 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Stanislawa Bazan-Socha | + 1916 word(s) | 1916 | 2021-06-18 11:27:54 | | | |
| 2 | Rita Xu | -29 word(s) | 1887 | 2021-07-01 09:36:06 | | | | |
| 3 | Catherine Yang | Meta information modification | 1887 | 2021-09-30 03:37:11 | | |
Video Upload Options
Increased airway wall thickness and remodeling of bronchial mucosa are characteristic of asthma and may arise from altered integrin signaling on airway cells. Here, we analyzed the expression of β1-subfamily integrins on blood and airway cells (flow cytometry), inflammatory biomarkers in serum and bronchoalveolar lavage, reticular basement membrane (RBM) thickness and collagen deposits in the mucosa (histology), and airway geometry (CT-imaging) in 92 asthma patients (persistent airflow limitation subtype: n=47) and 36 controls. Persistent airflow limitation was associated with type-2 inflammation, elevated soluble α2 integrin chain, and changes in the bronchial wall geometry. Both subtypes of asthma showed thicker RBM than control, but collagen deposition and epithelial α1 and α2 integrins staining were similar. Type-I collagen accumulation and RBM thickness were inversely related to the epithelial expression of the α2 integrin chain. Expression of α2β1 integrin on T-cells and eosinophils was not altered in asthma. Collagen I deposits were, however, more abundant in patients with lower α2β1 integrin on blood and airway CD8+ T-cells. Thicker airway walls in CT were associated with lower α2 integrin chain on blood CD4+ T-cells and airway eosinophils. Our data suggest that α2β1 integrin on inflammatory and epithelial cells may protect against airway remodeling advancement in asthma.
1. Introduction
2. Clinical Characteristics and Airway Inflammatory Signature in Asthma Patients with Persistent Airflow Limitation
| Non-Persistent Airflow Limitation n = 45 |
Persistent Airflow Limitation n = 47 |
Control n = 36 |
p-Value Non-Persistent vs. Persistent Limitation |
p-Value Non-Persistent Limitation vs. Control |
p-Value Persistent Limitation vs. Control |
|
|---|---|---|---|---|---|---|
| Demographic variables | ||||||
| Age, years | 52 (41–59) | 58 (52–65) | 55 (45–65) | 0.004 | 0.07 | 0.27 |
| Male gender, n (%) | 10 (22) | 16 (34) | 5 (14) | 0.15 | 0.5 | 0.07 |
| Body mass index, kg/m2 | 27.8 (24.8–30.8) | 26.4 (23.5–31.6) | 27.3 (23.0–27.9) | 0.53 | 0.13 | 0.95 |
| Smoking history | ||||||
| Past smoking, n (%) | 13 (29) | 15 (32) | 12 (33) | 0.93 | 0.85 | 0.92 |
| Pack-years of smoking | 0 (0–7) | 0 (0–8) | 0 (0–4) | 0.85 | 0.84 | 0.9 |
| Comorbidities | ||||||
| Atopy, n (%) | 27 (60) | 23 (49) | 6 (17) | 0.39 | 0.0002 | 0.005 |
| GERD, n (%) | 16 (36) | 22 (47) | 23 (64) | 0.38 | 0.02 | 0.19 |
| Arterial hypertension, n (%) | 18 (40) | 28 (60) | 15 (42) | 0.09 | 0.94 | 0.16 |
| Diabetes mellitus, n (%) | 6 (13) | 12 (26) | 3 (8) | 0.23 | 0.72 | 0.08 |
| Hypercholesterolemia, n (%) | 9 (20) | 16 (34) | 6 (17) | 0.2 | 0.92 | 0.13 |
| Coronary heart disease, n (%) | 2 (4) | 5 (11) | 2 (6) | 0.47 | 0.77 | 0.67 |
| Asthma-related variables | ||||||
| Asthma duration, years | 11.5 (5–19.5) | 10 (7–20) | 0.86 | |||
| Asthma severity (GINA): persistent mild, n (%) persistent moderate, n (%) persistent severe, n (%) |
8 (18) 22 (49) 15 (33) |
7 (15) 15 (32) 25 (53) |
0.14 |
|||
| Asthma treatment: Inhaled corticosteroids, n (%) Long-acting β2-agonists, n (%) Montelukast, n (%) Theophylline, n (%) Oral corticosteroids, n (%) |
45 (100) 31 (69) 9 (20) 4 (9) 8 (18) |
47 (100) 42 (89) 4 (9) 10 (21) 15 (32) |
0.17 |
|||
| Spirometry results | ||||||
| FEV1 before bronchodilator, L | 2.79 ± 0.76 | 1.79 ± 0.8 | 2.71 ± 0.75 | <0.001 | 0.65 | <0.001 |
| FEV1 before bronchodilator, % of the predicted value | 100.3 (89.5–111.1) | 66.7 (54.1–80.6) | 110.9 (106.8–114.7) | <0.001 | <0.001 | <0.001 |
| FEV1 after bronchodilator, L | 2.92 ± 0.73 | 2.07 ± 0.95 | 2.84 ± 0.79 | <0.001 | 0.66 | <0.001 |
| FEV1 after bronchodilator, % of the predicted value | 103.8 (96.4–116.5) | 79.2 (62.8–87.2) | 116 (112.1–122.3) | <0.001 | <0.001 | <0.001 |
| FEV1/VC (before bronchodilator) | 73.3 (67.8–78.18) | 59.1 (51.7–63.8) | 74.84 (73.23–78.38) | <0.001 | 0.16 | <0.001 |
| FEV1/VC (after bronchodilator) | 76.99 (73.05–81.88) | 65.4 (54.5–68.6) | 79.33 (77.25–80.38) | <0.001 | 0.25 | <0.001 |
| Reference Range | Non-Persistent Airflow Limitation n = 45 |
Persistent Airflow Limitation n = 47 |
Control n = 36 |
p-Value Non-Persistent vs. Persistent Limitation |
p-Value Non-Persistent Limitation vs. Control |
p-Value Persistent Limitation vs. Control |
|
|---|---|---|---|---|---|---|---|
| Basic laboratory tests | |||||||
| Red blood cells, 106/μL | 4–5 | 4.65 ± 0.4 | 4.7 ± 0.5 | 4.48 ± 0.4 | 0.68 | 0.048 | 0.03 |
| White blood cells, 103/μL | 4–10 | 6.26 (5.43–7.33) | 7.44 (6.39–9.25) | 5.44 (5.16–7.08) | <0.001 | 0.07 | <0.001 |
| Neutrophils, 103/μL | 1.8–7.7 | 3.1 (2.7–4.1) | 3.7 (2.9–4.8) | 3.3 (2.9–3.6) | 0.049 | 0.76 | 0.04 |
| Lymphocytes, 103/μL | 1–4.5 | 1.94 (1.58–2.43) | 2.2 (1.58–2.61) | 1.65 (1.44–2.08) | 0.5 | 0.03 | 0.03 |
| Monocytes, 103/μL | 0.1–0.8 | 0.57 (0.49–0.74) | 0.71 (0.53–0.9) | 0.49 (0.41–0.62) | 0.02 | 0.006 | <0.001 |
| Blood platelets, 103/μL | 140–400 | 223 (193–247) | 225 (191–265) | 228 (189–246) | 0.78 | 0.98 | 0.85 |
| Asthma and inflammatory biomarkers (blood) | |||||||
| Eosinophilia/μL | 40–450 | 230 (130–310) | 400 (180–680) | 110 (70–170) | 0.009 | <0.001 | <0.001 |
| Immunoglobulin E, IU/mL | 0–100 | 90 (26–400) | 88 (43–511) | 23 (18–48) | 0.6 | <0.001 | <0.001 |
| C-reactive protein, mg/L | 0–5 | 1.64 (0.53–8) | 4.53 (0.58–9.38) | 1.78 (0.89–2.29) | 0.39 | 0.28 | 0.008 |
| Fibrinogen, g/L | 1.8–3.5 | 3.1 (2.8–3.5) | 3.5 (3.2–4.2) | 2.9 (2.3–3.7) | 0.03 | 0.11 | 0.002 |
| Periostin, ng/mL | 0.29–0.61 § | 0.28 (0.24–0.33) | 0.38 (0.31–0.51) | 0.37 (0.36–0.45) | 0.01 | 0.001 | 0.85 |
| Interleukin 6, pg/mL | 0.005–1.432 § | 0.72 (0.43–1.19) | 1.09 (0.47–2.38) | 0.57 (0.43–0.97) | 0.14 | 0.29 | 0.03 |
| Interleukin 10, pg/mL | 0.163–1.022 § | 0.6 (0.22–1.06) | 0.55 (0.35–0.89) | 0.43 (0.2–0.76) | 0.95 | 0.17 | 0.1 |
| Interleukin 12 (p70), pg/mL | 0.005–2.618 § | 0.005 (0.005–1.2) | 0.005 (0.005–1.25) | 0.005 (0.005–0.33) | 0.7 | 0.13 | 0.26 |
| ADAM-33, ng/mL | 0.083–2.257 § | 0.73 (0.2–1.29) | 1.32 (0.33–2.37) | 0.41 (0.13–1.5) | 0.01 | 0.65 | 0.007 |
| Circulating integrin subunits | |||||||
| α1 integrin, ng/mL | 6.45–103.67 § | 17.32 (6.88–52.4) | 32.7 (14.7–55.7) | 24.1 (8.90–76.5) | 0.14 | 0.21 | 0.83 |
| α2 integrin, ng/mL | 7.79–36.19 § | 15.5 (9.7–25.5) | 22.9 (15–39) | 20.5 (11.7–26.5) | 0.03 | 0.25 | 0.21 |
| Asthma and inflammatory biomarkers (bronchoalveolar lavage fluid) | |||||||
| Periostin, ng/mL | 0.1–1.15 § | 0.86 (0.8–0.99) | 0.81 (0.72–0.95) | 0.8 (0.51–0.88) | 0.34 | 0.17 | 0.49 |
| Eosinophils, % | 0–1 # | 0.5 (0–1) | 1 (0.1–3) | 0.1 (0–1) | 0.02 | 0.62 | 0.006 |
3. Asthma Is Characterized by Decreased Expression of α4 and β1 on Circulating Inflammatory Cells and Increased Expression of α1 Integrin Chain
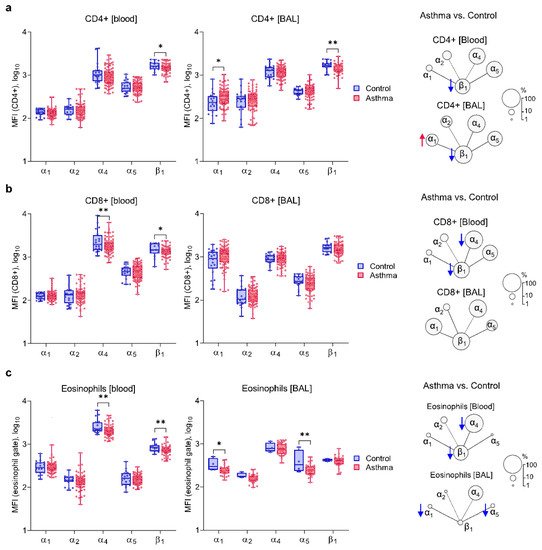
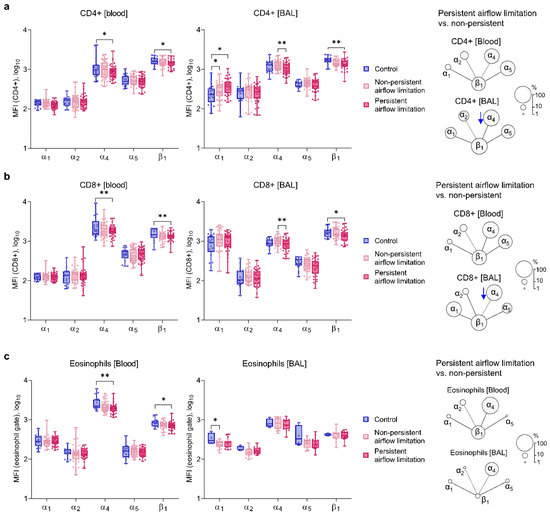
4. Similar Expressions of α1 and α2 Integrin Chains on Blood and BAL Inflammatory Cells of Both Asthma Subsets
References
- Busse, W.W.; Lemanske, R.F. Asthma. N. Engl. J. Med. 2001, 344, 350–362.
- Trejo Bittar, H.E.; Yousem, S.A.; Wenzel, S.E. Pathobiology of Severe Asthma. Annu. Rev. Pathol. Mech. Dis. 2015, 10, 511–545.
- Ito, J.T.; Lourenço, J.D.; Righetti, R.F.; Tibério, I.F.L.C.; Prado, C.M.; Lopes, F.D.T.Q.S. Extracellular Matrix Component Remodeling in Respiratory Diseases: What Has Been Found in Clinical and Experimental Studies? Cells 2019, 8, 342.
- Mostaço-Guidolin, L.B.; Osei, E.T.; Ullah, J.; Hajimohammadi, S.; Fouadi, M.; Li, X.; Li, V.; Shaheen, F.; Yang, C.X.; Chu, F.; et al. Defective Fibrillar Collagen Organization by Fibroblasts Contributes to Airway Remodeling in Asthma. Am. J. Respir. Crit. Care Med. 2019, 200, 431–443.
- Halfter, W.; Oertle, P.; Monnier, C.A.; Camenzind, L.; Reyes-Lua, M.; Hu, H.; Candiello, J.; Labilloy, A.; Balasubramani, M.; Henrich, P.B.; et al. New Concepts in Basement Membrane Biology. FEBS J. 2015, 282, 4466–4479.
- Hirota, N.; Martin, J.G. Mechanisms of Airway Remodeling. Chest 2013, 144, 1026–1032.
- Fehrenbach, H.; Wagner, C.; Wegmann, M. Airway Remodeling in Asthma: What Really Matters. In Cell and Tissue Research; Springer: Cham, Switzerland, 2017; pp. 551–569.
- James, A.L.; Wenzel, S. Clinical Relevance of Airway Remodelling in Airway Diseases. Eur. Respir. J. 2007, 30, 134–155.
- James, A.L.; Bai, T.R.; Mauad, T.; Abramson, M.J.; Dolhnikoff, M.; McKay, K.O.; Maxwell, P.S.; Elliot, J.G.; Green, F.H. Airway Smooth Muscle Thickness in Asthma Is Related to Severity but Not Duration of Asthma. Eur. Respir. J. 2009, 34, 1040–1045.
- Grainge, C.L.; Lau, L.C.K.; Ward, J.A.; Dulay, V.; Lahiff, G.; Wilson, S.; Holgate, S.; Davies, D.E.; Howarth, P.H. Effect of Bronchoconstriction on Airway Remodeling in Asthma. N. Engl. J. Med. 2011, 364, 2006–2015.
- Bazan-Socha, S.; Buregwa-Czuma, S.; Jakiela, B.; Zareba, L.; Zawlik, I.; Myszka, A.; Soja, J.; Okon, K.; Zarychta, J.; Kozlik, P.; et al. Reticular Basement Membrane Thickness Is Associated with Growth-and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients. Int. J. Mol. Sci. 2021, 22, 998.
- Kozlik, P.; Zuk, J.; Bartyzel, S.; Zarychta, J.; Okon, K.; Zareba, L.; Bazan, J.G.; Kosalka, J.; Soja, J.; Musial, J.; et al. The Relationship of Airway Structural Changes to Blood and Bronchoalveolar Lavage Biomarkers, and Lung Function Abnormalities in Asthma. Clin. Exp. Allergy 2020, 50.
- Choi, S.; Hoffman, E.A.; Wenzel, S.E.; Castro, M.; Fain, S.; Jarjour, N.; Schiebler, M.L.; Chen, K.; Lin, C.-L.; National Heart, Lung and Blood Institute’s Severe Asthma Research Program. Quantitative Computed Tomographic Imaging-Based Clustering Differentiates Asthmatic Subgroups with Distinctive Clinical Phenotypes. J. Allergy Clin. Immunol. 2017, 140, 690–700.e8.
- Bazan-Socha, S.; Bukiej, A.; Marcinkiewicz, C.; Musial, J. Integrins in Pulmonary Inflammatory Diseases. Curr. Pharm. Des. 2005, 11, 893–901.
- Zeltz, C.; Gullberg, D. The Integrin-Collagen Connection—A Glue for Tissue Repair? J. Cell Sci. 2016, 129, 653–664.
- Bertoni, A.; Alabiso, O.; Galetto, A.; Baldanzi, G. Integrins in T Cell Physiology. Int. J. Mol. Sci. 2018, 19, 485.
- Schuliga, M.; Harris, T.; Stewart, A.G. Plasminogen Activation by Airway Smooth Muscle Is Regulated by Type I Collagen. Am. J. Respir. Cell Mol. Biol. 2011, 44, 831–839.
- Xia, H.; Seeman, J.; Hong, J.; Hergert, P.; Bodem, V.; Jessurun, J.; Smith, K.; Nho, R.; Kahm, J.; Gaillard, P.; et al. Low α2β1 Integrin Function Enhances the Proliferation of Fibroblasts from Patients with Idiopathic Pulmonary Fibrosis by Activation of the β-Catenin Pathway. Am. J. Pathol. 2012, 181, 222–233.
- Boisvert, M.; Chetoui, N.; Gendron, S.; Aoudjit, F. Alpha2beta1 Integrin Is the Major Collagen-Binding Integrin Expressed on Human Th17 Cells. Eur. J. Immunol. 2010, 40, 2710–2719.
- Rao, W.H.; Hales, J.M.; Camp, R.D.R. Potent Costimulation of Effector T Lymphocytes by Human Collagen Type I. J. Immunol. 2000, 165, 4935–4940.
- Gendron, S.; Couture, J.; Aoudjit, F. Integrin Α2β1 Inhibits Fas-Mediated Apoptosis in T Lymphocytes by Protein Phosphatase 2A-Dependent Activation of the MAPK/ERK Pathway. J. Biol. Chem. 2003, 278, 48633–48643.
- Bazan-Socha, S.; Mastalerz, L.; Cybulska, A.; Zareba, L.; Kremers, R.; Zabczyk, M.; Pulka, G.; Iwaniec, T.; Hemker, C.; Undas, A. Prothrombotic State in Asthma Is Related to Increased Levels of Inflammatory Cytokines, IL-6 and TNFα, in Peripheral Blood. Inflammation 2017, 40, 1125–1235.
- Kuczia, P.; Zuk, J.; Iwaniec, T.; Soja, J.; Dropinski, J.; Malesa-Wlodzik, M.; Zareba, L.; Bazan, J.G.; Undas, A.; Bazan-Socha, S. Citrullinated Histone H3, a Marker of Extracellular Trap Formation, Is Increased in Blood of Stable Asthma Patients. Clin. Transl. Allergy 2020, 10, 31.
- Global Initiative for Asthma—Global Initiative for Asthma—GINA. Available online: (accessed on 26 September 2018).

