Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Karolina Szewczyk-Golec | + 5385 word(s) | 5385 | 2021-06-14 10:47:22 | | | |
| 2 | Lily Guo | Meta information modification | 5385 | 2021-06-17 04:35:43 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Szewczyk-Golec, K. Toxoplasmosis. Encyclopedia. Available online: https://encyclopedia.pub/entry/10931 (accessed on 08 August 2026).
Szewczyk-Golec K. Toxoplasmosis. Encyclopedia. Available at: https://encyclopedia.pub/entry/10931. Accessed August 08, 2026.
Szewczyk-Golec, Karolina. "Toxoplasmosis" Encyclopedia, https://encyclopedia.pub/entry/10931 (accessed August 08, 2026).
Szewczyk-Golec, K. (2021, June 16). Toxoplasmosis. In Encyclopedia. https://encyclopedia.pub/entry/10931
Szewczyk-Golec, Karolina. "Toxoplasmosis." Encyclopedia. Web. 16 June, 2021.
Copy Citation
Toxoplasma gondii is an apicomplexan parasite causing toxoplasmosis, a common disease, which is most typically asymptomatic. However, toxoplasmosis can be severe and even fatal in immunocompromised patients and fetuses. Available treatment options are limited, so there is a strong impetus to develop novel therapeutics.
antioxidants
drugs
oxidative stress
toxoplasmosis
Toxoplasma gondii
protozoan parasites
Apicomplexa
1. Introduction
Toxoplasma gondii is a worldwide distributed obligate intracellular protozoan pathogen of humans and more than 30 species of birds and 300 species of mammals [1]. It remains the best model system to study the phylum Apicomplexa, which includes a number of unicellular parasites that are causative agents of several diseases, such as malaria, babesiosis and cryptosporidiosis [2]. All the apicomplexan parasites invade the cells; thus, they have to adapt to the host cell environment and evade the host response. An important element to combat all invading pathogens is the oxidative burst induced by the immune system, so the apicomplexans have to deal with oxidative stress inside their host cells [3].
T. gondii causes toxoplasmosis, which is one of the most common parasitic infections in humans. The prevalence of the disease ranges between 30%–50% of the world’s human population [1]. Toxoplasmosis is a well-known disease entity, and the biology of the parasite is described quite well. Nonetheless, the disease continues to occur with high frequency in the human population and still presents a risk of severe (fatal) disease in some patients. Despite significant progress made in the field of the therapy of human diseases, few advances have been made in the treatment of toxoplasmosis [4]. Available treatment options for toxoplasmosis patients are limited, which underscores the urgent need for searching for new, more effective therapeutics for both the acute and latent forms of the infection. The development of host-targeted adjunctive therapies that could provide new options for toxoplasmosis treatment and reduce the risk of drug resistance seems to be of first importance. The contribution of pro- and antioxidant processes to toxoplasmosis, and its treatment was previously described [5][6][7]. Thus, the use of oxidant and/or antioxidant compounds against the parasite should be strongly considered. So far, the therapeutic effects and safety of drugs against T. gondii infections, including oxidative effects, have been investigated mainly on animal models, most commonly in mice [8]. The available data are still sparse.
Taking into account the relevance of toxoplasmosis as a healthcare issue, this article aims at reviewing the current state of knowledge on the involvement of oxidative stress in the course of the disease and in the host defense, as well as the antioxidant system in the parasite organism. Additionally, the presented review provides new scientific data on the antioxidant mechanisms of action of drugs used in the treatment of toxoplasmosis. Potent candidates for toxoplasmosis treatment, whose mechanisms of action are attributed to the oxidant–antioxidant balance both in the parasite organism and in its host, as well as the potentially supportive action of antioxidant supplements, are discussed. We would like to inspire scientists to further research on the use of compounds modifying oxidative stress in the treatment of toxoplasmosis.
2. Characteristics of Toxoplasma gondii
Toxoplasma gondii was first described in 1908 by Nicolle and Manceaux in the gundi, a small hamster-rodent of Ctenodactylidae [9]. The researchers named it Toxoplasma gondii basing on its morphology (modern Latin: toxo—arc or bow, plasma—life) and the host. In retrospect, the correct name for the parasite should have been T. gundi, but Nicolle and Manceaux identified the host as Ctenodactylus gondi instead of Ctenodactylus gundi, which is a correct name [9]. There are three main genotypes of Toxoplasma gondii (type I, II and III), with different virulence and epidemiological patterns, which largely predominate in Europe and North America [10][11]. Apart from these three main clonal lineages, there are also atypical genotypes, more complex and with higher genetic diversity. Dardé [11] suggests that there are relationships between the genotype and human disease, but these mechanisms are still poorly understood. Recombination may lead to acquiring new pathogenic mechanisms of Toxoplasma and manifest in humans by a severe course of toxoplasmosis, also in immunocompetent patients.
2.1. Life Cycle
The Toxoplasma gondii life cycle includes sporogony (sexual reproduction in definitive hosts) and schizogony (asexual reproduction in intermediate hosts). Cats are the only definitive hosts of this parasite and become infected by eating meat (mostly rodents, birds, or slaughterhouse remains) containing tissue cysts or by ingesting oocysts from the soil [12]. Bradyzoites released from tissue cysts form schizonts in the intestine of cats. After merulation, schizonts release merozoites. Only in cats can the parasite form gametocytes, which further develop into gametes [13]. After fertilization, they form oocysts, which are shed in feces. However, only sporulated oocysts containing sporozoites become infective for intermediate hosts [14]. In intermediate hosts, upon oral uptake, sporozoites transform into the invasive tachyzoite stage. Tachyzoites are proliferative, fast-multiplying forms of the parasite. They can multiply in a variety of cells and eventually encyst in several tissues, particularly in the brain [15]. Tissue cysts can be found in the retina, brain, skeletal and heart muscle [16]. They are the infective stages for intermediate and definitive hosts [15]. Humans can be infected by eating meat containing tissue cysts, which are resistant to gastric digestion and thus infectious orally, whereas tachyzoites are destroyed by gastric juice [12]. Infective tachyzoites develop from bradyzoites, which are released from lysed cysts in the intestine. Rapidly multiplying tachyzoites can transform to metabolically inactive bradyzoites and form tissue cysts. Tissue cysts may remain dormant for a long time, perhaps for the life of the host [13]. Occasionally, rupturing tissue cysts release bradyzoites, which are killed in immunocompetent hosts. However, in immunodeficient hosts, bradyzoites released from tissue cysts may multiply locally and spread to other organs [14][15].
2.2. Routes of Toxoplasma Infection in Humans
To summarize, humans can be infected by three different forms of the parasite, namely tachyzoites, bradyzoites and sporozoites. Oocysts of Toxoplasma, formed in the intestine of Felidae (cats), are excreted in the cat’s feces into the environment. About 2 or 3 days after excretion, oocysts undergo sporulation and become an infective form containing sporozoites [13][14][17]. These forms are resistant to environmental factors and can stay infective for more than one year [13]. It should be noted that humans can be infected by ingestion of vegetables, fruit or water contaminated with sporozoites, but contrary to appearances, it is not the most common way of getting infected. More commonly, people become infected by eating raw, cured, dried, smoked or undercooked meat, particularly lamb and pork, containing tissue cysts with bradyzoites [13][14][17][18][19]. Chicken meat is a source of infection for cats, but also for humans if eaten undercooked [18]. It should be considered that chickens are one of the most important hosts of toxoplasma. The prevalence of toxoplasmosis in chickens raised in backyards reaches up to 100% because they feed on potentially contaminated ground [18]. This makes pasture chickens a significant risk factor for Toxoplasma infection. However, studies on experimentally infected hens suggest that chicken eggs should not be considered as a source of infection [18]. In a case-control study, Jones et al. [19] identified eating raw mollusks, namely oysters, clams and mussels, as a new risk factor for Toxoplasma infection. As mollusks are filter feeders, the parasite concentrates in their organisms. Another potential source of transmission is un-pasteurized milk and milk products [14][17]. Toxoplasma gondii has multiple pathways to infect a human organism, apart from those mentioned. Tachyzoites can be transmitted also by blood transfusion. Congenital toxoplasmosis, which is an effect of vertical transmission of tachyzoites from mother to fetus through the placenta, should be also mentioned [13][14].
2.3. Prophylaxis of Toxoplasma Infection in Humans
In relation to the main sources of the parasite and routes of Toxoplasma transmission, the prevalence of the disease reflects mainly hygienic and dietary practices of human populations [13]. To protect the soil against contamination with sporozoites, litter boxes should be used by cat owners. As it takes several days for oocysts to sporulate, litter boxes should be cleaned up daily, but not by people from increased risk groups, such as pregnant women, or immunosuppressed people [19]. Although there are effective and simple methods to inactivate cysts of the parasite in meat, for example, deep-freezing (−12 °C or lower for at least 24 h), low social awareness of the disease causes Toxoplasma eradication still ineffective. Most people are aware of the association of toxoplasmosis with cats, but they are rather unaware of exposure to the parasite during their daily activities, especially the risk associated with undercooked meat [19]. It should be borne in mind that the parasite can also be transmitted if care is not taken to wash hands thoroughly after cutting meat and during meat cooking [18]. Hence the need arises to educate society about the risk of toxoplasmosis transmission, among others, by popularizing safe-cooking guidelines and to take into account other sources of the parasite in toxoplasmosis prevention. Reduction of T. gondii contamination of meat and increased education of health professionals and the public regarding risk factors could help to further reduce the burden of toxoplasmosis [19].
2.4. Toxoplasmosis Course in Humans
There are three clinical forms of the disease, including ocular, congenital and cerebral ones. Toxoplasma gondii is potentially responsible for significant morbidity and mortality in the congenitally infected fetus (especially in the first trimester of pregnancy) because it can cause a serious disease course and even miscarriage. In a congenital disease course, hepatitis and pneumonia may be followed by central nervous system involvement resulting in hydrocephalus, retinochoroiditis and cerebral calcifications [10]. In immunocompetent humans, the disease is said to be latent (clinically asymptomatic), but the disease course can be severe in immunocompromised or immunosuppressed patients [10][13]. In an oligosymptomatic course, the disease may produce mild, flu-like illness. In less than 10% of cases, it causes mononucleosis-like syndrome with headache, malaise, fever, cervical lymphadenopathy and fatigue [13]. Some tachyzoites can evade the immune response or drugs used in the treatment and transform into bradyzoites inside quiescent cysts, long-term forms of the parasite. Additionally, Toxoplasma can use immune cells to migrate and infiltrate the brain. Recent investigations indicate that toxoplasmosis may increase the risk of brain cancer [13][20]. In the brain, Toxoplasma prefers neurons because these cells do not react to inflammatory cytokines. Thus, a strong anti-parasitic immune response is not induced [13]. As a result, central nervous system (CNS) toxoplasmosis, or toxoplasmic encephalitis (TE), which is the most severe (even fatal) course of the disease, may develop in immunocompromised individuals. Encephalitis is the predominant clinical manifestation of toxoplasmosis in acquired immune deficiency syndrome (AIDS) patients and is believed to be due to the reactivation of latent infections [15]. Clinically, the most common manifestations of TE are neurological abnormalities and brain abscesses accompanied by fever [13]. Interestingly, Toxoplasma can change the behavior of hosts, including humans. Infected rodents experience less fear of feline odors, which makes them more likely to be eaten by the parasite definite host [21]. The possible relationship of Toxoplasma infection with mental illness in humans has focused the attention of many researchers [22]. There have been numerous studies on T. gondii infection and its impact on human behavior and mental diseases [23]. Nonetheless, the effect of the parasite on human behavior is still less certain than in rodents [24]. The influence of T. gondii on the increased probability of developing psychiatric disorders, including schizophrenia, bipolar disorder and obsessive-compulsive disorder, has been confirmed [21]. However, no parasite genes or effector proteins responsible for parasitic behavioral changes have been identified [21]. Postulated mechanisms linking toxoplasmosis and behavior include generalized inflammation, alteration in endocrine signaling and changes in neurotransmitter pathways [24]. The main weakness of the studies conducted so far is that they rely on the correlation between T. gondii seropositivity and behavioral or cognitive outcomes but do not assess directly the causal role of the infection in a given pattern of behavior [23]. Taking into account study limitations and the results of meta-analyses, it could be suggested that the effect of the parasite on human behavior or disease might be rather mild. It is likely that the influence of T. gondii on human behavior and mental health may be dependent on a number of factors, including the parasite strain type, route and duration of infection, host genotype, and others [23]. Undoubtedly, more research is needed in this area.
3. Oxidative Stress in Toxoplasma gondii Infection
Oxidative stress is defined as an imbalance between the production of reactive oxygen species (ROS) and the antioxidant system of the organism [25][26]. ROS include all highly reactive and unstable derivatives of molecular oxygen, such as hydrogen peroxide (H2O2), superoxide anion (O2−) and the most dangerous hydroxyl radical (OH∙), which is formed in the Fenton and/or Haber–Weiss reaction [27][28]. ROS homeostasis is closely related to many other reactive molecules, such as reactive carbonyl species (RCS) and reactive nitrogen species (RNS) [29]. Since ROS are produced as a consequence of oxygen metabolism, it is impossible to avoid them in aerobic organisms. They are generated in the cytosol and in such organelles, such as mitochondria and peroxisomes [30][31]. At physiological levels, ROS are involved in cell signaling processes, but enhanced oxidative stress due to the excessive ROS formation may cause damage to all cellular macromolecules such as lipids, proteins and nucleic acids, ultimately leading to cell death [7][31][32]. The characteristic action of ROS is the degradation of polyunsaturated fatty acids in the process termed lipid peroxidation, which leads to the production of harmful molecules, including malondialdehyde (MDA) [33]. MDA is a highly reactive aldehyde that causes oxidative damage in tissues. MDA belongs to thiobarbituric acid reactive substances (TBARS) and is a major TBARS in the organism. Therefore, for the sake of method simplicity, the TBARS measurement is commonly used to assess the MDA concentration [34]. Both TBARS and MDA are considered to be the most important markers of oxidative stress and lipid peroxidation in biological samples [35].
3.1. Oxidant–Antioxidant Balance in Humans
ROS are related to a wide variety of human disorders, such as chronic inflammation, age-related diseases and cancer [36][37]. Apart from that, ROS are also essential for various biological functions, including cell survival, cell growth, proliferation and differentiation, and immune response [38][39]. In phagocytes, ROS are generally produced in a specific process with nicotinamide adenine dinucleotide phosphate (NADPH) oxidase or by the mitochondrial respiratory chain [31][40]. NADPH oxidase plays a vital role in inflammatory processes by catalyzing the production of the superoxide anion radical and excessive production of other ROS, leading finally to cellular damage. The resulting cellular damage alters the immune response to pathogens and ultimately modifies susceptibility to bacterial, viral and parasitic infections [35].
All oxygen-metabolizing cells are equipped with cellular antioxidants to counteract the harmful effects of ROS action. Detoxification and removal of deleterious oxidants are essential in biological systems to restore redox homeostasis of the cell [30][41]. The complex endogenous antioxidant system includes antioxidant enzymes such as superoxide dismutases (SODs), catalase (CAT), glutathione peroxidases (GPxs) and glutathione reductase (GR) [42][43], as well as non-enzymatic compounds such as reduced glutathione (GSH) and uric acid [44][45]. GSH protects the cell against the detrimental effects of endogenous and exogenous oxidants by conjugation of reactive species and detoxification of lipid peroxidation products [46][47]. Moreover, GSH prevents the conversion of hemoglobin into methemoglobin in the oxidation reaction [7][48]. At present, antioxidant vitamins A, C and E are considered to be the main exogenous antioxidants playing a significant role in neutralizing free radicals and maintaining oxidative homeostasis [49][50].
3.2. Role of Oxidative Stress in Toxoplasma Infection
Oxidative stress plays a significant role in the course of Toxoplasma infection in the organism of both the host and the parasite [51] (see Figure 1). When T. gondii multiplies asexually, it causes cellular disruption and cell death in an infected host. The resulting necrosis attracts inflammatory host cells, such as lymphocytes and monocytes. In the immune response against the parasite, enormous amounts of ROS and RNS are generated [41]. Oxidative stress resulting from the host response is toxic to parasites, but on the other hand, many studies reported that deleterious consequences of parasitic infection in a host organism are the result of defense mechanisms involving increased production of ROS [52]. The processes related to oxidative stress were proved to play a pivotal role in the pathogenesis of toxoplasmosis in animals and humans [53][54].
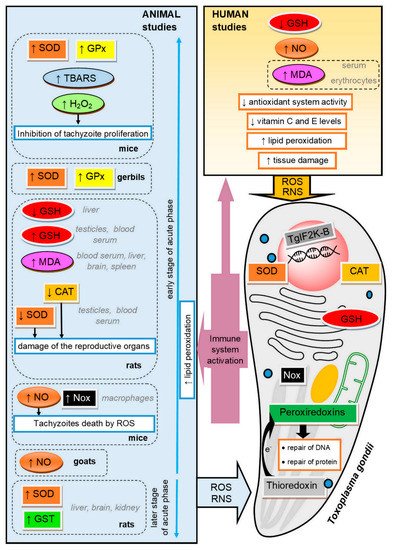
Figure 1. The mechanisms of oxidative injury and antioxidant defense in the course of toxoplasmosis in the parasite and in the host. SOD, superoxide dismutase; GPx, glutathione peroxidase; TBARS, thiobarbituric acid reactive substances; GSH, reduced glutathione; MDA, malondialdehyde; CAT, catalase; NO, nitric oxide; Nox, NADPH–oxidase complex; GST, glutathione S-transferase; RNS, reactive nitrogen species; ROS, reactive oxygen species; TgIF2K-B, Toxoplasma gondii eukaryotic initiation factor 2 (eIF2α) kinase.
3.3. Antioxidant Defense of Toxoplasma gondii
ROS, generated in immune response, are able to kill various intracellular pathogens, including T. gondii. It was found that ROS can inhibit the activity of T. gondii in monocytes of infected animals. For example, high H2O2 concentration may inhibit the intracellular proliferation of tachyzoites [55]. This is why the parasite must protect itself against the oxidative burst imposed by the host [5][55][56]. Toxoplasma cells express SODs (one cytosolic and two mitochondrial), CAT and three peroxiredoxins, including one 1-Cys peroxiredoxin and two 2-Cys peroxiredoxins [5][55][57][58]. SODs and CAT belong to the primary antioxidants involved in the defense against oxidative stress and oxidative metabolic by-products [59]. SODs are metalloproteins that catalyze the dismutation of superoxide anions to form molecular oxygen and hydrogen peroxide [60]. CAT catalyzes hydrogen peroxide conversion to water and oxygen and thereby diminishes its cellular level [56][61]. The intrinsic kinetic properties of CAT and its cytosolic localization suggest that this enzyme might be best suited to the protection against host cell oxidative stress [5]. According to knockout studies, CAT seems to have an important role in invasion and replication inside the parasitophorous vacuoles [6]. T. gondii deficient in CAT exhibited increased susceptibility to exogenous hydrogen peroxide and was less virulent in mice [5]. Peroxiredoxins have phospholipase activity and are able to detoxify H2O2 in the presence of dithiothreitol [57]. Increased peroxiredoxin expression was found to be associated with the protection of the parasite against oxidative stress [6]. In organisms possessing CAT, the peroxiredoxins are unlikely to play a pivotal role in antioxidant defense, and recent studies suggest that these enzymes might fulfil quite distinct functions [5]. However, it seems likely that peroxiredoxins are important to T. gondii in view of the fact that the parasite is able to survive the knockout of catalase [57].
The classical antioxidant systems such as thioredoxins (Trx) and GSH are also suggested to occur in Toxoplasma, which is corroborated by the presence of trx genes in the transcriptome [62]. The role of the Trx system is to deliver electrons to the peroxiredoxins, which enables the removal of ROS and RNS. In addition, this system is involved in the repair of oxidative damage of DNA and proteins [63]. Thioredoxin reductase, which catalyzes the reduction of the oxidized form of thioredoxin with the consumption of NADPH, has the ability to maintain the Trx redox state, thus protecting the parasite against free radical damage in host immune cells [55].
3.4. Adaptative Response of Toxoplasma gondii to Oxidative Stress
Regardless of the variety of antioxidant systems present in T. gondii, tachyzoites are highly susceptible to exogenous and endogenous oxidative stress. Minor changes in the redox balance of the parasite cell can lead to the destruction of its oxidative homeostasis and ultimately its death [57]. In eukaryotes, environmental stress induces the integrated stress response via phosphorylation of translation initiation factors, including eukaryotic initiation factor 2 (eIF2) [64]. Interestingly, a similar adaptive mechanism was observed in T. gondii. Augusto et al. [65] demonstrated that the Toxoplasma eIF2 kinase TgIF2K-B is activated in response to oxidative stress and affords protection. The presence of TgIF2K-B enables the parasite to regulate its replication rate under oxidative stress as the activation of this enzyme influences the maintenance of oxidant–antioxidant balance in the parasite organism. For Toxoplasma, the critical moment of adaptation is the differentiation into bradyzoite cysts. Parasites lacking TgIF2K-B would likely have reduced transmission through the predation route due to their compromised ability to form infectious tissue cysts [65].
3.5. Antioxidant Defense of the Host during Toxoplasma Infection
The enzymatic antioxidant defense is one of the mechanisms that protect the host cells against an excess of free radicals due to parasitic infections [42][66]. Nazarlu et al. [67] observed a decrease in SOD and CAT activity in the blood serum, but also in the testes of rats infected with T. gondii. It was also found that the activity of SOD was reduced in erythrocytes of gerbils with toxoplasmosis [30][41], while Bahrami et al. [68] observed a significant increase in GPx activity in the blood of rats infected with T. gondii on the seventh day after the infection.
Another compound that plays a relevant role in antioxidant mechanisms is GSH, as the thiol group of GSH transmits a reducing equivalent of ROS [69]. A decrease in GSH concentration may indicate the activation of antioxidant mechanisms in response to the T. gondii infection. Significant depletion in the GSH level, observed in rats infected with T. gondii, is supposed to be an important factor in toxicity, leading to host brain tissue damage and a reduction in the activities of antioxidant enzymes [68][70][71]. Similarly, Nazarlu et al. [67] observed decreased GSH levels in the blood serum and in the testes of rats infected with T. gondii. The authors believe that decreased GSH levels in chronic toxoplasmosis could change the detoxifying capacity of reproductive tissue and thus cause oxidative damage to reproductive organs and, consequently, affect adversely fertility. A decrease in GSH concentration was also detected in patients with toxoplasmosis, which suggests the occurrence of changes in the oxidant–antioxidant balance as a mechanism of tissue damage in cases of toxoplasmosis in humans [50][54]. The GSH action is suggested to be increased by vitamins C and E [54]. In addition to the enzymes, antioxidant vitamins are also involved in maintaining cell homeostasis and neutralizing free radicals. Vitamin C is a fundamental supplement involved in the repair of tissues, while vitamin E is a fat-soluble antioxidant interrupting the propagation of ROS [31][72]. Khaleel et al. [50] observed decreased levels of antioxidant vitamins in women with toxoplasmosis.
3.6. Oxidative Stress in the Early Stages of the Acute Phase of Toxoplasmosis
It is believed that, during the acute phase of toxoplasmosis, ROS are intensively produced, and oxidative stress is induced in the tissues of infected animals as the host defense against the infection. T. gondii seropositive cats, even when asymptomatic, show an increase in ROS levels [73]. The host immune response to infections typically involves phagocytosis after the activation of mononuclear phagocytes by lymphokines [74][75]. Although the NADPH–oxidase complex (Nox), which produces ROS, is expressed in almost every mammalian tissue, its function of producing ROS in host defense against pathogens is more pronounced in phagocytes [76]. Hence, the activation of parasiticidal mechanisms via ROS generation occurs mainly in host phagocytes. Nox-mediated ROS generation, as a factor involved in regulating the intracellular survival of T. gondii parasites within macrophages, was demonstrated in mice by Matta et al. [77].
An antiparasitic effect on protozoa was also revealed for nitric oxide (NO) derived from macrophages [78]. T. gondii infection is associated with an increased NO level, which is generated in an attempt to control the infection. Tonin et al. [79] noted an increase in NO levels in goats with toxoplasmosis. NO can directly cause the death of tachyzoites or stimulate the production of the heat-shock protein 70 in tachyzoites of both virulent and non-virulent strains, contributing to their conversion to the bradyzoite stage and cyst formation [79][80].
3.7. Oxidative Stress in the Later Stages of the Acute Phase of Toxoplasmosis
The vast majority of research has focused on describing oxidative stress during the early stages of the acute phase of toxoplasmosis. However, there are a few studies investigating the oxidant–antioxidant balance in the later stages of the acute phase of T. gondii infection. Türkoğlu et al. [81] observed increased SOD and glutathione S-transferase (GST) activities 30 days after infection in the brain, liver and kidney of rats. The researchers suggest that the increased antioxidant defense persisting later on in the acute phase can be used to diagnose T. gondii infection and may be helpful, especially in cases where the diagnosis of T. gondii infection is difficult in the earlier stage of the acute phase.
3.8. Oxidative Damage in the Host during Toxoplasma Infection
The rapid release of ROS and NO plays an important role against T. gondii, but also contributes to oxidative injury, inflicting tissue damage and the disease pathology [68][82][83]. Oxidative stress leads to intracellular lysosomal membrane damage, which is followed by apoptosis or necrosis [84]. The reduced activity of the defense system protecting tissue against free radical damage in Toxoplasma seropositive patients and animals is associated with increased lipid peroxidation [48][53]. Atmaca et al. [41] suggested that T. gondii induces lipid peroxidation in infected gerbils. In rats infected with T. gondii, an increased MDA concentration was observed in the liver cells [71], as well as in the blood serum and in the testes [67]. Moreover, [70] an increase in TBARS concentration was found in the brain of mice infected with T. gondii. Some authors reported a significant increase in the concentration of MDA in the serum of humans with chronic toxoplasmosis. Kiran et al. [47], Al-Kuraishy et al. [85] and Yazar et al. [53] reported that MDA levels were significantly higher in asymptomatic Toxoplasma seropositive patients compared to healthy subjects. There was no correlation between age or gender and the MDA concentration. An increased concentration of MDA was also observed in the erythrocytes of women with toxoplasmosis in comparison with the control group [48].
Despite the available literature data, more research on T. gondii infection in combination with other diseases is still needed to better understand the changes caused by this parasite in its host and to establish the diagnosis and appropriate treatment.
4. Oxidant–Antioxidant Effects in Toxoplasmosis Treatment
The gold standard in toxoplasmosis treatment is the combination of pyrimethamine and sulfadiazine (pyr–sulf), targeting the active stage of infection [4]. There are also other therapies available, including pyrimethamine in combination with some antibiotics, as well as monotherapy with sulfamethoxazole-trimethoprim (ST) or atovaquone [8][86]. Nevertheless, none of these therapies was found to be superior to pyr–sulf, and no therapeutic is able to eliminate T. gondii cysts from the infected host [87]. The main targets of typical anti-Toxoplasma drugs are enzymes involved in the folate synthesis pathway, namely dihydrofolate reductase (DHFR) and dihydropteroate synthetase (DHPS) [88]. DHFR is also present in humans, so this therapy may result in a folic acid deficiency, which in turn is possibly responsible for severe hematological side-effects [86]. To date, all current therapeutical strategies have several limitations, including adverse severe side-effects and treatment failure because of drug resistance [89]. In this respect, there is an urgent need for searching alternative compounds with novel mechanisms of action. The numerous drug targets identified against Toxoplasma gondii include the inhibition of the mitochondrial electron transport chain, synthesis of fatty acids, isoprenoid pathway, synthesis of DNA, synthesis of proteins, as well as the action of Toxoplasma gondii calcium-dependent protein kinase 1 (TgCDPK1) [90][91]. In addition, targeted host-directed immunotherapy aimed at activating or suppressing specific elements of the immune system has been considered [91].
In view of the research on the contribution of oxidative stress to the pathology of toxoplasmosis, the chemical compounds that affect the oxidant–antioxidant balance are of high interest as possible novel anti-Toxoplasma therapeutics (see Figure 2). The antioxidant system of T. gondii is known to play an important role as a defense mechanism against oxidative stress imposed by the host and to contribute as a virulence factor in vivo [5].
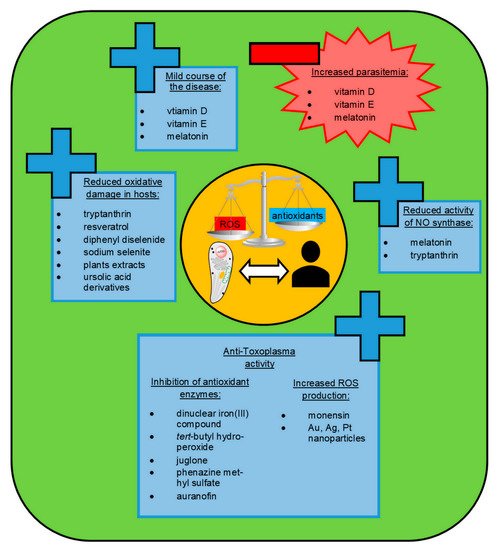
Figure 2. The chemical compounds that may affect the oxidant–antioxidant balance as promising novel anti-Toxoplasma therapeutics. ROS, reactive oxygen species; NO, nitric oxide.
4.1. Inhibition of Antioxidant Defense of Toxoplasma gondii
The inhibition of antioxidant enzymes of the Toxoplasma parasite seems to be a viable target for the development of novel drugs against this pathogen. One of the antioxidant enzymes is CAT, which lacks in most pathogenic protozoans, but it is present in the T. gondii cytoplasm, where it neutralizes the peroxides produced by the host [6]. CAT and SOD seem to be the most important antioxidants of T. gondii, reacting directly to free radicals [59]. Portes et al. [92] demonstrated that the exposure of the Toxoplasma parasite to dinuclear iron (III) compound reduced the activities of metalloenzymes, including SOD and CAT, which are important for the antioxidant protection of the parasite. This agent disturbs the redox balance of T. gondii, inducing parasite death in vitro, with no toxicity to the host cells. Moreover, Akerman et al. [57] demonstrated that Toxoplasma gondii is highly susceptible to oxidative stress that results from the disruption of its redox balance caused by tert-butyl hydroperoxide, juglone and phenazine methyl sulfate, with no effect on the host cells. Those compounds can affect the activity of peroxiredoxins, which are important in the protection of parasite cells against oxidative damage by hydroperoxides [57]. Another promising chemotherapeutic agent against T. gondii seems to be auranofin, which induces the accumulation of ROS in parasites via the inhibition of the thioredoxin reductase enzyme [93]. Thioredoxin reductase is essential for the survival of the parasite as maintaining a thioredoxin-dependent reduction state helps pathogens to resist oxidative-burst injury from host cells [55]. The results of Andrade et al. [94] revealed high efficiency of auranofin activity against T. gondii in vitro and in vivo, suggesting that it may be an effective alternative treatment for acute toxoplasmosis in the future. The induction of Toxoplasma parasite death due to increased ROS production was also reported as an impact of monensin treatment [56]. This drug is known to suppress the expression of the majority of T. gondii genes [95]. However, while monensin is used widely during anti-Toxoplasma therapy in animals, its toxicity precludes the use in humans.
The combination of pyrimethamine and sulfadiazine (the drugs of choice for toxoplasmosis) can suppress tachyzoite growth but has no effect on bradyzoites [96]. Therefore, most current therapies are ineffective against the latent stage of infection. Augusto et al. [65], using a CRISPR/Cas9 technique, successfully generated a genetic knockout of eIF2 kinase TgIF2K-B in the Toxoplasma parasite. As a result, the parasite failed to phosphorylate TgIF2α in response to oxidative stress, declining the ability to differentiate into tissue cysts. Those results may suggest that inhibition of TgIF2K-B may be a promising direction to develop new therapies based on diminished antioxidant protection, especially against persistent infection of T. gondii in its host.
4.2. Inorganic Nanoparticles
A possible source of alternative and effective agents in the treatment of Toxoplasma gondii infections may be also inorganic nanoparticles (NPs). The study of Adeyemi et al. [97] demonstrated that gold, silver, and platinum nanoparticles had promising in vitro anti-Toxoplasma activity without detectable host cell toxicity. The measurement through a fluorescent probe provided direct evidence that treatment with the use of nanoparticles promoted ROS production with a consequent contribution to parasite death. Other papers also show that silver nanoparticles (AgNPs), which have the broadest biomedical applications of all nanoparticles, may cause oxidative stress and DNA damage [98][99].
In studies on the impact of repeated administration of AgNPs on the antioxidant status of Wistar rats, an increase in lipid peroxidation and alteration of antioxidant enzymes were revealed in animal tissues and serum [98]. Thus, potential risks of host cell damage may be associated with the use of silver nanoparticles in the anti-Toxoplasma treatment. Hudecova et al. [99] confirmed oxidative DNA damage induced by AgNPs in human kidney cells in vitro. However, this oxidative damage was reported to be neutralized by pre-treatment with bioactive compounds, such as Gentiana extracts. An alternate solution may be the use of silver nanoparticles synthesized in combination with natural extracts of plants. Alajmi et al. [83] reported that AgNPs green synthesized with date palm and nabka exhibited some anti-Toxoplasma effects and significantly decreased the level of lipid peroxidation and NO concentrations with a concomitant increase in antioxidant enzymes activity in liver homogenate of mice infected with T. gondii.
Undoubtedly, the results of the presented studies support the idea that oxidative stress may be a good target for toxoplasmosis treatment. However, only limited data concerning oxidant–antioxidant effects of anti-toxoplasmosis treatment are available, and thus, further research is needed in this area.
5. Plant-Derived Antioxidants in Toxoplasmosis Treatment
The pathophysiology of toxoplasmosis is known to be related to the increased risk of oxidative stress in host cells due to the activation of inflammatory response against the parasite [85]. In chronic acquired toxoplasmosis, the level of MDA is elevated and can reflect indirectly the degree of oxidative cell damage [54]. Therefore, all molecules that diminish oxidative stress in the host cells may provide viable sources of alternative therapies for this parasitic infection (see Figure 2). The literature gives new insights into various natural extracts of plants with antioxidant properties, which may be good candidates for the discovery of novel drugs or may be useful as an alternative or adjuvant treatment option in the course of toxoplasmosis. It is worth mentioning that the current antimalarial drugs of choice (chloroquine and artemisinin derivatives) were derived from plants [100].
References
- Flegr, J.; Prandota, J.; Sovičková, M.; Israili, Z.H. Toxoplasmosis—A Global Threat. Correlation of Latent Toxoplasmosis with Specific Disease Burden in a Set of 88 Countries. PLoS ONE 2014, 9, e90203.
- Kim, K.; Weiss, L.M. Toxoplasma gondii: The model apicomplexan. Int. J. Parasitol. 2004, 34, 423–432.
- Vonlaufen, N.; Kanzok, S.M.; Wek, R.C.; Sullivan, W.J., Jr. Stress response pathways in protozoan parasites. Cell. Microbiol. 2008, 10, 2387–2399.
- Konstantinovic, N.; Guegan, H.; Stäjner, T.; Belaz, S.; Robert-Gangneux, F. Treatment of toxoplasmosis: Current options and future perspectives. Food Waterborne Parasitol. 2019, 15, e00036.
- Kwok, L.Y.; Schlüter, D.; Clayton, C.; Soldati, D. The antioxidant systems in Toxoplasma gondii and the role of cytosolic catalase in defence against oxidative injury. Mol. Microbiol. 2003, 51, 47–61.
- Bosch, S.S.; Kronenberger, T.; Meissner, K.A.; Zimbres, F.M.; Stegehake, D.; Izui, N.M.; Schettert, I.; Liebau, E.; Wrenger, C. Oxidative Stress Control by Apicomplexan Parasites. BioMed Res. Int. 2015, 2015, 1–10.
- Dincel, G.C.; Atmaca, H.T. Role of oxidative stress in the pathophysiology of Toxoplasma gondii infection. Int. J. Immunopathol. Pharmacol. 2016, 29, 226–240.
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal Models, and Current Clinical Practice. Clin. Microbiol. Rev. 2018, 31, e00057-17.
- Dubey, J.P. The History of Toxoplasma gondii—The First 100 Years. J. Eukaryot. Microbiol. 2008, 55, 467–475.
- Dalimi, A.; Abdoli, A. Latent Toxoplasmosis and Human. Iran. J. Parasitol. 2012, 7, 1–17.
- Dardé, M. Toxoplasma gondii, “new” genotypes and virulence. Parasite 2008, 15, 366–371.
- Tenter, A.M. Toxoplasma gondii in animals used for human consumption. Memórias Do Inst. Oswaldo Cruz 2009, 104, 364–369.
- Elsheikha, H.M.; Marra, C.M.; Zhu, X.-Q. Epidemiology, Pathophysiology, Diagnosis, and Management of Cerebral Toxoplasmosis. Clin. Microbiol. Rev. 2020, 34, 1–28.
- Schlüter, D.; Däubener, W.; Schares, G.; Groß, U.; Pleyer, U.; Lüder, C. Animals are key to human toxoplasmosis. Int. J. Med. Microbiol. 2014, 304, 917–929.
- Dubey, J. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 1998, 28, 1019–1024.
- Song, H.B.; Jung, B.-K.; Kim, J.H.; Lee, Y.-H.; Choi, M.-H.; Kim, J.H. Investigation of tissue cysts in the retina in a mouse model of ocular toxoplasmosis: Distribution and interaction with glial cells. Parasitol. Res. 2018, 117, 2597–2605.
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasites Vectors 2020, 13, 1–13.
- Dubey, J.P. Toxoplasma gondii Infections in Chickens (Gallus domesticus): Prevalence, Clinical Disease, Diagnosis and Public Health Significance. Zoonoses Public Health 2010, 57, 60–73.
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk Factors for Toxoplasma gondii Infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884.
- Hodge, J.M.; Coghill, A.E.; Kim, Y.; Bender, N.; Smith-Warner, S.A.; Gapstur, S.; Teras, L.R.; Grimsrud, T.K.; Waterboer, T.; Egan, K.M. Toxoplasma gondii infection and the risk of adult glioma in two prospective studies. Int. J. Cancer 2021, 148, 2449–2456.
- Tong, W.H.; Pavey, C.; O’Handley, R.; Vyas, A. Behavioral biology of Toxoplasma gondii infection. Parasites Vectors 2021, 14, 1–6.
- Xiao, J.; Prandovszky, E.; Kannan, G.; Pletnikov, M.V.; Dickerson, F.; Severance, E.G.; Yolken, R.H. Toxoplasma gondii: Biological Parameters of the Connection to Schizophrenia. Schizophr. Bull. 2018, 44, 983–992.
- Johnson, H.J.; Koshy, A.A. Latent Toxoplasmosis Effects on Rodents and Humans: How Much is Real and How Much is Media Hype? mBio 2020, 11, 1–10.
- Johnson, S.K.; Johnson, P.T. Toxoplasmosis: Recent Advances in Understanding the Link Between Infection and Host Behavior. Annu. Rev. Anim. Biosci. 2021, 9, 249–264.
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8.
- Kehrer, J.P. The Haber–Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50.
- Southorn, P.A.; Powis, G. Free Radicals in Medicine. I. Chemical Nature and Biologic Reactions. Mayo Clin. Proc. 1988, 63, 381–389.
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265.
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Interact. 2014, 224, 164–175.
- Kamata, H.; Hirata, H. Redox Regulation of Cellular Signalling. Cell. Signal. 1999, 11, 1–14.
- Nordberg, J.; Arnér, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312.
- Halliwell, B. Oxygen and nitrogen are pro-carcinogens. Damage to DNA by reactive oxygen, chlorine and nitrogen species: Measurement, mechanism and the effects of nutrition. Mutat. Res. Toxicol. Environ. Mutagen. 1999, 443, 37–52.
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328.
- Dalle-Donne, I.; Rossi, R.; Colombo, R.; Giustarini, D.; Milzani, A. Biomarkers of Oxidative Damage in Human Disease. Clin. Chem. 2006, 52, 601–623.
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed Res. Int. 2014, 2014, 1–19.
- Shakiba, Y.; Mahmoudi, M. Chronic Inflammation and Oxidative Stress as a Major Cause of Age- Related Diseases and Cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80.
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and oxidative stress in human diseases: From molecular mechanisms to novel treatments. Int. J. Mol. Sci. 2019, 20, 4472.
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95.
- Kim, J.H.; Lee, J.; Bae, S.-J.; Kim, Y.; Park, B.-J.; Choi, J.-W.; Kwon, J.; Cha, G.-H.; Yoo, H.J.; Jo, E.-K.; et al. NADPH oxidase 4 is required for the generation of macrophage migration inhibitory factor and host defense against Toxoplasma gondii infection. Sci. Rep. 2017, 7, 6361.
- Scherz-Shouval, R.; Elazar, Z. ROS, mitochondria and the regulation of autophagy. Trends Cell Biol. 2007, 17, 422–427.
- Çinar, A.N. Evaluation of oxidative stress, hematological and biochemical parameters during Toxoplasma gondii infection in gerbils. Ank. Üniv. Vet. Fak. Derg. 2015, 62, 165–170.
- Al-Kayiem, A.H.H.; Ibrahim, M.A. The influence of the equivalent hydraulic diameter on the pressure drop prediction of annular test section. IOP Conf. Ser.: Mater. Sci. Eng. 2015, 100, 012049.
- Pietarinen-Runtti, P.; Lakari, E.; Raivio, K.O.; Kinnula, V.L. Expression of antioxidant enzymes in human inflammatory cells. Am. J. Physiol. Cell Physiol. 2000, 278, C118–C125.
- Stinefelt, B.; Leonard, S.S.; Blemings, K.P.; Shi, X.; Klandorf, H. Free radical scavenging, DNA protection, and inhibition of lipid peroxidation mediated by uric acid. Ann. Clin. Lab. Sci. 2005, 35, 37–45.
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12.
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74.
- Kiran, T.; Karaman, U.; Arici, Y.; Yildiz, S. Comparison of malondialdehyde, nitric oxide, adenosine deaminase and glutathione levels in patients with Entamoeba coli, Enterobius vermicularis, Giardia intestinalis, Demodex spp. positive, hydatid cyst and Toxoplasma gondii serum positive. Ann. Med. Res. 2019, 26, 1420.
- Al-Azzauy, A.A.M. Evaluation of Erythrocyte Malondialdehyde, Glutathione Concentration and Serum Nitric Oxide Levels in Patients with Toxoplasma gondii. Ibn AL-Haitham J. Pure Appl. Sci. 2011, 24, 35–40.
- McCall, M.R.; Frei, B. Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radic. Biol. Med. 1999, 26, 1034–1053.
- Khaleel, F.M.; Hameed, A.S.; Dawood, A.S. Evaluation of antioxidant (GSH, Vitamin A, E, C) and MDA in Iraqi women with toxoplasmosis. Indian J. Forensic Med. Toxicol. 2020, 14, 1446–1449.
- Van De Crommenacker, J.; Richardson, D.S.; Koltz, A.M.; Hutchings, K.; Komdeur, J. Parasitic infection and oxidative status are associated and vary with breeding activity in the Seychelles warbler. Proc. R. Soc. B Biol. Sci. 2011, 279, 1466–1476.
- Zhuang, H.; Yao, C.; Zhao, X.; Chen, X.; Yang, Y.; Huang, S.; Pan, L.; Du, A.; Yang, Y. DNA double-strand breaks in the Toxoplasma gondii-infected cells by the action of reactive oxygen species. Parasites Vectors 2020, 13, 1–9.
- Yazar, S.; Kilic, E.; Saraymen, R.; Sahin, I. MDA Levels in Toxoplasma seropositive patients. Ann. Saudi Med. 2003, 23, 413–415.
- Karaman, U.; Çelik, T.; Kiran, T.R.; Colak, C.; Daldal, N.U. Malondialdehyde, Glutathione and Nitric Oxide Levels in Toxoplasma gondii Seropositive Patients. Korean J. Parasitol. 2008, 46, 293–295.
- Xue, J.; Jiang, W.; Chen, Y.; Gong, F.; Wang, M.; Zeng, P.; Xia, C.; Wang, Q.; Huang, K. Thioredoxin reductase from Toxoplasma gondii: An essential virulence effector with antioxidant function. FASEB J. 2017, 31, 4447–4457.
- Charvat, R.; Arrizabalaga, G. Oxidative stress generated during monensin treatment contributes to altered Toxoplasma gondii mitochondrial function. Sci. Rep. 2016, 6, 22997.
- Akerman, S.E.; Müller, S. Peroxiredoxin-linked Detoxification of Hydroperoxides in Toxoplasma gondii. J. Biol. Chem. 2005, 280, 564–570.
- Deponte, M.; Becker, K. Biochemical characterization of Toxoplasma gondii 1-Cys peroxiredoxin 2 with mechanistic similarities to typical 2-Cys Prx. Mol. Biochem. Parasitol. 2005, 140, 87–96.
- Ding, M.; Clayton, C.; Soldati, D. Toxoplasma gondii catalase: Are there peroxisomes in toxoplasma? J. Cell Sci. 2000, 113, 2409–2419.
- Scandalios, J.G. Oxygen Stress and Superoxide Dismutases. Plant Physiol. 1993, 101, 7–12.
- Amalric, L.; Guillard, C.; Pichat, P. Use of catalase and superoxide dismutase to assess the roles of hydrogen peroxide and superoxide in the TiO2 or ZnO photocatalytic destruction of 1,2-dimethoxybenzene in water. Res. Chem. Intermed. 1994, 20, 579–594.
- DeRocher, A.E.; Coppens, I.; Karnataki, A.; Gilbert, L.A.; Rome, M.E.; Feagin, J.E.; Bradley, P.J.; Parsons, M. A Thioredoxin Family Protein of the Apicoplast Periphery Identifies Abundant Candidate Transport Vesicles in Toxoplasma gondii. Eukaryot. Cell 2008, 7, 1518–1529.
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87.
- Wek, R.C. Role of eIF2α Kinases in Translational Control and Adaptation to Cellular Stress. Cold Spring Harb. Perspect. Biol. 2018, 10, a032870.
- Augusto, L.; Martynowicz, J.; Amin, P.H.; Carlson, K.R.; Wek, R.C.; Sullivan, W.J. TgIF2K-B Is an eIF2α Kinase in Toxoplasma gondii That Responds to Oxidative Stress and Optimizes Pathogenicity. mBio 2021, 12, 1–14.
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26.
- Nazarlu, Z.H.A.; Matini, M.; Bahmanzadeh, M.; Foroughi-Parvar, F. Toxoplasma gondii: A Possible Inducer of Oxidative Stress in Re-productive System of Male Rats. Iran. J. Parasitol. 2020, 15, 521–529.
- Bahrami, S.; Shahriari, A.; Tavalla, M.; Azadmanesh, S.; Hamidinejat, H. Blood Levels of Oxidant/Antioxidant Parameters in Rats Infected with Toxoplasma gondii. Oxidative Med. Cell. Longev. 2016, 2016, 1–6.
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27.
- Machado, V.S.; Bottari, N.B.; Baldissera, M.D.; Rech, V.C.; Ianiski, F.R.; Signor, C.; Rubin, M.A.; Waczuk, E.P.; Schwertz, C.I.; Mendes, R.E.; et al. Diphenyl diselenide supplementation in infected mice by Toxoplasma gondii: Protective effect on behavior, neuromodulation and oxidative stress caused by disease. Exp. Parasitol. 2016, 169, 51–58.
- Delavari, I.; Khadjeh, G.; Bahrami, S.; Jalali, S.M.; Esmaeelzadeh, S. Evaluation of local tissue oxidative stress and its possible involvement in the pathogenesis of toxoplasmosis in rats experimentally infected with Toxoplasma gondii. Trop. Biomed. 2017, 34, 708–716.
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health Lien. Int. J. Biomed. Sci. Rev. 2008, 4, 89–96.
- Faria, J.; Couto, C.; Wierzynski, S.; Bottari, N.; Baldissera, M.; Pereira, W.; Da Silva, A. Feline Toxoplasmosis: Tumor Necrosis Factor, Nitric Oxide, and Free Radicals in Seropositive Cats. J. Parasitol. 2018, 104, 86–88.
- Dahlgren, C.; Karlsson, A. Respiratory burst in human neutrophils. J. Immunol. Methods 1999, 232, 3–14.
- James, S.L.; Nacy, C. Effector functions of activated macrophages against parasites. Curr. Opin. Immunol. 1993, 5, 518–523.
- Bedard, K.; Krause, K.H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313.
- Matta, S.K.; Patten, K.; Wang, Q.; Kim, B.-H.; MacMicking, J.D.; Sibley, L.D. NADPH Oxidase and Guanylate Binding Protein 5 Restrict Survival of Avirulent Type III Strains of Toxoplasma gondii in Naive Macrophages. mBio 2018, 9, e01393-18.
- James, S.L. Role of nitric oxide in parasitic infections. Microbiol. Rev. 1995, 59, 533–547.
- Tonin, A.; Weber, A.; Ribeiro, A.; Camillo, G.; Vogel, F.; Moura, A.; Bochi, G.; Moresco, R.; Da Silva, A. Serum levels of nitric oxide and protein oxidation in goats seropositive for Toxoplasma gondii and Neospora caninum. Comp. Immunol. Microbiol. Infect. Dis. 2015, 41, 55–58.
- Miller, C.; Smith, N.; Johnson, A. Cytokines, Nitric Oxide, Heat Shock Proteins and Virulence in Toxoplasma. Parasitol. Today 1999, 15, 418–422.
- Türkoğlu, Ş.A.; Yaman, K.; Orallar, H.; Camsari, C.; Karabörk, Ş.; Ayaz, E. Acute toxoplasmosis and antioxidant levels in the liver, kidney and brain of rats. Ann. Parasitol. 2018, 64, 241–247.
- Elsheikha, H.M.; El-Motayam, M.H.; Abouel-Nour, M.F.; Morsy, A.T.A. Oxidative stress and immune-suppression in Toxoplasma gondii positive blood donors: Implications for safe blood transfusion. J. Egypt. Soc. Parasitol. 2009, 39, 421–428.
- Alajmi, R.A.; Al-Megrin, W.A.; Metwally, D.M.; Al-Subaie, H.; Altamrah, N.; Barakat, A.M.; Moneim, A.E.A.; Al-Otaibi, T.T.; El-Khadragy, M.F. Anti-Toxoplasma activity of silver nanoparticles green synthesized with Phoenix dactylifera and Ziziphus spina-christi extracts which inhibits inflammation through liver regulation of cytokines in Balb/c mice. Biosci. Rep. 2019, 39, 1–16.
- Terman, A.; Kurz, T.; Gustafsson, B.; Brunk, U.T. Lysosomal labilization. IUBMB Life 2006, 58, 531–539.
- Al-Kuraishy, H.M.; Al-Kuraishi, A.H.; Al-Windy, S.; Al-Gareeb, A.I. Toxoplasmosis and Risk of Endothelial Dysfunction: Role of Oxidative Stress and Pro-Inflammatory Mediators. Arch. Clin. Infect. Dis. 2020, 14, 6–11.
- Meneceur, P.; Bouldouyre, M.-A.; Aubert, D.; Villena, I.; Menotti, J.; Sauvage, V.; Garin, J.-F.; DeRouin, F. In Vitro Susceptibility of Various Genotypic Strains of Toxoplasma gondii to Pyrimethamine, Sulfadiazine, and Atovaquone. Antimicrob. Agents Chemother. 2008, 52, 1269–1277.
- Alday, P.H.; Doggett, J.S. Drugs in development for toxoplasmosis: Advances, challenges, and current status. Drug Des. Dev. Ther. 2017, 11, 273–293.
- Capasso, C.; Supuran, C.T. Sulfa and trimethoprim-like drugs—Antimetabolites acting as carbonic anhydrase, dihydropteroate synthase and dihydrofolate reductase inhibitors. J. Enzym. Inhib. Med. Chem. 2014, 29, 379–387.
- Anacleto-Santos, J.; López-Camacho, P.; Mondragón-Flores, R.; Vega-Ávila, E.; Islas, G.B.; Mondragón-Castelán, M.; Carrasco-Ramírez, E.; Rivera-Fernández, N. Anti-toxoplasma, antioxidant and cytotoxic activities of Pleopeltis crassinervata (Fée) T. Moore hexane fraction. Saudi J. Biol. Sci. 2020, 27, 812–819.
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Shahdin, S.; Daryani, A. Activities of anti-Toxoplasma drugs and compounds against tissue cysts in the last three decades (1987 to 2017), a systematic review. Parasitol. Res. 2018, 117, 3045–3057.
- Smith, N.C.; Goulart, C.; Hayward, J.A.; Kupz, A.; Miller, C.M.; van Dooren, G.G. Control of human toxoplasmosis. Int. J. Parasitol. 2021, 51, 95–121.
- Portes, J.A.; Souza, T.G.; Dos Santos, T.A.T.; Da Silva, L.L.R.; Ribeiro, T.P.; Pereira, M.D.; Horn, A.; Fernandes, C.; DaMatta, R.A.; De Souza, W.; et al. Reduction of Toxoplasma gondii Development Due to Inhibition of Parasite Antioxidant Enzymes by a Dinuclear Iron(III) Compound. Antimicrob. Agents Chemother. 2015, 59, 7374–7386.
- Andrade, R.M.; Reed, S.L. New drug target in protozoan parasites: The role of thioredoxin reductase. Front. Microbiol. 2015, 6, 975.
- Andrade, R.M.; Chaparro, J.D.; Capparelli, E.; Reed, S.L. Auranofin Is Highly Efficacious against Toxoplasma gondii In Vitro and in an In Vivo Experimental Model of Acute Toxoplasmosis. PLoS Negl. Trop. Dis. 2014, 8, e2973.
- Zhai, B.; He, J.-J.; Elsheikha, H.M.; Li, J.-X.; Zhu, X.-Q.; Yang, X. Transcriptional changes in Toxoplasma gondii in response to treatment with monensin. Parasites Vectors 2020, 13, 1–11.
- Murata, Y.; Sugi, T.; Weiss, L.M.; Kato, K. Identification of compounds that suppress Toxoplasma gondii tachyzoites and bradyzoites. PLoS ONE 2017, 12, e0178203.
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Kato, K. Inorganic nanoparticles kill Toxoplasma gondii via changes in redox status and mitochondrial membrane potential. Int. J. Nanomed. 2017, 12, 1647–1661.
- Adeyemi, O.S.; Faniyan, T.O. Antioxidant status of rats administered silver nanoparticles orally. J. Taibah Univ. Med. Sci. 2014, 9, 182–186.
- Hudecová, A.; Kusznierewicz, B.; Rundén-Pran, E.; Magdolenová, Z.; Hašplová, K.; Rinna, A.; Fjellsbø, L.M.; Kruszewski, M.; Lankoff, A.; Sandberg, W.J.; et al. Silver nanoparticles induce premutagenic DNA oxidation that can be prevented by phytochemicals from Gentiana asclepiadea. Mutagenesis 2012, 27, 759–769.
- Pan, W.-H.; Xu, X.-Y.; Shi, N.; Tsang, S.W.; Zhang, H.-J. Antimalarial Activity of Plant Metabolites. Int. J. Mol. Sci. 2018, 19, 1382.
More
Information
Subjects:
Medicine, General & Internal
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.9K
Revisions:
2 times
(View History)
Update Date:
17 Jun 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No