 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Abenaya Muralidharan | + 1599 word(s) | 1599 | 2021-06-04 10:04:02 | | | |
| 2 | Enzi Gong | Meta information modification | 1599 | 2021-07-05 05:42:48 | | |
Video Upload Options
Arboviruses are known to cause large-scale epidemics in many parts of the world. These arthropod-borne viruses are a large group consisting of viruses from a wide range of families. The ability of their vector to enhance viral pathogenesis and transmission makes the development of treatments against these viruses challenging. Neutrophils are generally the first leukocytes to be recruited to a site of infection, playing a major role in regulating inflammation and, as a result, viral replication and dissemination. However, the underlying mechanisms through which neutrophils control the progression of inflammation and disease remain to be fully understood. In this review, we highlight the major findings from recent years regarding the role of neutrophils during arboviral infections. We discuss the complex nature of neutrophils in mediating not only protection, but also augmenting disease pathology. Better understanding of neutrophil pathways involved in effective protection against arboviral infections can help identify potential targets for therapeutics.
1. Introduction
2. Mosquitoes
3. Alphaviruses
4. Conclusions
| Arbovirus | Beneficial Roles of Neutrophils | Detrimental Roles of Neutrophils | Replication in Neutrophils |
|---|---|---|---|
|
Zika virus (ZIKV) |
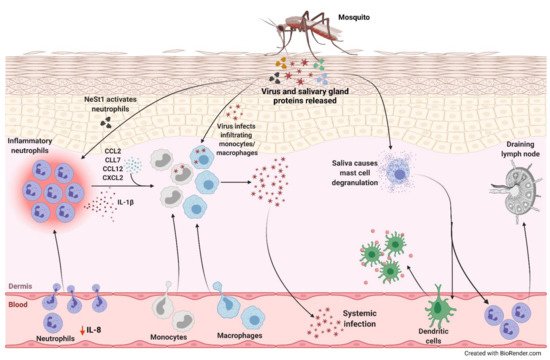
• In AG129 IFN-α/β receptor knockout mice, ZIKV induced neutrophil infiltration, which inversely correlated with virus-induced paresis protecting mice from motor deficits [28] |
• Neutrophil-stimulating factor 1 (NeSt1) in mosquito saliva activated neutrophils, inducing IL-1β, CXCL2 and CCL2 expression, which augmented early viral infection in mice [29]
|
|
|
Dengue virus (DENV) |
Clinical studies:
|
||
|
West Nile virus (WNV) |
• Neutrophil depletion after infection increased viremia and death rate in mice [5] |
• Neutrophil depletion before infection reduced viral burden and enhanced survival in mice [5] |
Yes [5] |
|
Semliki Forest virus (SFV) |
Mice: • At later stages of infection, neutrophils were required to resolve the infection and decrease mortality [34] |
Mice:
|
|
|
Ross River virus (RRV) |
• Deletion of Arg1 in macrophages and neutrophils enhanced viral clearance and improved skeletal muscle tissue pathology in late stages of infection in mice, with no effect in the acute phase of infection [35] |
||
|
Chikungunya virus (CHIKV) |
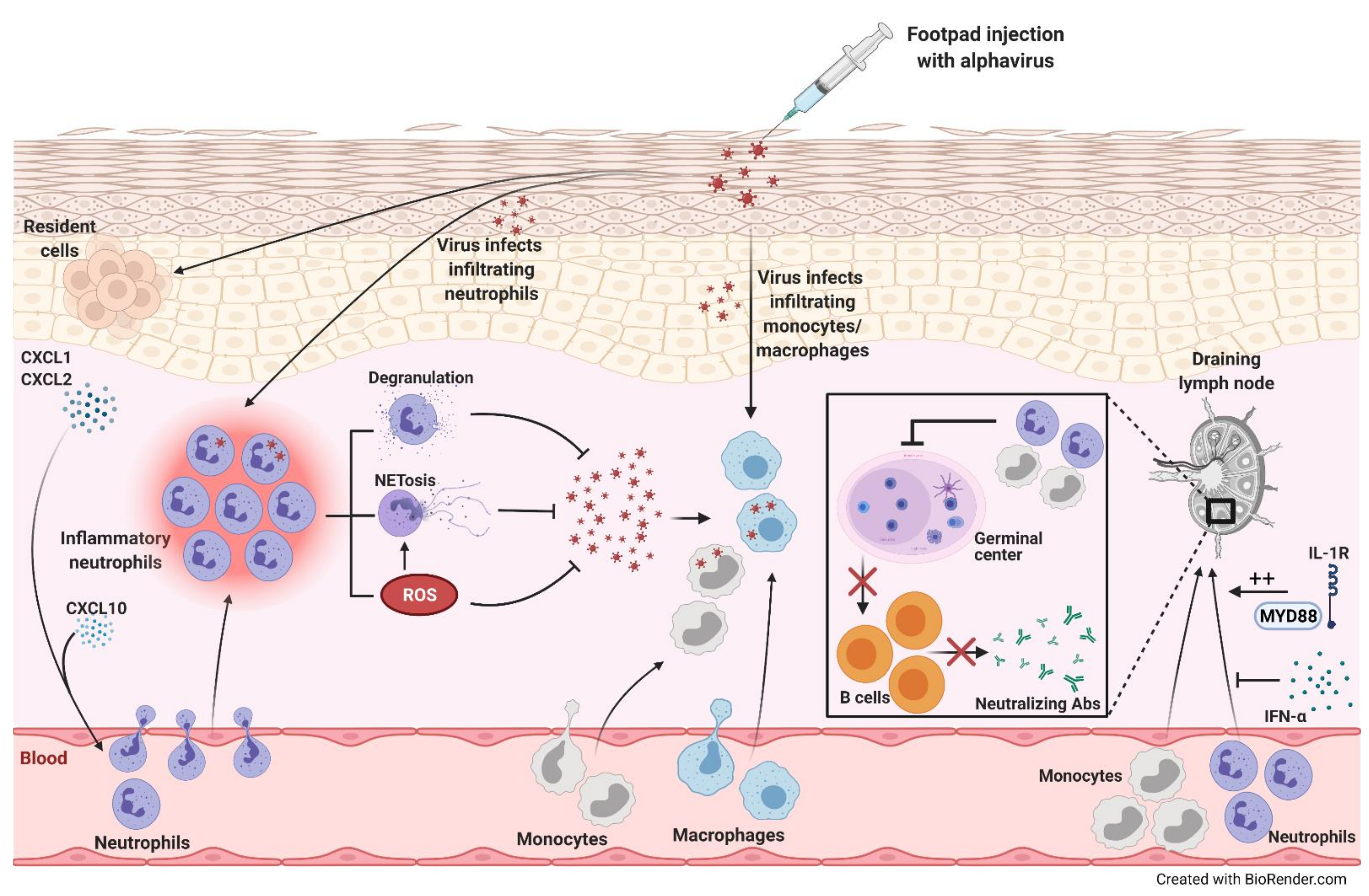
Zebrafish: • Neutrophils served as a major source of type I interferon for eliminating the virus and alleviating disease [36] Mice: |
Mice:
|
|
|
O’nyong nyong virus (ONNV) |
• CXCL10−/− mice had reduced influx of macrophages and neutrophils, which was associated with decreased viral loads and foot swelling [41] |
Yes [41] |
References
- Leliefeld, P.H.C.; Koenderman, L.; Pillay, J. How neutrophils shape adaptive immune responses. Front. Immunol. 2015, 6, 471.
- Geerdink, R.J.; Pillay, J.; Meyaard, L.; Bont, L. Neutrophils in respiratory syncytial virus infection: A target for asthma prevention. J. Allergy Clin. Immunol. 2015, 136, 838–847.
- Li, Y.; Wang, W.; Yang, F.; Xu, Y.; Feng, C.; Zhao, Y. The regulatory roles of neutrophils in adaptive immunity. Cell Commun. Signal. 2019, 17, 147.
- Zhao, Y.; Lu, M.; Lau, L.T.; Lu, J.; Gao, Z.; Liu, J.; Yu, A.C.H.; Cao, Q.; Ye, J.; McNutt, M.A.; et al. Neutrophils may be a vehicle for viral replication and dissemination in human h5n1 avian influenza. Clin. Infect. Dis. 2008, 47, 1575–1578.
- Bai, F.; Kong, K.F.; Dai, J.; Qian, F.; Zhang, L.; Brown, C.R.; Fikrig, E.; Montgomery, R.R. A paradoxical role for neutrophils in the pathogenesis of West Nile virus. J. Infect. Dis. 2010, 202, 1804–1812.
- Elliott, R.M. Orthobunyaviruses: Recent genetic and structural insights. Nat. Rev. Microbiol. 2014, 12, 673–685.
- Gould, E.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509.
- Powers, A.M.; Brault, A.C.; Shirako, Y.; Strauss, E.G.; Kang, W.; Strauss, J.H.; Weaver, S.C. Evolutionary relationships and systematics of the alphaviruses. J. Virol. 2001, 75, 10118–10131.
- Durbin, A.P.; Mayer, S.V.; Rossi, S.L.; Amaya-Larios, I.Y.; Ramos-Castaneda, J.; Eong Ooi, E.; Jane Cardosa, M.; Munoz-Jordan, J.L.; Tesh, R.B.; Messer, W.B.; et al. Emergence potential of sylvatic dengue virus type 4 in the urban transmission cycle is restrained by vaccination and homotypic immunity. Virology 2013, 439, 34–41.
- Huang, Y.J.S.; Higgs, S.; Horne, K.M.E.; Vanlandingham, D.L. Flavivirus-Mosquito interactions. Viruses 2014, 6, 4703–4730.
- Lambrechts, L.; Scott, T.W.; Gubler, D.J. Consequences of the expanding global distribution of aedes albopictus for dengue virus transmission. PLoS Negl. Trop. Dis. 2010, 4, e646.
- Miller, M.J.; Loaiza, J.R. Geographic expansion of the invasive mosquito aedes albopictus across Panama—Implications for control of dengue and chikungunya viruses. Plos Negl. Trop. Dis. 2015, 9.
- Guagliardo, S.A.; Barboza, J.L.; Morrison, A.C.; Astete, H.; Vazquez-Prokopec, G.; Kitron, U. Patterns of geographic expansion of aedes aegypti in the Peruvian Amazon. PLoS Negl. Trop. Dis. 2014, 8.
- Díaz-Nieto, L.M.; Maciá, A.; Perotti, M.A.; Berón, C.M. Geographical limits of the southeastern distribution of aedes aegypti (diptera, culicidae) in Argentina. Plos Negl. Trop. Dis. 2013, 7.
- MacKenzie, J.S.; Williams, D.T. The zoonotic flaviviruses of southern, South-Eastern and eastern Asia, and Australasia: The potential for emergent viruses. Zoonoses Public Health 2009, 56, 338–356.
- Gabriel, C.; Her, Z.; Ng, L.F.P. Neutrophils: Neglected players in viral diseases. DNA Cell Biol. 2013, 32, 665–675.
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J. Virol. 2012, 86, 7637–7649.
- Edwards, J.F.; Higgs, S.; Beaty, B.J. Mosquito feeding-induced enhancement of cache valley virus (Bunyaviridae) infection in mice. J. Med. Entomol. 1998, 35, 261–265.
- Limesand, K.H.; Higgs, S.; Pearson, L.D.; Beaty, B.J. Potentiation of vesicular stomatitis New Jersey virus infection in mice by mosquito saliva. Parasite Immunol. 2000, 22, 461–467.
- Schneider, B.S.; Soong, L.; Girard, Y.A.; Campbell, G.; Mason, P.; Higgs, S. Potentiation of West Nile encephalitis by mosquito feeding. Viral Immunol. 2006, 19, 74–82.
- Dessens, J.T.; Nuttall, P.A. Mx1-based resistance to thogoto virus in A2G mice is bypassed in tick-mediated virus delivery. J. Virol. 1998, 72, 8362–8364.
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507.
- Weaver, S.C.; Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 2015, 372, 1231–1239.
- WHO. A Global Brief on Vector-Borne Diseases; WHO: Geneva, Switzerland, 2014.
- WHO. WHO Factsheet: Vector-Borne Diseases, Factsheet #387; WHO: Geneva, Switzerland, 2014.
- Baxter, V.K.; Heise, M.T. Genetic control of alphavirus pathogenesis. Mamm. Genome 2018, 29, 408–424.
- Hollidge, B.S.; González-Scarano, F.; Soldan, S.S. Arboviral encephalitides: Transmission, emergence, and pathogenesis. J. Neuroimmune Pharmacol. 2010, 5, 428–442.
- Zukor, K.; Wang, H.; Siddharthan, V.; Julander, J.G.; Morrey, J.D. Zika virus-induced acute myelitis and motor deficits in adult interferon αβ/γ receptor knockout mice. J. Neurovirol. 2018, 24, 273–290.
- Hastings, A.K.; Uraki, R.; Gaitsch, H.; Dhaliwal, K.; Stanley, S.; Sproch, H.; Williamson, E.; MacNeil, T.; Marin-Lopez, A.; Hwang, J.; et al. Aedes aegypti NeSt1 protein enhances zika virus pathogenesis by activating neutrophils. J. Virol. 2019, 93.
- Hoang, L.T.; Lynn, D.J.; Henn, M.; Birren, B.W.; Lennon, N.J.; Le, P.T.; Duong, K.T.H.; Nguyen, T.T.H.; Mai, L.N.; Farrar, J.J.; et al. The early whole-blood transcriptional signature of dengue virus and features associated with progression to dengue shock syndrome in vietnamese children and young adults. J. Virol. 2010, 84, 12982–12994.
- Kunder, M.; Lakshmaiah, V.; Moideen Kutty, A.V. Plasma neutrophil elastase, α1-antitrypsin, α2-macroglobulin and neutrophil elastase–α1-antitrypsin complex levels in patients with dengue fever. Indian J. Clin. Biochem. 2018, 33, 218–221.
- Opasawatchai, A.; Amornsupawat, P.; Jiravejchakul, N.; Chan-in, W.; Spoerk, N.J.; Manopwisedjaroen, K.; Singhasivanon, P.; Yingtaweesak, T.; Suraamornkul, S.; Mongkolsapaya, J.; et al. Neutrophil activation and early features of NET formation are associated with dengue virus infection in human. Front. Immunol. 2019, 9, 3007.
- Sung, P.S.; Huang, T.F.; Hsieh, S.L. Extracellular vesicles from CLEC2-activated platelets enhance dengue virus-induced lethality via CLEC5A/TLR2. Nat. Commun. 2019, 10, 1–13.
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host inflammatory response to mosquito bites enhances the severity of arbovirus infection. Immunity 2016, 44, 1455–1469.
- Stoermer, K.A.; Burrack, A.; Oko, L.; Montgomery, S.A.; Borst, L.B.; Gill, R.G.; Morrison, T.E. Genetic ablation of arginase 1 in macrophages and neutrophils enhances clearance of an arthritogenic alphavirus. J. Immunol. 2012, 189, 4047–4059.
- Palha, N.; Guivel-Benhassine, F.; Briolat, V.; Lutfalla, G.; Sourisseau, M.; Ellett, F.; Wang, C.-H.; Lieschke, G.J.; Herbomel, P.; Schwartz, O.; et al. Real-time whole-body visualization of chikungunya virus infection and host interferon response in zebrafish. PLoS Pathog. 2013, 9, e1003619.
- Hiroki, C.H.; Toller-Kawahisa, J.E.; Fumagalli, M.J.; Colon, D.F.; Figueiredo, L.T.M.; Fonseca, B.A.L.D.; Franca, R.F.O.; Cunha, F.Q. Neutrophil extracellular traps effectively control acute chikungunya virus infection. Front. Immunol. 2020, 10.
- Poo, Y.S.; Nakaya, H.; Gardner, J.; Larcher, T.; Schroder, W.A.; Le, T.T.; Major, L.D.; Suhrbier, A. CCR2 deficiency promotes exacerbated chronic erosive neutrophil-dominated chikungunya virus arthritis. J. Virol. 2014, 88, 6862–6872.
- McCarthy, M.K.; Reynoso, G.V.; Winkler, E.S.; Mack, M.; Diamond, M.S.; Hickman, H.D.; Morrison, T.E. MyD88-dependent influx of monocytes and neutrophils impairs lymph node B cell responses to chikungunya virus infection via Irf5, Nos2 and Nox2. PLoS Pathog. 2020, 16, e1008292.
- Cook, L.E.; Locke, M.C.; Young, A.R.; Monte, K.; Hedberg, M.L.; Shimak, R.M.; Sheehan, K.C.F.; Veis, D.J.; Diamond, M.S.; Lenschow, D.J. Distinct roles of interferon alpha and beta in controlling chikungunya virus replication and modulating neutrophil-mediated inflammation. J. Virol. 2019, 94.
- Lin, T.; Geng, T.; Harrison, A.G.; Yang, D.; Vella, A.T.; Fikrig, E.; Wang, P. CXCL10 signaling contributes to the pathogenesis of arthritogenic alphaviruses. Viruses 2020, 12, 1252.

