Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Dickson Kofi Wiredu Ocansey | + 2955 word(s) | 2955 | 2021-05-19 11:59:17 | | | |
| 2 | Vivi Li | Meta information modification | 2955 | 2021-05-26 10:10:51 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ocansey, D.K.W. Stem Cell-Derived Extracellular Vesicles. Encyclopedia. Available online: https://encyclopedia.pub/entry/10088 (accessed on 29 June 2026).
Ocansey DKW. Stem Cell-Derived Extracellular Vesicles. Encyclopedia. Available at: https://encyclopedia.pub/entry/10088. Accessed June 29, 2026.
Ocansey, Dickson Kofi Wiredu. "Stem Cell-Derived Extracellular Vesicles" Encyclopedia, https://encyclopedia.pub/entry/10088 (accessed June 29, 2026).
Ocansey, D.K.W. (2021, May 26). Stem Cell-Derived Extracellular Vesicles. In Encyclopedia. https://encyclopedia.pub/entry/10088
Ocansey, Dickson Kofi Wiredu. "Stem Cell-Derived Extracellular Vesicles." Encyclopedia. Web. 26 May, 2021.
Copy Citation
Extracellular vesicles (EVs), which are the main paracrine components of stem cells, mimic the regenerative capacity of these cells. Stem cell-derived EVs (SC-EVs) have been used for the treatment of various forms of tissue injury in preclinical trials through maintenance of their stemness, induction of regenerative phenotypes, apoptosis inhibition, and immune regulation. The efficiency of SC-EVs may be enhanced by selecting the appropriate EV-producing cells and cell phenotypes, optimizing cell culture conditions for the production of optimal EVs, and further engineering the EVs produced to transport therapeutic and targeting molecules.
stem cell
extracellular vesicle
tissue damage
regenerative medicine
1. Introduction
Extracellular vesicles (EVs) are vesicular entities with lipid bilayer membranes. They were initially defined as “platelet dust” in 1967 [1]. Intense research regarding EVs in the past half century has enabled a thorough understanding of the origin and biological function of EVs and has positioned EVs on the front line of treatments for various diseases.
EVs exist in all bodily fluids and are produced by all types of cells. Smaller vesicles, known as “exosomes” (EXs), are released from cells through the multivesicular endosomal pathway. Larger vesicles, known as “microvesicles” (MVs), are formed by cell membrane budding and apoptotic bodies are produced by the blebbing of aging or dying cells [2][3]. Apoptotic bodies have been studied less often; thus, EXs and MVs are mainly discussed in this article. EVs can mediate cellular waste degradation and interact with recipient cells through surface receptor binding, endosomal uptake, membrane fusion, membrane protein translocation, and by shuttling RNAs and proteins through vesicle cell channels [2].
EVs carry components of EV-producing cells. They have been shown to exert similar pathophysiological/regenerative effects on tissue and cellular functions when they are applied to experimental animal models. Stem cells are the most common EV-producing cells. Stem cells can be isolated successfully from bone marrow, fat, umbilical cords, embryos, and other tissues. Stem cells can differentiate into many types of cells and they can substitute for injured tissues and aid the repair process through their paracrine mechanism at the injury location. Stem cells have been used successfully in the treatment of hematological malignancies, graft-versus-host disease, acute thrombocytopenia, and autoimmune diseases in several experimental in vivo studies [4][5]. However, large-scale production, storage, immune rejection, gene mutation, and tumorigenesis or tumor promotion in vivo limit its application. Stem cell derived-EVs (SC-EVs), as the main paracrine executor, overcome most limitations of stem cell applications. SC-EVs have allowed major advances in preclinical and clinical studies.
2. Stem Cell-Derived EVs in the Treatment of Damaged Tissue
Numerous preclinical trials have reported that SC-EVs can carry active molecules, such as proteins, lipids, and nucleic acids, and exhibit good therapeutic effect against various diseases regarding different systems, including the nervous system, respiratory system, circulatory system, digestive system, and urinary system, among others.
2.1. Neurological System
Brain trauma is a common event that can cause nerve damage and disability. EXs derived from human adipose mesenchymal stem cells (AdMSC-EXs) can significantly increase the number of neurons, reduce inflammation, improve sensory and cognitive function, and produce better effects than AdMSCs alone in rats with traumatic brain injury (TBI) [6]. Kim et al. indicated that systemic administration of CD63+CD81+ EVs produced by human bone marrow-derived stem cells (BMSC-EVs) decreased neuroinflammation 12 h after a TBI in a mouse model of TBI induced by a controlled cortical impact device [7]. They also found that BMSC-EV infusion preserved the pattern separation and spatial learning abilities of mice, which were demonstrated respectively by an object-based behavioral test and a water maze test [7].
Stroke is the sudden rupture or occlusion of cerebral blood vessels that interrupts the blood supply. It is the main cause of death and disability in Chinese adults. Preclinical studies have shown that SC-EVs seem to be a promising candidate for stroke treatment. Xin et al. showed that infusion of BMSC-EXs enhanced oligodendrogenesis and neurogenesis, remodeled synapses, reduced the incidence of stroke, and accelerated the recovery of neurological functions in a rat model of stroke induced by transient middle cerebral artery occlusion [8]. Webb et al. tested the effect of SC-EVs on stroke in a translational large animal model. In their study, they utilized human neural stem cell-derived EVs (NSC-EVs) to treat ischemic stroke that was manufactured by permanent middle cerebral artery occlusion in pigs, and they found that NSC-EVs eliminated the symptoms of intracranial hemorrhage, decreased the cerebral lesion volume and brain swelling, and preserved the white matter integrity compared to the control pigs [9]. They also indicated that NSC-EV treatment improved behavior and mobility in this model [9]. In an ongoing clinical trial, MSC-EXs were engineered to overexpress miR-124 for the treatment of ischemic stroke (NCT03384433; the estimated primary completion date is April 17, 2020 and the estimated study completion is December 17, 2021).
Alzheimer’s disease (AD) is a common disease that usually occurs in older people. There is no effective treatment for AD. Aβ protein accumulation in nerve cells may promote neurodegeneration and memory impairment in AD. Studies have shown that AdMSC-EXs transport Aβ protease, which effectively inhibits Aβ protein accumulation in nerve cells in vitro, suggesting a possible treatment application for AD [10]. In addition, Cui et al. indicated that BMSC-EX administration ameliorated animal cognitive function and symptoms in a mouse model of AD [11]. In addition, EXs derived from hypoxic-preconditioning BMSCs displayed enhanced therapeutic effects study [11].
The high incidence of spinal cord injury (SCI), which has occurred in recent years, often results in serious sequelae, such as lower limb paralysis and incontinence, for which effective treatments are currently lacking. Liu et al. showed that BMSC-EXs could target specific sites of SCI and promote functional recovery in a rat model of SCI by inhibiting neuronal apoptosis and increasing neuroinflammatory improvement, angiogenesis, and the inactivation of A1 astrocytes [12]. Additionally, Ruppert et al. found that the administration of human BMSC-EVs displayed reduced inflammatory responses, improved motor function, and enhanced mechanical sensitivity thresholds in a rat model of SCI [13]. Recently, a study demonstrated that EXs from human umbilical cord MSCs (hucMSC-EXs) promoted functional recovery in mice with SCI by reducing inflammation [14].
2.2. Cardiovascular System
Cardiovascular diseases seriously threaten human health because they have high incidence and mortality rates. Improvement of treatment methods remains a worldwide objective. The proliferation ability of cardiomyocytes is normally considerably weak, and their ability to achieve complete repair through self-regulation after an injury is low. Preclinical studies have indicated that SC-EVs have great application prospects in the treatment of myocardial infarction (MI). Adamiak et al. showed that intravenous injection of induced pluripotent stem cell (iPSC)-derived EVs (iPSC-EVs) at 48 h after MI in mice resulted in improved left ventricular function, reduced left ventricular mass, preservation of viable myocardia and reduction in apoptosis in the infarct zone [15]. A similar phenomenon was observed by Khan who reported that embryonic stem cell (ESC)-EX treatment of mouse MI promoted cardiac blood flow recovery, alleviated myocardial fibrosis, reduced the infarct size, and significantly recovered cardiac function [16]. SC-EVs encapsulate microRNA (miRNA), which reinforces cardiac repair by facilitating angiogenesis. Wang et al. demonstrated that miR-210 enriched in BMSC-EVs drives endothelial proliferation and migration in vitro, and it improves cardiac angiogenesis and cardiac function after MI in mice by targeting Ephrin-A3 [17]. Zhu et al. showed that BMSC hypoxic pretreatment increased the enrichment of angiogenesis-associated miRNAs in BMSC-EXs, which promoted angiogenesis more effectively and improved the therapeutic effects of MI treatment in mice [18].
Preclinical studies have indicated that BMSC-EXs could alleviate inflammatory responses in cardiovascular diseases. Macrophages and neutrophils are activated and recruited to the injured site after a MI and they release a large number of inflammatory factors that trigger a series of inflammatory reactions. In mouse models of doxorubicin-induced dilated cardiomyopathy, intravenous injection of BMSC-EXs improved cardiac function, inhibited cardiac dilation, alleviated cardiomyocyte apoptosis, and reduced the expression of inflammatory factors in addition to the number of proinflammatory macrophages at the infiltration site and in the blood [19].
2.3. Respiratory System
Potter et al. showed human BMSC-EVs could significantly reduce pulmonary vascular permeability that was caused by hemorrhagic shock in mice via regulation of cytoskeletal signaling [20]. In another study, Tang et al. showed that BMSC-MVs could promote the stability of the pulmonary vascular structure and improve inflammation in the lungs by delivering angiopoietin-1 (Ang-1) messenger RNAs (mRNAs) in mice. In their research, Tang et al. showed that unmodified BMSC-MVs, but not Ang-1 mRNA-deficient MVs, were able to maintain the integrity of endotoxin-stimulated microvascular endothelial cells in vitro, and they could reduce lung inflammation in a mouse model of lipopolysaccharide (LPS)-induced acute lung injury [21]. Vascular endothelial growth factor (VEGF) can mediate the therapeutic effects of EVs originating from human umbilical cord blood-derived MSCs (hUCB-MSC-EVs) after neonatal hyperoxia-induced lung injury in rats. Ahn et al. demonstrated that treatment with 20 μg of hUCB-MSC-EVs improved lung injury, protected blood vessels, and maintained normal function of the alveoli in newborn rats, whereas VEGF-deficient MSC-MVs functioned poorly [22].
Khatr et al. reported swine BMSC-EVs could disturb the agglutination reaction of various influenza viruses, restrict influenza virus replication in lung epithelial cells in vitro, and alleviate virus-induced apoptosis [23]. The authors suggested that intratracheal administration of BMSC-EVs reduced viral shedding, inhibited lung influenza virus replication, and reduced the production of proinflammatory cytokines in a pig influenza virus model [23]. Additionally, Chaubey demonstrated that intraperitoneal injection of hucMSC-EXs could improve experimental bronchopulmonary dysplasia in mice [24].
2.4. Liver
Preclinical studies have demonstrated that SC-EVs harbor the potential to treat liver diseases through the delivery of various active molecules. Haga et al. demonstrated that BMSC-EVs protected against hepatic ischemia/reperfusion injury (IRI) in a mouse model [25]. Similarly, Tamura et al. discovered murine BMSC-EXs improved chemical-induced hepatocyte injury and promoted hepatocyte regeneration in mice, which was mediated through immunosuppression and immune protection [26]. In addition, Rigo et al. discovered human liver SC-EVs effectively reduced liver injury in an ex vivo normothermic hypoxic rat liver perfusion model [27]. Normothermic machine perfusion is an emerging approach for liver preservation before transplantation and it may induce hypoxic injury. Rigo and colleagues indicated that liver SC-EVs reduced the level of alanine aminotransferase and lactate dehydrogenase in perfusate samples, and they protected histological damage and apoptosis in damaged livers [27].
Liver disease often includes liver fibrosis. Our studies have revealed that hucMSC-EXs can relieve liver fibrosis in mice by inactivating transforming growth factor (TGF)-β/Smad signaling, reducing collagen deposition, and alleviating inflammation [28]. Qu et al. also suggested that BMSC-EXs effectively deliver miRNA-181-5p to damaged hepatocyte sites and they prevented liver fibrosis in a mouse model by activating autophagy [29].
Our study reported that hucMSC-EXs carrying glutathione peroxidase1 protect against liver failure in mice by reducing inflammation and oxidative stress [30]. In a murine model of liver failure that was induced by d-galactosamine/tumor necrosis factor-α (TNF-α), Haga et al. found that BMSC-EV administration reduced liver damage and regulated the inflammatory response [31]. In this study, imaging of fluorescence-labeled BMSC-EVs suggested that EVs preferentially migrate to the livers of mice, and their density increases after chemically induced damage. They further demonstrated that EVs from murine or human BMSCs significantly increased the survival rates (murine BMSC-MVs: 57.1%; human BMSC-MVs: 37.5%; Control: 0%) of mice with liver failure, which were superior to those of BMSCs [31]. They demonstrated that Y-RNA-1 is enriched within BMSC-EVs and Y-RNA-1 mediates the protective effects of BMSC-EVs on hepatocyte apoptosis [31].
3. Stem Cell Culture for Extracellular Vesicle Production
The contents and functions of EVs produced by diverse stem cells vary widely. Some EVs secreted by the same type of stem cells under various culture conditions differ in content and function. The selection of appropriate parental stem cells that are cultured under a specific condition for large-scale production of safer and more effective EVs is essential for the development of new treatments for diseases in the future.
3.1. Stem Cell Selection
Stem cells can be successfully isolated from bone marrow, fat, umbilical cords, embryos, placentas, amniotic fluid, blood, livers, skin, and other tissues. The first four types of stem cells listed here based on their origins are currently the most widely studied.
BMSCs exert many biological functions, and their earliest application was for leukemia treatment [32]. Preclinical studies have confirmed that BMSC-EVs can be used to treat various diseases, as expressed in this review. However, BMSC acquirement is difficult and invasive, and BMSCs have poor ability for in vitro expansion, which is not conducive to large-scale production of EVs for clinical use. AdMSCs are easy to obtain and proliferate, and autologous AdMSC transplantation can prevent immune rejection and provide beneficial effects. However, AdMSCs carry the risk of tumor promotion [33]. Therefore, clarifying the effects of AdMSC-EVs on tumors before attempts at clinical application is essential. ESCs are difficult to obtain and utilize for ethical reasons, and they contain the risk of tumorigenesis in the body, which also limits their clinical applications. hucMSCs are easy to acquire and are not associated with ethical controversies. They have strong expansion ability and low immunogenicity. Preclinical studies have reported that hucMSC-EVs have good therapeutic effects in tissue repair and regeneration, as expressed in this review, and hucMSCs have an inhibitory effect on some tumors [33][34]. Currently, commercial cell banks that collect and store hucMSCs are emerging. These advantages enable large-scale production of SC-EVs for clinical applications and iPSC culture makes large-scale production of stem cells easier. Numerous preclinical investigations have demonstrated that iPSC-EVs have similar therapeutic effects to iPSCs in repairing damaged tissue. Therefore, iPSC culture is an alternative for the large-scale production of EVs.
3.2. Cell Culture
Different culture conditions result in different functions and phenotypes of stem cells, which then affect the content and function of EVs. Studies have suggested that various conditions, including cell density, cell phenotype, hypoxia, drug preconditioning, inflammatory stimuli, and 3D culture, influence the properties and activities of stem cells (Figure 1).
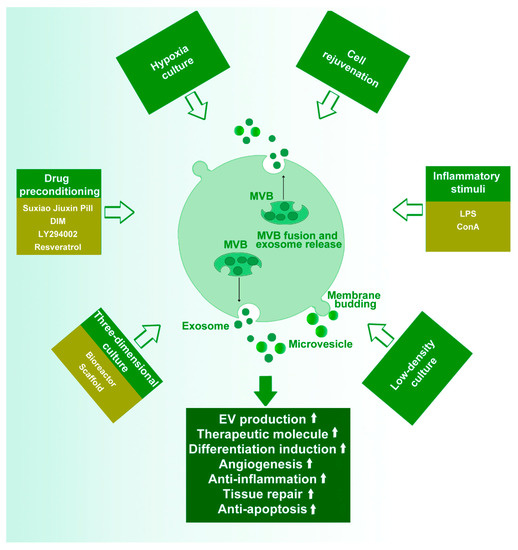
Figure 1. The effect of various culture conditions or treatment on the stem cell-derived EVs. Cell density, cell phenotype, hypoxia treatment, inflammatory stimuli, drug preconditioning, and three-dimensional (3D) culture influence the production and capabilities of extracellular vesicles (EVs). LPS, lipopolysaccharide; DIM, 3,3′-Diindolylmethane; MVB, multivesicular bodies.
3.2.1. Cell Density
Seeding density may affect EV secretion. EVs are typically collected from culture media at 60%–90% cell confluence. Stem cells cultured at high density demonstrate contact inhibition and are induced to enter a resting state [35]. Low-density inoculation (1.5 cells/cm2) may result in faster stem cell proliferation [36]. Low-density cultures may activate EV-mediated paracrine signaling of stem cells and promote EV secretion [37].
3.2.2. Cell Phenotypes
Cell phenotypes affect the components and functions of EVs. Stem cells alter phenotypes and functions as the in vitro expansion time increases. Early MSCs have the potential for osteogenic and adipogenic differentiation, and the differentiation potential tilts toward adipogenesis as the in vitro culture is extended [37]. MSCs secrete less EVs after aging [37]. Kulkarni et al. revealed that MVs produced from young BMSCs are rich in autophagy-related gene mRNAs, which better support hematopoietic stem cell transplantation in mice compared to MVs from aging BMSCs [38]. Our unpublished research has indicated that resveratrol treatment improves the senescence phenotype of hucMSCs and increases the differentiation potentials. EXs originating from hucMSCs pretreated with resveratrol had better healing effects on murine lung cancer metastasis than EXs from untreated hucMSCs.
Therefore, improving culture conditions and ensuring stem cells remain as young as possible are necessary, and EVs secreted by younger cells may have superior therapeutic effects on certain diseases. In a different study, Wang et al. noted that EXs produced from MSCs in the late stages of osteogenic differentiation, but not in the early stages, delivered osteogenic differentiation-associated miRNAs to simulate homotypic cell osteogenic differentiation and mineralization in vitro [39]. This provided the hypothesis that stem cells can be induced into a specific tissue phenotype in vitro for collection of their EVs that can be used in corresponding tissue damage repair.
3.2.3. Hypoxia Culture
Cui et al. revealed that EXs produced from hypoxia-preconditioned mouse BMSCs exhibited considerably improved learning and memory capabilities and reduced plaque deposition, neuroinflammation, and Aβ expression in an AD mouse model [11]. Zhu et al. showed that hypoxia (0.5% O2) stimulated exosomal mRNA-210 secretion through nSMase2 in mouse BMSCs to promote angiogenesis and improve cardiac function in a mouse model of MI [18].
3.2.4. Drug Preconditioning and Inflammatory Stimuli
Ruan et al. revealed that Suxiao Jiuxin Pill, a type of traditional Chinese medicine, could promote mouse BMSC-EV secretion [40]. Our study revealed that 3,3′-Diindolylmethane-stimulated hucMSCs secreted Wnt11-overexpressing EVs that could be used to improve wound healing in rats [41]. In addition, Kulkarni et al. noted that aging BMSCs treated with AKT signal inhibitor LY294002 reduced exosomal miR-17 and miR-34a, and both downregulated autophagy-associated mRNA expression in recipient cells [38]. LY294002 treatment increased autophagy-associated genes in BMSC-MVs [38]. MVs from aging BMSCs that were pretreated with LY294002 were more powerful than MVs from young BMSCs in supporting hematopoietic stem transplantation in mice [38]. In another study, EVs from human BMSCs activated by LPS or ConA displayed enhanced anti-inflammatory ability and reduced the release of TNF-α and IFN-γ from spleen cells in vitro [42].
3.2.5. Three-Dimensional Culture
Cell culture configurations for producing EVs include both 2D and 3D systems. The 2D system uses conventional polystyrene flasks for the adhesion growth of EV-producing cells and the 3D system for EV production mainly includes a scaffold-free bioreactor and scaffold-based approach [43]. The 3D system can bestow EV production with higher yields, more natural features, and better therapeutic effects compared to the 2D system [43].
Bioreactors are typically used for large-scale production of EVs. In these devices, EV-producing cells were sown into cylindrical hollow fibers that provided a high surface area that was available to billions of cells. These cells were seeded into the bioreactor and could produce 4-fold more EVs than cells cultured in a traditional 2D flask [44]. EXs from stem cells cultured in a bioreactor protected human dopaminergic neurons from apoptosis in vitro, which was not observed in EXs collected from the 2D culture [45]. In a rat model of experimental TBI, EXs from BMSCs cultured in 3D scaffolds exhibited greater spatial learning ability than EVs harvested from 2D conditions [46].
References
- Hargett, L.A.; Bauer, N.N. On the origin of microparticles: From “platelet dust” to mediators of intercellular communication. Pulm. Circ. 2013, 3, 329–340.
- Wiklander, O.P.B.; Brennan, M.A.; Lotval, J.; Breakefield, X.O.; El Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, eaav8521.
- Andaloussi, S.E.; Mager, I.; Breakefield, X.O.; Wood, M.J. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357.
- Dias, I.E.; Pinto, P.O.; Barros, L.C.; Viegas, C.A.; Dias, I.R.; Carvalho, P.P. Mesenchymal stem cells therapy in companion animals: Useful for immune-mediated diseases? BMC Vet. Res. 2019, 15, 358.
- Abdelrazik, H.; Giordano, E.; Barbanti Brodano, G.; Griffoni, C.; De Falco, E.; Pelagalli, A. Substantial Overview on Mesenchymal Stem Cell Biological and Physical Properties as an Opportunity in Translational Medicine. Int. J. Mol. Sci. 2019, 20, 5386.
- Patel, N.A.; Moss, L.D.; Lee, J.Y.; Tajiri, N.; Acosta, S.; Hudson, C.; Parag, S.; Cooper, D.R.; Borlongan, C.V.; Bickford, P.C. Long noncoding RNA MALAT1 in exosomes drives regenerative function and modulates inflammation-linked networks following traumatic brain injury. J. Neuroinflamm. 2018, 15, 204.
- Kim, D.K.; Nishida, H.; An, S.Y.; Shetty, A.K.; Bartosh, T.J.; Prockop, D.J. Chromatographically isolated CD63+CD81+ extracellular vesicles from mesenchymal stromal cells rescue cognitive impairments after TBI. Proc. Natl. Acad. Sci. USA 2016, 113, 170–175.
- Xin, H.; Katakowski, M.; Wang, F.; Qian, J.Y.; Liu, X.S.; Ali, M.M.; Buller, B.; Zhang, Z.G.; Chopp, M. MicroRNA cluster miR-17-92 Cluster in Exosomes Enhance Neuroplasticity and Functional Recovery After Stroke in Rats. Stroke 2017, 48, 747–753.
- Webb, R.L.; Kaiser, E.E.; Jurgielewicz, B.J.; Spellicy, S.; Scoville, S.L.; Thompson, T.A.; Swetenburg, R.L.; Hess, D.C.; West, F.D.; Stice, S.L. Human Neural Stem Cell Extracellular Vesicles Improve Recovery in a Porcine Model of Ischemic Stroke. Stroke 2018, 49, 1248–1256.
- De Godoy, M.A.; Saraiva, L.M.; de Carvalho, L.R.P.; Vasconcelos-Dos-Santos, A.; Beiral, H.J.V.; Ramos, A.B.; Silva, L.R.P.; Leal, R.B.; Monteiro, V.H.S.; Braga, C.V.; et al. Mesenchymal stem cells and cell-derived extracellular vesicles protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-beta oligomers. J. Biol. Chem. 2018, 293, 1957–1975.
- Cui, G.H.; Wu, J.; Mou, F.F.; Xie, W.H.; Wang, F.B.; Wang, Q.L.; Fang, J.; Xu, Y.W.; Dong, Y.R.; Liu, J.R.; et al. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice. FASEB J. 2018, 32, 654–668.
- Liu, W.; Wang, Y.; Gong, F.; Rong, Y.; Luo, Y.; Tang, P.; Zhou, Z.; Xu, T.; Jiang, T.; Yang, S.; et al. Exosomes Derived from Bone Mesenchymal Stem Cells Repair Traumatic Spinal Cord Injury by Suppressing the Activation of A1 Neurotoxic Reactive Astrocytes. J. Neurotrauma 2019, 36, 469–484.
- Ruppert, K.A.; Nguyen, T.T.; Prabhakara, K.S.; Toledano Furman, N.E.; Srivastava, A.K.; Harting, M.T.; Cox, C.S., Jr.; Olson, S.D. Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles Modify Microglial Response and Improve Clinical Outcomes in Experimental Spinal Cord Injury. Sci. Rep. 2018, 8, 480.
- Sun, G.; Li, G.; Li, D.; Huang, W.; Zhang, R.; Zhang, H.; Duan, Y.; Wang, B. hucMSC derived exosomes promote functional recovery in spinal cord injury mice via attenuating inflammation. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 89, 194–204.
- Adamiak, M.; Cheng, G.; Bobis-Wozowicz, S.; Zhao, L.; Kedracka-Krok, S.; Samanta, A.; Karnas, E.; Xuan, Y.T.; Skupien-Rabian, B.; Chen, X.; et al. Induced Pluripotent Stem Cell (iPSC)-Derived Extracellular Vesicles Are Safer and More Effective for Cardiac Repair Than iPSCs. Circ. Res. 2018, 122, 296–309.
- Khan, M.; Nickoloff, E.; Abramova, T.; Johnson, J.; Verma, S.K.; Krishnamurthy, P.; Mackie, A.R.; Vaughan, E.; Garikipati, V.N.; Benedict, C.; et al. Embryonic stem cell-derived exosomes promote endogenous repair mechanisms and enhance cardiac function following myocardial infarction. Circ. Res. 2015, 117, 52–64.
- Wang, N.; Chen, C.; Yang, D.; Liao, Q.; Luo, H.; Wang, X.; Zhou, F.; Yang, X.; Yang, J.; Zeng, C.; et al. Mesenchymal stem cells-derived extracellular vesicles, via miR-210, improve infarcted cardiac function by promotion of angiogenesis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2085–2092.
- Zhu, J.; Lu, K.; Zhang, N.; Zhao, Y.; Ma, Q.; Shen, J.; Lin, Y.; Xiang, P.; Tang, Y.; Hu, X.; et al. Myocardial reparative functions of exosomes from mesenchymal stem cells are enhanced by hypoxia treatment of the cells via transferring microRNA-210 in an nSMase2-dependent way. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1659–1670.
- Sun, X.; Shan, A.; Wei, Z.; Xu, B. Intravenous mesenchymal stem cell-derived exosomes ameliorate myocardial inflammation in the dilated cardiomyopathy. Biochem. Biophys. Res. Commun. 2018, 503, 2611–2618.
- Potter, D.R.; Miyazawa, B.Y.; Gibb, S.L.; Deng, X.; Togaratti, P.P.; Croze, R.H.; Srivastava, A.K.; Trivedi, A.; Matthay, M.; Holcomb, J.B.; et al. Mesenchymal stem cell-derived extracellular vesicles attenuate pulmonary vascular permeability and lung injury induced by hemorrhagic shock and trauma. J. Trauma Acute Care Surg. 2018, 84, 245–256.
- Tang, X.D.; Shi, L.; Monsel, A.; Li, X.Y.; Zhu, H.L.; Zhu, Y.G.; Qu, J.M. Mesenchymal Stem Cell Microvesicles Attenuate Acute Lung Injury in Mice Partly Mediated by Ang-1 mRNA. Stem Cells 2017, 35, 1849–1859.
- Ahn, S.Y.; Park, W.S.; Kim, Y.E.; Sung, D.K.; Sung, S.I.; Ahn, J.Y.; Chang, Y.S. Vascular endothelial growth factor mediates the therapeutic efficacy of mesenchymal stem cell-derived extracellular vesicles against neonatal hyperoxic lung injury. Exp. Mol. Med. 2018, 50, 26.
- Khatri, M.; Richardson, L.A.; Meulia, T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell. Res. Ther. 2018, 9, 17.
- Chaubey, S.; Thueson, S.; Ponnalagu, D.; Alam, M.A.; Gheorghe, C.P.; Aghai, Z.; Singh, H.; Bhandari, V. Early gestational mesenchymal stem cell secretome attenuates experimental bronchopulmonary dysplasia in part via exosome-associated factor TSG-6. Stem Cell. Res. Ther. 2018, 9, 173.
- Haga, H.; Yan, I.K.; Borrelli, D.A.; Matsuda, A.; Parasramka, M.; Shukla, N.; Lee, D.D.; Patel, T. Extracellular vesicles from bone marrow-derived mesenchymal stem cells protect against murine hepatic ischemia/reperfusion injury. Liver Transpl. 2017, 23, 791–803.
- Tamura, R.; Uemoto, S.; Tabata, Y. Immunosuppressive effect of mesenchymal stem cell-derived exosomes on a concanavalin A-induced liver injury model. Inflamm. Regen. 2016, 36, 26.
- Rigo, F.; De Stefano, N.; Navarro-Tableros, V.; David, E.; Rizza, G.; Catalano, G.; Gilbo, N.; Maione, F.; Gonella, F.; Roggio, D.; et al. Extracellular Vesicles from Human Liver Stem Cells Reduce Injury in an Ex Vivo Normothermic Hypoxic Rat Liver Perfusion Model. Transplantation 2018, 102, e205–e210.
- Li, T.; Yan, Y.; Wang, B.; Qian, H.; Zhang, X.; Shen, L.; Wang, M.; Zhou, Y.; Zhu, W.; Li, W.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013, 22, 845–854.
- Qu, Y.; Zhang, Q.; Cai, X.; Li, F.; Ma, Z.; Xu, M.; Lu, L. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J. Cell. Mol. Med. 2017, 21, 2491–2502.
- Yan, Y.; Jiang, W.; Tan, Y.; Zou, S.; Zhang, H.; Mao, F.; Gong, A.; Qian, H.; Xu, W. hucMSC Exosome-Derived GPX1 Is Required for the Recovery of Hepatic Oxidant Injury. Mol. Ther. 2017, 25, 465–479.
- Haga, H.; Yan, I.K.; Takahashi, K.; Matsuda, A.; Patel, T. Extracellular Vesicles from Bone Marrow-Derived Mesenchymal Stem Cells Improve Survival from Lethal Hepatic Failure in Mice. Stem Cells Transl. Med. 2017, 6, 1262–1272.
- Lee, M.W.; Ryu, S.; Kim, D.S.; Lee, J.W.; Sung, K.W.; Koo, H.H.; Yoo, K.H. Mesenchymal stem cells in suppression or progression of hematologic malignancy: Current status and challenges. Leukemia 2019, 33, 597–611.
- Christodoulou, I.; Goulielmaki, M.; Devetzi, M.; Panagiotidis, M.; Koliakos, G.; Zoumpourlis, V. Mesenchymal stem cells in preclinical cancer cytotherapy: A systematic review. Stem Cell. Res. Ther. 2018, 9, 336.
- Hendijani, F.; Javanmard, S.H.; Sadeghi-aliabadi, H. Human Wharton’s jelly mesenchymal stem cell secretome display antiproliferative effect on leukemia cell line and produce additive cytotoxic effect in combination with doxorubicin. Tissue Cell 2015, 47, 229–234.
- Ho, J.H.; Chen, Y.F.; Ma, W.H.; Tseng, T.C.; Chen, M.H.; Lee, O.K. Cell contact accelerates replicative senescence of human mesenchymal stem cells independent of telomere shortening and p53 activation: Roles of Ras and oxidative stress. Cell Transplant. 2011, 20, 1209–1220.
- Colter, D.C.; Class, R.; DiGirolamo, C.M.; Prockop, D.J. Rapid expansion of recycling stem cells in cultures of plastic-adherent cells from human bone marrow. Proc. Natl. Acad. Sci. USA 2000, 97, 3213–3218.
- Patel, D.B.; Gray, K.M.; Santharam, Y.; Lamichhane, T.N.; Stroka, K.M.; Jay, S.M. Impact of cell culture parameters on production and vascularization bioactivity of mesenchymal stem cell-derived extracellular vesicles. Bioeng. Transl. Med. 2017, 2, 170–179.
- Kulkarni, R.; Bajaj, M.; Ghode, S.; Jalnapurkar, S.; Limaye, L.; Kale, V.P. Intercellular Transfer of Microvesicles from Young Mesenchymal Stromal Cells Rejuvenates Aged Murine Hematopoietic Stem Cells. Stem Cells 2018, 36, 420–433.
- Wang, X.; Omar, O.; Vazirisani, F.; Thomsen, P.; Ekstrom, K. Mesenchymal stem cell-derived exosomes have altered microRNA profiles and induce osteogenic differentiation depending on the stage of differentiation. PLoS ONE 2018, 13, e0193059.
- Ruan, X.F.; Ju, C.W.; Shen, Y.; Liu, Y.T.; Kim, I.M.; Yu, H.; Weintraub, N.; Wang, X.L.; Tang, Y. Suxiao Jiuxin pill promotes exosome secretion from mouse cardiac mesenchymal stem cells in vitro. Acta Pharmacol. Sin. 2018, 39, 569–578.
- Shi, H.; Xu, X.; Zhang, B.; Xu, J.; Pan, Z.; Gong, A.; Zhang, X.; Li, R.; Sun, Y.; Yan, Y.; et al. 3,3’-Diindolylmethane stimulates exosomal Wnt11 autocrine signaling in human umbilical cord mesenchymal stem cells to enhance wound healing. Theranostics 2017, 7, 1674–1688.
- Harting, M.T.; Srivastava, A.K.; Zhaorigetu, S.; Bair, H.; Prabhakara, K.S.; Toledano Furman, N.E.; Vykoukal, J.V.; Ruppert, K.A.; Cox, C.S., Jr.; Olson, S.D. Inflammation-Stimulated Mesenchymal Stromal Cell-Derived Extracellular Vesicles Attenuate Inflammation. Stem Cells 2018, 36, 79–90.
- Patel, D.B.; Santoro, M.; Born, L.J.; Fisher, J.P.; Jay, S.M. Towards rationally designed biomanufacturing of therapeutic extracellular vesicles: Impact of the bioproduction microenvironment. Biotechnol. Adv. 2018, 36, 2051–2059.
- Watson, D.C.; Bayik, D.; Srivatsan, A.; Bergamaschi, C.; Valentin, A.; Niu, G.; Bear, J.; Monninger, M.; Sun, M.; Morales-Kastresana, A.; et al. Efficient production and enhanced tumor delivery of engineered extracellular vesicles. Biomaterials 2016, 105, 195–205.
- Jarmalaviciute, A.; Tunaitis, V.; Pivoraite, U.; Venalis, A.; Pivoriunas, A. Exosomes from dental pulp stem cells rescue human dopaminergic neurons from 6-hydroxy-dopamine-induced apoptosis. Cytotherapy 2015, 17, 932–939.
- Zhang, Y.; Chopp, M.; Zhang, Z.G.; Katakowski, M.; Xin, H.; Qu, C.; Ali, M.; Mahmood, A.; Xiong, Y. Systemic administration of cell-free exosomes generated by human bone marrow derived mesenchymal stem cells cultured under 2D and 3D conditions improves functional recovery in rats after traumatic brain injury. Neurochem. Int. 2017, 111, 69–81.
More
Information
Subjects:
Cell Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.0K
Revisions:
2 times
(View History)
Update Date:
26 May 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No