 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Mhd Mouayad Zuraik | + 2359 word(s) | 2359 | 2021-06-16 08:36:39 | | | |
| 2 | Ron Wang | Meta information modification | 2359 | 2021-06-23 09:46:36 | | |
Video Upload Options
Selenium is an essential trace element. Although this chalcogen forms a wide variety of compounds, there are surprisingly few small-molecule organic selenium compounds (OSeCs) in biology. Besides its more prominent relative selenocysteine (SeCys), the amino acid selenomethionine (SeMet) is one example. SeMet is synthesized in plants and some fungi and, via nutrition, finds its way into mammalian cells. In contrast to its sulfur analog methionine (Met), SeMet is extraordinarily redox active under physiological conditions and via its catalytic selenide (RSeR’)/selenoxide (RSe(O)R’) couple provides protection against reactive oxygen species (ROS) and other possibly harmful oxidants. In contrast to SeCys, which is incorporated via an eloquent ribosomal mechanism, SeMet can enter such biomolecules by simply replacing proteinogenic Met. Interestingly, eukaryotes, such as yeast and mammals, also metabolize SeMet to a small family of reactive selenium species (RSeS). Together, SeMet, proteins containing SeMet and metabolites of SeMet form a powerful triad of redox-active metabolites with a plethora of biological implications. In any case, SeMet and its family of natural RSeS provide plenty of opportunities for studies in the fields of nutrition, aging, health and redox biology.
1. Introduction
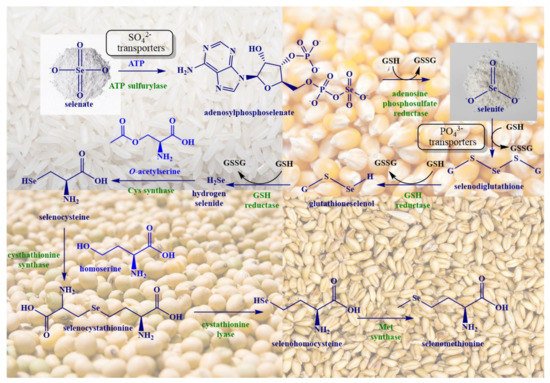
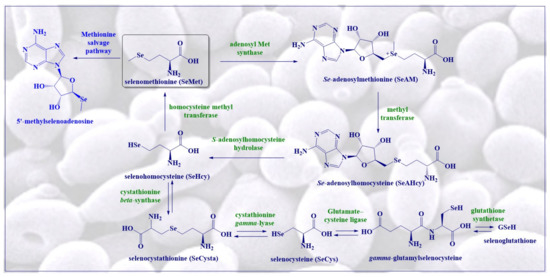
2. Biosynthesis of SeMet in Plants
3. SeMet in Yeast
4. Redox Activity and Catalysis of SeMet
5. Implications of SeMet in Aging and Diseases Related to OS
References
- Fernández-Lázaro, D.; Fernandez-Lazaro, C.I.; Mielgo-Ayuso, J.; Navascués, L.J.; Martínez, A.C.; Seco-Calvo, J. The Role of Selenium Mineral Trace Element in Exercise: Antioxidant Defense System, Muscle Performance, Hormone Response, and Athletic Performance. A Systematic Review. Nutrients 2020, 12, 1790.
- Roy, G.; Sarma, B.K.; Phadnis, P.P.; Mugesh, G. Selenium-containing enzymes in mammals: Chemical perspectives. J. Chem. Sci. 2005, 117, 287–303.
- Kobayashi, R.; Hasegawa, M.; Kawaguchi, C.; Ishikawa, N.; Tomiwa, K.; Shima, M.; Nogami, K. Thyroid function in patients with selenium deficiency exhibits high free T4 to T3 ratio. Clin. Pediatr. Endocrinol. 2021, 30, 19–26.
- Yamashita, Y.; Yabu, T.; Yamashita, M. Discovery of the strong antioxidant selenoneine in tuna and selenium redox metabolism. World J. Biol. Chem. 2010, 1, 144–150.
- Leblanc, K.L.; Smith, M.S.; Wallschläger, D. Production and Release of Selenocyanate by Different Green Freshwater Algae in Environmental and Laboratory Samples. Environ. Sci. Technol. 2012, 46, 5867–5875.
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203.
- Le, D.T.; Liang, X.; Fomenko, D.E.; Raza, A.S.; Chong, C.-K.; Carlson, B.A.; Hatfield, D.L.; Gladyshev, V.N. Analysis of Methionine/Selenomethionine Oxidation and Methionine Sulfoxide Reductase Function Using Methionine-Rich Proteins and Antibodies against Their Oxidized Forms†. Biochemistry 2008, 47, 6685–6694.
- Burk, R.F. Selenium: Recent clinical advances. Curr. Opin. Gastroenterol. 2001, 17, 162–166.
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695.
- Solovyev, N.; Drobyshev, E.; Blume, B.; Michalke, B. Selenium at the Neural Barriers: A Review. Front. Neurosci. 2021, 15.
- Kang, D.; Lee, J.; Wu, C.; Guo, X.; Lee, B.J.; Chun, J.-S.; Kim, J.-H. The role of selenium metabolism and selenoproteins in cartilage homeostasis and arthropathies. Exp. Mol. Med. 2020, 52, 1198–1208.
- Liu, Q.; Zhao, X.; Ma, J.; Mu, Y.; Wang, Y.; Yang, S.; Wu, Y.; Wu, F.; Zhou, Y. Selenium (Se) plays a key role in the biological effects of some viruses: Implications for COVID-19. Environ. Res. 2021, 196, 110984.
- Hou, L.; Lin, Z.; Xu, A.; Le, G.; Ge, L.; Liu, S.; Muhmood, A.; Gan, F.; Huang, K. Combined protective effects of icariin and selenomethionine on novel chronic tubulointerstitial nephropathy models in vivo and in vitro. Br. J. Nutr. 2021, 1–33, 1–11.
- Falandysz, J. Selenium in Edible Mushrooms. J. Environ. Sci. Health Part C 2008, 26, 256–299.
- Klimaszewska, M.; Górska, S.; Dawidowski, M.; Podsadni, P.; Turło, J. Biosynthesis of Se-methyl-seleno-l-cysteine in Basidiomycetes fungus Lentinula edodes (Berk.) Pegler. SpringerPlus 2016, 5, 1–8.
- Li, H.; McGrath, S.P.; Zhao, F. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102.
- Zhao, X.Q.; Mitani, N.; Yamaji, N.; Shen, R.F.; Ma, J.F. Involvement of Silicon Influx Transporter OsNIP2;1 in Selenite Uptake in Rice. Plant Physiol. 2010, 153, 1871–1877.
- Liang, Y.; Su, Y.; Li, L.; Huang, X.; Panhwar, F.H.; Zheng, T.; Tang, Z.; Ei, H.H.; Farooq, M.U.; Zeng, R.; et al. Quick selenium accumulation in the selenium-rich rice and its physiological responses in changing selenium environments. BMC Plant Biol. 2019, 19, 1–11.
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235.
- Winkel, L.H.; Vriens, B.; Jones, G.D.; Schneider, L.S.; Pilon-Smits, E.; Bañuelos, G.S. Selenium Cycling Across Soil-Plant-Atmosphere Interfaces: A Critical Review. Nutrients 2015, 7, 4199–4239.
- Ng, B.; Anderson, J.W. Synthesis of selenocysteine by cysteine synthases from selenium accumulator and non-accumulator plants. Phytochemistry 1978, 17, 2069–2074.
- Van Hoewyk, D.; Çakir, O. Manipulating Selenium Metabolism in Plants: A Simple Twist of Metabolic Fate Can Alter Selenium Tolerance and Accumulation. In Plant Ecophysiology; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.-Q., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 11, pp. 165–176.
- Zhou, Z.S.; Smith, A.E.; Matthews, R.G. l-selenohomocysteine: One-step synthesis from l-selenomethionine and kinetic analysis as substrate for methionine synthases. Bioorg. Med. Chem. Lett. 2000, 10, 2471–2475.
- Gajda, J.; Potrzebowski, M.J.; Bujacz, A.; Bujacz, G. Application of the 77Se Solid State NMR for Investigation of Bioorganic Compounds—The Case of Selenomethionine. Phosphorus Sulfur Silicon Relat. Elements 2008, 183, 1061–1066.
- Al Ghanem, A.; Nasim, M.J.; Alnahas, F.; Ney, Y.; Weiss, A.-V.; Koch, M.; Schneider, M.; Jacob, C. Incredible edible selenium nanoparticles produced by food-grade microorganisms. Curr. Nutraceuticals 2020, 1, 1–10.
- Herrero, E.; Wellinger, R.E. Yeast as a model system to study metabolic impact of selenium compounds. Microb. Cell 2015, 2, 139–149.
- Hahn, G.A.; Brown, J.W. Properties of a methionyl-tRNA systhetase from Sarcina lutea. Biochim. Biophys. Acta (BBA) Enzym. 1967, 146, 264–271.
- Kitajima, T.; Chiba, Y. Selenomethionine metabolism and its toxicity in yeast. Biomol. Concepts 2013, 4, 611–616.
- Chiao, J.S.; Peterson, W.H. Yeasts, Methionine and Cystine Contents. J. Agric. Food Chem. 1953, 1, 1005–1008.
- Ouerdane, L.; Mester, Z. Production and Characterization of Fully Selenomethionine-Labeled Saccharomyces cerevisiae. J. Agric. Food Chem. 2008, 56, 11792–11799.
- Encinar, J.R.; Ouerdane, L.; Buchmann, W.; Tortajada, J.; Lobinski, R.; Szpunar, J. Identification of Water-Soluble Selenium-Containing Proteins in Selenized Yeast by Size-Exclusion-Reversed-Phase HPLC/ICPMS Followed by MALDI-TOF and Electrospray Q-TOF Mass Spectrometry. Anal. Chem. 2003, 75, 3765–3774.
- Rao, Y.; Mc Cooeye, M.; Windust, A.; Bramanti, E.; D’Ulivo, A.; Mester, Z. Mapping of Selenium Metabolic Pathway in Yeast by Liquid Chromatography−Orbitrap Mass Spectrometry. Anal. Chem. 2010, 82, 8121–8130.
- Craddock, V.M. Reactivity of selenomethionine in nucleic acid methylase reactions in the rat. Chem. Interact. 1972, 5, 207–211.
- Kitajima, T.; Jigami, Y.; Chiba, Y. Cytotoxic Mechanism of Selenomethionine in Yeast*. J. Biol. Chem. 2012, 287, 10032–10038.
- Miranda, S.; Wang, Y.; Purdie, N.; Osborne, V.; Coomber, B.; Cant, J. Selenomethionine stimulates expression of glutathione peroxidase 1 and 3 and growth of bovine mammary epithelial cells in primary culture. J. Dairy Sci. 2009, 92, 2670–2683.
- Briviba, K.; Roussyn, I.; Sharov, V.S.; Sies, H. Attenuation of oxidation and nitration reactions of peroxynitrite by selenomethionine, selenocystine and ebselen. Biochem. J. 1996, 319, 13–15.
- Roussyn, I.; Briviba, K.; Masumoto, H.; Sies, H. Selenium-Containing Compounds Protect DNA fromSingle-Strand Breaks Caused by Peroxynitrite. Arch. Biochem. Biophys. 1996, 330, 216–218.
- Padmaja, S.; Squadrito, G.; Lemercier, J.-N.; Cueto, R.; Pryor, W.A. Rapid oxidation of dl-selenomethionine by peroxynitrite. Free Radic. Biol. Med. 1996, 21, 317–322.
- Alvarez, B.; Ferrer-Sueta, G.; Freeman, B.A.; Radi, R. Kinetics of Peroxynitrite Reaction with Amino Acids and Human Serum Albumin. J. Biol. Chem. 1999, 274, 842–848.
- Krause, R.J.; Glocke, S.C.; Sicuri, A.R.; Ripp, A.S.L.; Elfarra, A.A. Oxidative Metabolism of Seleno-l-methionine tol-Methionine Selenoxide by Flavin-Containing Monooxygenases. Chem. Res. Toxicol. 2006, 19, 1643–1649.
- Rahmanto, A.S.; Davies, M.J. Catalytic activity of selenomethionine in removing amino acid, peptide, and protein hydroperoxides. Free Radic. Biol. Med. 2011, 51, 2288–2299.
- Rahmanto, A.S.; Davies, M.J. Selenium-containing amino acids as direct and indirect antioxidants. IUBMB Life 2012, 64, 863–871.
- Skaff, O.; Pattison, D.I.; Morgan, P.E.; Bachana, R.; Jain, V.K.; Priyadarsini, K.I.; Davies, M.J. Selenium-containing amino acids are targets for myeloperoxidase-derived hypothiocyanous acid: Determination of absolute rate constants and implications for biological damage. Biochem. J. 2011, 441, 305–316.

