Clin. Endocrinol. Metab. 2001, 86, 875–880.
- Nakagawa, N.; Yasuda, H.; Yano, K.; Mochizuki, S.; Kobayashi, N.; Fujimoto, H.; Shima, N.; Morinaga, T.; Chikazu, D.; Kawaguchi, H.; et al. Basic fibroblast growth factor induces osteoclast formation by reciprocally regulating the production of osteoclast differentiation factor and osteoclastogenesis inhibitory factor in mouse osteoblastic cells. Biochem. Biophys. Res. Commun. 1999, 265, 158–163.
- Kawaguchi, H.; Chikazu, D.; Nakamura, K.; Kumegawa, M.; Hakeda, Y. Direct and indirect actions of fibroblast growth factor 2 on osteoclastic bone resorption in cultures. J. Bone Miner. Res. 2000, 15, 466–473.
- Matsuzaki, K.; Yoshitake, Y.; Matuo, Y.; Sasaki, H.; Nishikawa, K. Monoclonal antibodies against heparin-binding growth factor II/basic fibroblast growth factor that block its biological activity: Invalidity of the antibodies for tumor angiogenesis. Proc. Natl. Acad. Sci. USA 1989, 86, 9911–9915.
- Rege, A.A.; Bjercke, R.J.; Erichsen, D.; Owens, R.; Stephan, C.C.; Brock, T.A. Development of novel monoclonal antibodies for the analysis of functional sites in FGF-2. Growth Factors 1999, 16, 161–169.
- Kuhn, H.; Kopff, C.; Konrad, J.; Riedel, A.; Gessner, C.; Wirtz, H. Influence of basic fibroblast growth factor on the proliferation of non-small cell lung cancer cell lines. Lung Cancer 2004, 44, 167–174.
- Hori, A.; Sasada, R.; Matsutani, E.; Naito, K.; Sakura, Y.; Fujita, T.; Kozai, Y. Suppression of solid tumor growth by immunoneutralizing monoclonal antibody against human basic fibroblast growth factor. Cancer Res. 1991, 51, 6180–6184.
- Wang, L.; Park, H.; Chhim, S.; Ding, Y.; Jiang, W.; Queen, C.; Kim, K.J. A novel monoclonal antibody to fibroblast growth factor 2 effectively inhibits growth of hepatocellular carcinoma xenografts. Mol. Cancer Ther. 2012, 11, 864–872.
- Jin, L.; Nonaka, Y.; Miyakawa, S.; Fujiwara, M.; Nakamura, Y. Dual therapeutic action of a neutralizing anti-FGF2 aptamer in bone diseases and bone cancer pain. Mol. Ther. 2016, 24, 1974–1986.
- Keefe, A.D.; Pai, S.; Ellington, A. Aptamers as therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550.
- Ellington, A.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822.
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510.
- Nomura, Y.; Sugiyama, S.; Sakamoto, T.; Miyakawa, S.; Adachi, H.; Takano, K.; Murakami, S.; Inoue, T.; Mori, Y.; Nakamura, Y.; et al. Conformational plasticity of RNA for target recognition as revealed by the 2.15 Å crystal structure of a human IgG-aptamer complex. Nucl. Acids Res. 2010, 38, 7822–7829.
- Nakamura, Y. Aptamers as therapeutic middle molecules. Biochimie 2018, 145, 22–33.
- Matsuda, Y.; Nonaka, Y.; Futakawa, S.; Imai, H.; Akita, K.; Nishihata, T.; Fujiwara, M.; Ali, Y.; Bhisitkul, R.B.; Nakamura, Y. Anti-angiogenic and anti-scarring dual action of an anti-fibroblast growth factor 2 aptamer in animal models of retinal disease. Mol. Ther. Nucl. Acids 2019, 17, 819–828.
- Oladipupo, S.S.; Smith, C.; Santeford, A.; Park, C.; Sene, A.; Wiley, L.A.; Osei-Owusu, P.; Hsu, J.; Zapata, N.; Liu, F.; et al. Endothelial cell FGF signaling is required for injury response but not for vascular homeostasis. Proc. Natl. Acad. Sci. USA 2014, 111, 13379–13384.
- Dong, Z.; Santeford, A.; Ban, N.; Lee, T.J.; Smith, C.; Smith, C.; Ornitz, D.M.; Apte, R.S. FGF2-induced STAT3 activation regulates pathologic neovascularization. Exp. Eye Res. 2019, 187, 107775.
More
Information
Subjects:
Pharmacology & Pharmacy
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.5K
Revisions:
2 times
(View History)
Update Date:
08 Jul 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No