 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jean-luc Teddy,gerard Wautier | + 1455 word(s) | 1455 | 2021-06-15 07:43:17 | | | |
| 2 | Bruce Ren | Meta information modification | 1455 | 2021-06-15 11:08:34 | | | | |
| 3 | Conner Chen | Meta information modification | 1455 | 2021-09-22 03:28:22 | | |
Video Upload Options
Inflammation was already described in the antique Egyptian medicine. Several steps have been made in the understanding of the phenomenon since the discovery of blood circulation in the 17th century, then of endothelial cell functions in the second part of the 20th century. The research studies on endothelial cells in vascular disorders associated to atheroma, diabetes mellitus and infectious diseases opened new possibilities in terms of treatment and prevention. The recent SARS-CoV-2 pandemy demonstrated the pivotal role of endothelial cells in thrombotic and hemorrhagic complications in infectious diseases. Endothelial cells (ECs) are the most efficient antithrombotic interface, and participate in the immune response to micro bacterial organisms. The predominant surface of exchange, which is the endothelium monolayer, is not a passive exchange barrier but a selective and adaptive frontier. Beside their barrier function, ECs have a determinant role in the thrombotic hemostatic equilibrium, first by heparinoid surface molecule, and then as receptors for coagulation components, von Willebrand factor (VWF), and Factor VIII coagulation factors.
1. Introduction
Blood circulation was described by William Harvey in the 17th century (1628 Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus). Using the microscope, Marcello Malpighi carried out important work on capillaries and microcirculation. This led to his discovery of capillaries in 1661, which was fundamental to our understanding of the vascular system in the brain (1661De pumonibus epistolae II and Borellium). The study of the vessel functions and pathology started to significantly progress in the 19th century.
A new era was opened in 1970 with the culture of vascular cells. Animal endothelial cells and smooth muscle cells were first accessible and cultured in vitro. Robert Furchgott, Louis Ignarro and Ferid Murad received the Nobel Prize in Physiology or Medicine (1998) for their discovery of the function of nitric oxide (NO) as a major regulator of vascular tone [1]. The reticuloendothelial system, associated with mononuclear system, was originally described at the beginning of the 20th century as a system that clears old cells and foreign bodies.
During the 1980s, it was described that specialized endothelial cells (scavenger endothelial cells) phagocyted cell debris and dead cells. In the liver, sinusoidal endothelial cells, but not macrophages (Kupffer cells), were part of the system described by Ludwig Aschoff (Aschoff L. Das reticulo-endothelial system. Ergeb Inn Med Kinderheilkd, 1924).
The development of the culture of human endothelial cells was an essential step for understanding the physiopathology of vascular disease and inflammatory processes [2][3]. The researchers working on endothelial cells organized meetings in which they shared their results. This informal group became the association “Vascular Biology” during the meeting in San Diego (CA, USA) in 1992. Later, several scientists and medical doctors joined this group.
2. Endothelial Cells
Endothelial cells (ECs) are the most efficient antithrombotic interface, and participate in the immune response to micro bacterial organisms. Beside their barrier function, ECs have a determinant role in the thrombotic hemostatic equilibrium, first by heparinoid surface molecule, and then as receptors for coagulation components, von Willebrand factor (VWF), and Factor VIII coagulation factors. A fraction of plasma antithrombin is bonded to the heparan present on endothelial cells. The heparan sulfate insures the non-thrombotic properties of EC [4].
The functions of EC are difficult to explore in vivo since it is a unique linear cell inside the vessel. Fortunately, human ingenuity has made available endothelial cells in culture and facilitated the investigation of their functions.
The method described by Maruyama [5] using human umbilical vein as a source of endothelial cells was subsequently developed by several research groups. The cord is separated from the placenta several hours after birth, placed in a sterile container, as described by EA Jaffe [3]. Human umbilical vein endothelial cells (HUVEC) are harvested from human umbilical vein after collagenase digestion. The cells reach confluence after 8–10 days [6].
Weibel-Palade bodies are storage granules present in endothelial cells. They contain P-selectin and von Willebrand factor, which is essential for hemostasis. P-selectin is an adhesion molecule participating in vascular permeability, and plays a role in leukocyte adhesion and platelet aggregation. Weibel-Palade bodies contain other molecules such as interleukine-8, endothelin-1, eotaxin-3, angiopoietin-2 and fructosyltransferase.
Several attempts have been made to establish a continuous human endothelial cell line [7][8]. (SV40) transfection has been proposed by several authors as a possible way of obtaining a permanent endothelial cell line [9][10]. An immortalized vascular endothelial cell line (IVEC) was obtained from human umbilical vein endothelial cells, which were transformed by intranuclear microinjection of a recombinant gene composed of the large T-encoding DNA of SV40 driven by the truncated vimentin promoter (HuVim 830-T/t). IVEC is a reliable and convenient cell line for studying endothelial cell reactivity in the presence of various stimuli.
Since we observed unexpected results with two different cell lines of Human Microvascular Endothelial Cells (HMEC), as reported by Xu Y. [11], we tested these two types of human microvascular endothelial cells (HMEC-1, HMVEC-d) in parallel. The spontaneous proliferation of HMEC-1 measured by [H3-methyl] thymidine incorporation was higher compared to HUVEC.
Endothelial cells obtained from arteriovenous malformations called AMEC [12] in culture demonstrated a higher propensity to proliferate and a reduced reactivity to antiproliferative compounds such as Interferon γ.
3. Vascular Permeability and Endothelial Cell Junctions
In the vessels, cell–cell junctions connect and modulate the activities of the individual endothelial cells, specifically for the control of vascular permeability and immune cell trafficking [13]. The cell–cell junctions in the endothelium are defined as tight junctions and adherens junctions, which alternate. The binding of vascular endothelial cadherin (VE-cadherin) to the same type of molecule is essential for the organization of the vascular network. VE-cadherin can also form complexes with signaling systems, like the endothelial specific VEGF receptor-2 (VEGFR-2), the vascular endothelial phosphotyrosine phosphatase, the transforming growth factor β receptor complex, and also the fibroblast growth factor receptor-1.
G-protein-coupled receptors can induce VEGFR-2. Gα13 is a member of the G protein family, which transduces signals from G-protein-coupled receptors through interactions with downstream effectors [14].
4. Endothelial Cell Receptors in Inflammation
Before the availability of endothelial cells (ECs) in culture, the interaction between blood cells and endothelial cells was investigated by means of transcapillary microscopy in animal or human eyes [15].
Observing the vessels in frogs, Ilya Mechnikov, called also Elie Metchnikoff [16][17], described phagocytosis, one of the main functions of leukocytes in the battle against infection. The characterization of adhesion molecules on endothelial cells provided insight into the pathophysiology of inflammation and atherosclerosis.
This first step of leukocyte adhesion is then followed by interactions between members of the CD18 family and their respective ligands, which mediate firm adhesion/arrest of leukocytes on ECs. The leukocyte adhesion mediated by P-selectin can occur when inflammatory mediators such as histamine or thrombin activate ECs. The expression of E-selectin on the endothelial cell surface is indeed induced only by stimulation of ECs by inflammatory cytokines (e.g., IL-1, TNF-α). The intercellular cell adhesion molecules (ICAM) are also major adhesion molecules that arrest circulating leukocytes on ECs [18][19][20][21][22][23][24][25][26][27].
Monocytes and polymorphonuclear neutrophils in inflammatory conditions are associated with venous thrombosis. Monocytes secrete tumor necrosis factor α (TNF α). In vitro TNF α increased the shedding of Protein C inhibiting thrombomodulin production and stimulated tissue factor synthesis by endothelial cells and monocyte-macrophages (Figure 1).
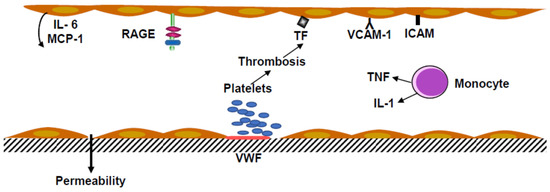
Figure 1. Inflammation and thrombosis in the vascular endothelium. Platelets adhering to the subendothelium, including von Willebrand factor (VWF) recruiting circulating platelets to form an aggregate. Leukocyte integrins bind to Vascular Cell Adhesion Molecule-1 (VCAM-1) and Intercellular Adhesion Molecule (ICAM). Activation of the receptor RAGE, a multiligand receptor, induces the release of Macrophage Chemoattractant Protein-1 (MCP-1) and Interleukin-6 (IL-6), and the production of Tissue Factor (TF). Monocytes stimulated by bacteria, endotoxins and viruses release tumor necrosis factor (TNF) and interleukin-1 (IL-1).
5. Physiology and pathophysiology of human vascular cells
The multiligand receptor named RAGE appeared involved in the pathophysiology of several disorders: diabetes mellitus, Alzheimer disease. RAGE not only binds advanced glycation end products (AGE), but also other macromolecules, including members of the S100 family, amyloid-ß-peptide and HMGB1, a nuclear protein, released upon cell damage. All ligands of RAGE have structural features characterized by an alpha-helix and ß-sheet. The latter line of thought prompted attempts to develop RAGE inhibitors for treating atherosclerosis, diabetic complications, septicemia and other diseases.
Glycation of proteins has deleterious effects in the pathogenesis of diabetic cataract. Retinal lesions are the major ocular complications in diabetes mellitus. Retinal micro vessels are altered as a consequence of the selective loss of intramural pericytes. Pyridoxamine, an inhibitor of AGE formation and lipoxidation end products, limits diabetes-induced retinal vascular lesions. On other hand, benfotiamine inhibits hyperglycemia-dependent pathways and NFκB activation, and prevents experimental diabetic retinopathy. The incubation of human endothelial cells with specific AGE (carboxymethyl-lysine-modified adducts) stimulated intracellular generation of hydrogen peroxide. Retinal vein occlusion (RVO) is a common cause of permanent visual loss and is the fifth cause of unilateral blindness.
References
- Furchgott, R.F. The 1996 Albert Lasker Medical Research Awards. The discovery of endothelium-derived relaxing factor and its importance in the identification of nitric oxide. JAMA 1996, 276, 1186–1188.
- Gimbrone, M.A.; Cotran, R.S.; Folkman, J. Human vascular endothelial cells in culture. J. Cell Biol. 1974, 60, 673–684.
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of Human Endothelial Cells Derived from Umbilical Veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745–2756.
- Rosenberg, R.D. Biochemistry of heparin antithrombin interactions, and the physiologic role of this natural anticoagulant mechanism. Am. J. Med. 1989, 87, S2–S9.
- Maruyama, Y. The human endothelial cell in tissue culture. Cell Tissue Res. 1963, 60, 69–79.
- Wautier, J.-L.; Paton, R.C.; Wautier, M.-P.; Pintigny, D.; Abadie, E.; Passa, P.; Caen, J.P. Increased Adhesion of Erythrocytes to Endothelial Cells in Diabetes Mellitus and Its Relation to Vascular Complications. N. Engl. J. Med. 1981, 305, 237–242.
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737.
- Faller, D.V.; Kourembanas, S.; Ginsberg, D.; Hannan, R.; Collins, T.; Ewenstein, B.M.; Pober, J.S.; Tantravahi, R. Immortalization of human endothelial cells by murine sarcoma viruses, without morphologic transformation. J. Cell. Physiol. 1988, 134, 47–56.
- Gimbrone, M.A.; Fareed, G.C. Transformation of cultured human vascular endothelium by SV40 DNA. Cell 1976, 9, 685–693.
- Ide, H.; Minamishima, Y.; Eizuru, Y.; Okada, M.; Sakihama, K.; Katsuki, T. “Transformation” of human endothelial cells by SV40 virions. Microbiol. Immunol. 1988, 32, 45–55.
- Xu, Y.; Swerlick, R.; Sepp, N.; Bosse, D.; Ades, E.W.; Lawley, T.J. Characterization of Expression and Modulation of Cell Adhesion Molecules on an Immortalized Human Dermal Microvascular Endothelial Cell Line (HMEC-1). J. Investig. Dermatol. 1994, 102, 833–837.
- Wautier, M.P.; Boval, B.; Chappey, O.; Enjolras, O.; Wernert, N.; Merland, J.J.; Wautier, J.L. Cultured endothelial cells from human arteriovenous malformations have defective growth regulation. Blood 1999, 94, 2020–2028.
- Lampugnani, M.G.; Dejana, E.; Giampietro, C. Vascular Endothelial (VE)-Cadherin, Endothelial Adherens Junctions, and Vascular Disease. Cold Spring Harb. Perspect. Biol. 2018, 10, a029322.
- Kelly, E.; Bailey, C.P.; Henderson, G. Agonist-selective mechanisms of GPCR desensitization. Br. J. Pharmacol. 2008, 153, S379–S388.
- Simionescu, N. Cellular aspects of transcapillary exchange. Physiol. Rev. 1983, 63, 1536–1579.
- Silverstein, A.M. Editorial: Ilya Metchnikoff, the phagocytic theory, and how things often work in science. J. Leukoc. Biol. 2011, 90, 409–410.
- Underhill, D.M.; Gordon, S.; Imhof, B.A.; Núñez, G.; Bousso, P. Élie Metchnikoff (1845–1916): Celebrating 100 years of cellular immunology and beyond. Nat. Rev. Immunol. 2016, 16, 651–656.
- Wautier, J.-L.; Wautier, M.-P. Cellular and Molecular Aspects of Blood Cell–Endothelium Interactions in Vascular Disorders. Int. J. Mol. Sci. 2020, 21, 5315.
- Rainger, G.; Chimen, M.; Harrison, M.J.; Yates, C.M.; Harrison, P.; Watson, S.P. The role of platelets in the recruitment of leukocytes during vascular disease. Platelets 2015, 26, 507–520.
- Hansson, G.K.; Libby, P. The immune response in atherosclerosis: A double-edged sword. Nat. Rev. Immunol. 2006, 6, 508–519.
- Libby, P. Mechanisms of Acute Coronary Syndromes and Their Implications for Therapy. N. Engl. J. Med. 2013, 368, 2004–2013.
- Wautier, J.-L.; Setiadi, H.; Vilette, D.; Weill, D.; Wautier, M.-P. Leukocyte adhesion to endothelial cells. Biorheology 1990, 27, 425–432.
- Barbe, L.L.; Boval, B.M.; Wautier, M.P.; Wautier, J.L. An in vitro system for testing leucocyte and leukaemic cell line adhesion to synthetic fibres. Br. J. Haematol. 2001, 115, 664–671.
- Liote, F.; Boval-Boizard, B.; Weill, D.; Kuntz, D.; Wautier, J.-L. Blood monocyte activation in rheumatoid arthritis: Increased monocyte adhesiveness, integrin expression, and cytokine release. Clin. Exp. Immunol. 1996, 106, 13–19.
- Nash, G.B. Adhesion Between Platelets and Leukocytes or Endothelial Cells. Platelets Megakaryocytes 2004, 272, 199–214.
- Setiadi, H.; McEver, R.P. Clustering endothelial E-selectin in clathrin-coated pits and lipid rafts enhances leukocyte adhesion under flow. Blood 2008, 111, 1989–1998.
- Jones, D.P.; True, H.D.; Patel, J. Leukocyte Trafficking in Cardiovascular Disease: Insights from Experimental Models. Mediat. Inflamm. 2017, 2017, 1–9.


