 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Miriana d'Alessandro | + 2161 word(s) | 2161 | 2021-04-25 05:20:03 | | | |
| 2 | Lily Guo | Meta information modification | 2161 | 2021-04-27 03:36:11 | | | | |
| 3 | Lily Guo | -1 word(s) | 2160 | 2021-04-27 03:49:56 | | |
Video Upload Options
The house dust mite (HDM) is globally ubiquitous in human habitats. The present entry summarizes information about the role of Der p 1 as key coordinator of the HDM-induced allergic response and reports a series of Italian patients allergic to HDM. Our study offers insights into the role of Der p groups in a population of patients with allergic rhinitis and asthma who were candidate for ITS. Interestingly, Der p1 positivity was associated with bronchial asthma and co-sensitization.
1. Introduction
The house dust mite (HDM) is globally ubiquitous in human habitats. It is considered the main indoor allergen cause of allergic rhinitis and asthma [1]. At total of 600 million people suffer from allergic rhinitis and 200 million cases are concomitant with asthma, as reported by The World Health Organization (WHO) [2][3]. Allergies are common in 50% of all adult asthmatic patients, and up to 85% of them are allergic to HDMs, independent of differences in geography, temperature, and humidity [3].According to the WHO position paper [4][5], the best approach to allergic rhinitis consists in allergen avoidance, pharmacotherapy (including antihistamines, leukotriene receptor antagonists, and inhaled/intranasal corticosteroids), and allergen immunotherapy. Allergen immunotherapy has been used in selected patients for 50 years to achieve clinical tolerance to causative allergens through administration of allergen extracts. It is effective in alleviating the symptoms of allergic rhinitis and/or asthma and in improving quality of life. Allergen immunotherapy has been used in selected patients to achieve clinical tolerance to allergens through exposure of patients to different amounts of allergen extracts in order to modify the immune system’s response [6][7].The main HDM species include Dermatophagoides pteronyssius (Der p), Dermatophagoides farina (Der f), and Blomia tropicalis, which coexist in most geographical regions [8]. Group 1 (Der p 1, Der f1), group 2 (Der p 2, Der f 2), and group 23 (Der p 23) are considered the dominant allergens [9][10][11].Twenty-one group 1 allergens have been found to have protease activity that can destroy epithelial tight junctions. Thirty-two group 2 allergens may mimic the effect of Toll-like receptor 4 coreceptor MD-2, and new ones are still being reported [12][13]. In group 23, a new allergen has been identified as a main allergen, a gut-derived peritrophin in the outer membrane of mite feces. This allergen reacts with IgE antibodies in 74% of patients allergic to Der p [13][14].The first aim of the present study was to summarize the information about the role of Der p1 as a key coordinator of the HDM-induced allergic response. The second aim was to investigate a series of Italian patients allergic to HDM who were screened for anti-HDM treatment in order to establish the role of specific IgE in Der p groups measured by ImmunoCAP 250.In 1980, Chapman and Platts-Mills were the first to isolate the HDM allergen Der p 1 [8]. Der p 1 has been associated with different paradigms over the past 30 years [15][16][17][18][19][20][21].Der p 1 is a papain-like cysteine protease [10][12]. It is considered to be not only the most abundant HDM allergen in house dust and mite cultures but also a powerful allergenic protein [15][22][23]. High levels of IgE specific for this protease have been developed by more than 80% of patients allergic to HDMs [15][24][25][26].In the last 10 years, several authors identified in vitro the protease precursors of the mite cysteine (Der p1) and serine (Der p 3, Der p 6, and Der p 9) protease precursors [27][28][29][30]. The crystal structure of Der p 1 demonstrates that this allergen is a papain-like cysteine protease, while sequence homologies and protease inhibition assays have shown that Der p 3, Der p 6, and Der p 9 belong to the trypsin-like, chymotrypsin-like, and collagenolytic-like serine protease families, respectively [31]. Der p1, 3, 6, and 9 are synthesized as pre-zymogens, consisting of a signal peptide needed for secretion and an N-terminal propeptide, followed by the mature protease domain. The expression of prosequences inhibit the respective proteases to prevent cell toxicity. Literature data reported the role of Der p 1 as the “maestro” in the maturation processes of the different HDM protease allergens [15][16][17]. The behavior of Der p1 appears to be rather uncommon in the protease world and depends on specific sequences at the C-terminus of the different propeptides [32]. Although the biological roles of these proteases in mites have not been completely unraveled, the allergens probably have a digestive function for the mite, as they are detected in the gut as well as in mite feces.The proteolytically active HDM allergens have a critical role in the initiation of an allergic response. The elucidation of the maturation pathways of these allergens is required and could provide insights into their proteolytic specificities. Moreover, the corresponding protein substrates on the innate target and adaptive immune cells will need to be identified. The initial simplistic representation of Der p 1 as an HDM-digestive cysteine protease with IgE-binding properties has been replaced by a much more complex scenario [26][32]. Recent evidence shows a pivotal role of Der p 1 in the maturation of HDM serine protease allergens, suggesting that Der p 1 protein substrates could shape the allergenic potential of HDM proteins. Huge progress has highlighted the key role of environmental and microbial adjuvant factors in activation of innate immune pathways that are essential for the development of allergic responses [33][34][35][36]. These factors combine with the intrinsic biological activity of the allergens, which stimulates innate immune pathways, leading to allergy.Different mechanisms influence the proteolytic activity of HDM allergens in the development of the allergic response, including cleavage of lung epithelium surfactant proteins (SP-A, SP-D) [37], tight junction protein occludins, and immune receptors expressed by dendritic [38], B [39], and T [40] cells; activation of damage-associated molecular patterns (DAMPS) [41] and protease-activated receptors (PARs) expressed on airway epithelial cells [42]; and inactivation of protease inhibitors such as α1-antitrypsin [43]. Altogether, cleavage of these cell receptors and secreted proteins by HDM proteases influences the development of allergic sensitization and may exacerbate allergic inflammation by promoting a pro-Th2 environment and/or by downregulating Th1/Treg differentiation [44].However, the different cell pathways triggered by active Der p 1 are still unclear due to the difficulty of isolating natural Der p 1, free of serine protease contamination, from HDM allergen extracts Accordingly, these limitations stimulated research into the contribution of Der p 1 protease activity in developing a HDM-induced allergic response by means of papain, a structurally similar cysteine protease that is considered to be a surrogate for Der p 1 and HDM allergen extracts, in the presence or absence of cysteine protease inhibitors. Kubo M. reported that proteolytically active papain stimulates an innate cell network (IL-33–ILC-2–IL-13 axis) that can trigger eosinophilia in patients with early airway inflammation in the absence of specific IgE production [45]. It is intriguing to speculate that a similar mechanism could be activated by Der p 1. Cayrol et al. showed than Der p 1 can regulate the cytokine activity of alarmin IL-33 through cleavage of its sensor domain [46].Controversial data exist on the capacity of Der p 1 to activate PAR-2 (not commonly activated by cysteine proteases), leading to proinflammatory cytokine release. The finding may have been due to residual HDM serin protease contamination of Der p 1. Recently, Der p 1 was shown to indirectly stimulate PAR-1/PAR-4 signaling pathways through thrombin, their canonical activator [47].Bioinformatic tools and peptide substrate libraries are useful for predicting potential new targets of Der p 1 in the human cell-surface proteome, targets that could play a role in allergic airway inflammation [17][43]. Besides the capacity of Der p 1 to cleave IL-10 and IL-12 receptors, offering a new pathway for promotion of Th2-polarization, Der p 1 can cleave cell-surface inflammatory receptors (including IL-2 type II, IL-17, IL-18, and IL-23), suggesting that it is involved in a complex balance of pro- and anti-inflammatory effects [48].Many questions still remain about the possibility of Der p 1 being a digestive enzyme and of its role as a potent activator of other mite allergens and innate immune pathways. However, the drastic effect on allergen content and release from mite fecal pellets through the protease activity of Der p1 confirms the role of Der p 1 as a key coordinator of the HDM-induced allergic response.
2. Case Series
Sensitization to mite allergens in the first years of life has a significant impact on lung function in pediatric populations who suffer from wheezing and has been associated with poorer long-term clinical outcomes. This might explain why the approach advocated by current guidelines for allergic rhinitis (AR) (ARIA, Allergic Rhinitis and its Impact on Asthma) [49] and allergic asthma (GINA, Global Initiative for Asthma) [50] classifies disease based on the severity of symptoms, often leaving the underlying allergic cause unaddressed.
2.1. Aim
The present study analyzed the specific IgE profiles in a population of patients with allergic asthma who were undergoing rhinitis anti-HDM treatment, with the aim of obtaining insights into the pathogenic role of Der p 1 (Figure 1).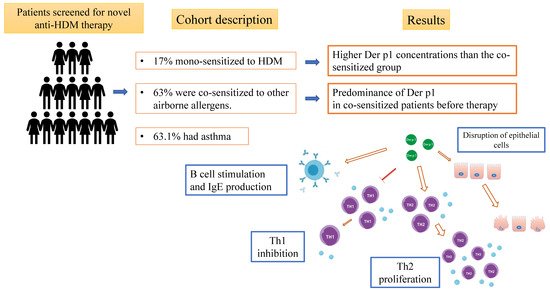 Figure 1. A summary of the aim and main results of the present study, including the biological role of Der p1.All patients gave written informed consent to participate in the study, which was approved by the local ethics committee C.E.A.V.S.E. (code number 180712; Markerlung 17431).
Figure 1. A summary of the aim and main results of the present study, including the biological role of Der p1.All patients gave written informed consent to participate in the study, which was approved by the local ethics committee C.E.A.V.S.E. (code number 180712; Markerlung 17431).
2.2. Methods
A total of 38 patients (M/F: 20/18; median (interquartile range, IQR) age 35 (26–40) years) were screened: eight were mono-sensitized to HDM, 13 were co-sensitized to other airborne allergens, and the others were co-sensitized to other allergens. Fifteen (39.4%) had asthma and the other 23 (60%) had allergic rhinitis. Specific IgE for Der p 1, Der p 2, Der p 10, and Der p 23 were measured by ImmunoCAP 250 (Thermo-Fisher Scientific, Uppsala, Sweden) as previously reported [51]. Levels >0.35 kU/L were considered positive. From August 2019, nineteen patients (M/F: 13/6; median (IQR) age 32 (27–40) years) were enrolled for anti-HDM treatment. The inclusion criteria were persistent allergic rhinitis (moderate-to-severe) to dust mites, despite the use of symptomatic medications, and dust mite allergic asthma not well controlled by inhaled corticosteroids, associated with mild-to-severe dust mite allergic rhinitis. Seven of nineteen patients were mono-sensitized to HDM and 12 were co-sensitized to other airborne allergens. Twelve (63.1%) had asthma. Nasal symptoms (runny nose/nasal drip, nasal congestion/stuffiness, sneezing, and itchy nose) were tallied as the total nasal symptom score (TNSS; max. 12) expressed as median ± standard deviation.
2.3. Results
The prevalence of rhinitis and/or asthma did not show gender or age differences. TNSS was 7.82 ± 0.51 before starting the anti-HDM treatment. Patients co-sensitized to other airborne allergens showed a higher prevalence of asthma (9/12 (75%) vs. 2/7 (29%); p < 0.05) than did HDM mono-sensitized patients. The latter group showed higher Der p1 concentrations than that of the co-sensitized group (p = 0.0360), and a direct correlation between Der p1 and Der p2 (r = 0.93; p = 0.0003) was observed.An AR exacerbation responder was defined as a patient with no AR exacerbations during the 8-week efficacy evaluation period. All of our patients were classified as responders to anti-HDM treatment (based on the above definition of responders) and seven of them (17%) reached a one-year follow-up. The other ones continued to be followed beyond one year.
2.4. Discussion
Our study reports on a cohort of patients screened for anti-HDM treatment. This treatment has been available in Italy since June 2019 and was recently decreed to be free of charge for Tuscan residents. Our data showed a predominance of Der p1 in co-sensitized patients before therapy. Pioneering studies showed that Der p 1 is not only the most abundant allergen from mite fecal pellets but also a major inducer of the IgE response in patients with HDM allergies [17][52]. Although Der p 1 is mainly present in the mite digestive tract, its actual biological function (in relation to the development of asthma and rhinitis) is still unclear. Literature data highlighted the pivotal role of Der p 1 in the maturation of HDM serine protease allergens, suggesting the allergenic potential of HDM proteins could be shaped by the processing of protein substrates [32]. As the first allergen characterized, Der p 1 was naturally chosen to develop a general paradigm for predicting protein allergenicity based on protease activity. Proteolytic activities have subsequently been demonstrated by biochemical and structural characterization of Der p 1 [12][17]. Great progress in immunology concomitantly evidenced the role of environmental and microbial factors in activation of innate immune pathways, which are essential for the development of allergic responses [21][23][46]. Taken together, the Der p 1 paradigm inevitably shifted to a more broadly applicable model of protein allergenicity: the allergy is caused by the association of intrinsic biological activity of an allergen along with its ability to stimulate innate immune pathways. The cysteine protease activity of Der p1 promotes a Th2-allergic response, even if the elucidation of different cell pathways triggered by active Der p 1 is still challenging. Future studies will involve solving the technical problem of isolating natural Der p 1 from HDM allergen extracts (isolation from serine protease contamination or preparation in reproducible batches).Chevigné A et al. suggested potential new targets for Der p 1 in the human cell-surface proteome. These could play a role in allergic airway inflammation [17]. The increased concentrations of Der p1 and the high prevalence of asthma in our co-sensitized HDM patients before treatment with anti-HDM treatment suggest that underlying severe inflammation supports allergic airway diseases.In conclusion, our study offers insights into the role of Der p groups in a population of patients with allergic rhinitis and asthma who were candidate for specific immunotherapy. Interestingly, Der p1 positivity was associated with bronchial asthma and co-sensitization.
References
- Frankland, A.W. Asthma and mites. Postgrad Med. J. 1971, 47, 178–180.
- Mims, J.W. Epidemiology of allergic rhinitis. Int. Forum Allergy Rhinol. 2014, 4 (Suppl S2), S18–S20.
- Öçal, R.; Bayar Muluk, N.; Mullol, J. Epidemiology of Allergic Rhinitis. In All Around the Nose: Basic Science, Diseases and Surgical Management; Cingi, C., Bayar Muluk, N., Eds.; Springer International Publishing: Cham, Switzerland, 2020.
- Koyanagi, S.; Murakami, T.; Maeda, T.; Kawatsu, K.; Okamura, H.; Oda, Y.; Miyatsu, Y.; Sugawara, K.; Mizokami, H. Production-scale purification of the recombinant major house dust mite allergen Der f 2 mutant C8/119S. J. Biosci. Bioeng. 2010, 110, 597–601.
- Bousquet, J.; Lockey, R.; Malling, H.J. Allergen immunotherapy: Therapeutic vaccines for allergic diseases. A WHO position paper. J. Allergy Clin. Immunol. 1998, 102, 558–562.
- Inal, A.; Altintas, D.U.; Yilmaz, M.; Karakoc, G.B.; Kendirli, S.G.; Sertdemir, Y. Prevention of new sensitizations by specific immunotherapy in children with rhinitis and/or asthma monosensitized to house dust mite. J. Investig. Allergol. Clin. Immunol. 2007, 17, 85–91.
- Jacobsen, L.; Niggemann, B.; Dreborg, S.; Ferdousi, H.A.; Halken, S.; Høst, A.; Koivikko, A.; Norberg, L.A.; Valovirta, E.; Wahn, U.; et al. Specific immunotherapy has long-term preventive effect of seasonal and perennial asthma: 10-year follow-up on the PAT study. Allergy 2007, 62, 943–948.
- Arlian, L.G.; Morgan, M.S.; Neal, J.S. Dust mite allergens: Ecology and distribution. Curr. Allergy Asthma Rep. 2002, 2, 401–411.
- Aggarwal, P.; Senthilkumaran, S. Dust Mite Allergy. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021.
- Thomas, W.R. Hierarchy and molecular properties of house dust mite allergens. Allergol. Int. 2015, 64, 304–311.
- Bessot, J.-C.; Pauli, G. House dust mites allergens. Rev. Mal. Respir. 2011, 28, 475–495.
- Cui, Y. Structural biology of mite allergens. Mol. Biol. Rep. 2013, 40, 681–686.
- Celi, G.; Brusca, I.; Scala, E.; Villalta, D.; Pastorello, E.; Farioli, L.; Cortellini, G.; Deleonardi, G.; Galati, P.; Losappio, L.; et al. House dust mite allergy in Italy-Diagnostic and clinical relevance of Der p 23 (and of minor allergens): A real-life, multicenter study. Allergy 2019, 74, 1787–1789.
- Weghofer, M.; Grote, M.; Resch, Y.; Casset, A.; Kneidinger, M.; Kopec, J.; Thomas, W.R.; Fernández-Caldas, E.; Kabesch, M.; Ferrara, R.; et al. Identification of Der p 23, a peritrophin-like protein, as a new major Dermatophagoides pteronyssinus allergen associated with the peritrophic matrix of mite fecal pellets. J. Immunol. 2013, 190, 3059–3067.
- Yang, X.; Fan, G.; Li, J. Diagnostic value of Der p 1 and Der p 2 specific IgE in Dermatophagoides pteronyssinus IgE sensitization. Ann. Allergy. Asthma Immunol. 2016, 116, 295–301.
- Tian, M.; Zhou, Y.; Zhang, W.; Cui, Y. Der p 1 and Der p 2 specific immunoglobulin E measurement for diagnosis of Dermatophagoides pteronyssinus allergy: A systematic review and meta-analysis. Allergy Asthma Proc. 2017, 38, 333–342.
- Chevigné, A.; Jacquet, A. Emerging roles of the protease allergen Der p 1 in house dust mite-induced airway inflammation. J. Allergy Clin. Immunol. 2018, 142, 398–400.
- Bessot, J.C.; Pauli, G. Mite allergens: An overview. Eur. Ann. Allergy Clin. Immunol. 2011, 43, 141–156.
- Mattoli, S. Allergen-induced generation of mediators in the mucosa. Environ. Health Perspect. 2001, 109 (Suppl. S4), 553–557.
- Yao, Y.; Wang, Z.-C.; Yu, D.; Liu, Z. Role of allergen-specific T-follicular helper cells in immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2018, 18, 495–501.
- Bush, R.K. Indoor allergens, environmental avoidance, and allergic respiratory disease. Allergy Asthma Proc. 2008, 29, 575–579.
- Bronnert, M.; Mancini, J.; Birnbaum, J.; Agabriel, C.; Liabeuf, V.; Porri, F.; Cleach, I.; Fabre, A.; Deneux, I.; Grandne, V.; et al. Component-resolved diagnosis with commercially available D. pteronyssinus Der p 1, Der p 2 and Der p 10: Relevant markers for house dust mite allergy. Clin. Exp. Allergy 2012, 42, 1406–1415.
- Wilson, J.M.; Platts-Mills, T.A.E. Home Environmental Interventions for House Dust Mite. J. Allergy Clin. Immunol. Pract. 2018, 6, 1–7.
- Chapman, M.D.; Rowntree, S.; Mitchell, E.B.; Di Prisco de Fuenmajor, M.C.; Platts-Mills, T.A. Quantitative assessments of IgG and IgE antibodies to inhalant allergens in patients with atopic dermatitis. J. Allergy Clin. Immunol. 1983, 72, 27–33.
- Thomas, W.R. IgE and T-cell responses to house dust mite allergen components. Mol. Immunol. 2018, 100, 120–125.
- Chapman, M.D.; Wünschmann, S.; Pomés, A. Proteases as Th2 adjuvants. Curr. Allergy Asthma Rep. 2007, 7, 363–367.
- Reddy, V.B.; Lerner, E.A. Activation of mas-related G-protein-coupled receptors by the house dust mite cysteine protease Der p1 provides a new mechanism linking allergy and inflammation. J. Biol. Chem. 2017, 292, 17399–17406.
- Dumez, M.-E.; Teller, N.; Mercier, F.; Tanaka, T.; Vandenberghe, I.; Vandenbranden, M.; Devreese, B.; Luxen, A.; Frère, J.M.; Matagne, A.; et al. Activation mechanism of recombinant Der p 3 allergen zymogen: Contribution of cysteine protease Der p 1 and effect of propeptide glycosylation. J. Biol. Chem. 2008, 283, 30606–30617.
- López-Rodríguez, J.C.; Manosalva, J.; Cabrera-García, J.D.; Escribese, M.M.; Villalba, M.; Barber, D.; Martínez-Ruiz, A.; Batanero, E. Human glutathione-S-transferase pi potentiates the cysteine-protease activity of the Der p 1 allergen from house dust mite through a cysteine redox mechanism. Redox Biol. 2019, 26, 101256.
- Miranda, D.O.; Silva, D.A.O.; Fernandes, J.F.C.; Queirós, M.G.J.; Chiba, H.F.; Ynoue, L.H.; Resende, R.O.; Pena, J.; Sung, S.S.J.; Segundo, G.R.; et al. Serum and salivary IgE, IgA, and IgG4 antibodies to Dermatophagoides pteronyssinus and its major allergens, Der p1 and Der p2, in allergic and nonallergic children. Clin. Dev. Immunol. 2011, 302739.
- Thomas, W.R.; Smith, W.-A.; Hales, B.J.; Mills, K.L.; O’Brien, R.M. Characterization and immunobiology of house dust mite allergens. Int. Arch. Allergy Immunol. 2002, 129, 1–18.
- Dumez, M.-E.; Herman, J.; Campizi, V.; Galleni, M.; Jacquet, A.; Chevigné, A. Orchestration of an Uncommon Maturation Cascade of the House Dust Mite Protease Allergen Quartet. Front. Immunol. 2014, 5, 138.
- Bao, Z.-J.; Fan, Y.-M.; Cui, Y.-F.; Sheng, Y.-F.; Zhu, M. Effect of PM2.5 mediated oxidative stress on the innate immune cellular response of Der p1 treated human bronchial epithelial cells. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2907–2912.
- Thomas, R.G.; Rivera Reyes, B.M.; Gaston, B.M.; Rivera Acosta, N.B.; Bederman, I.R.; Smith, L.A.; Sutton, M.T.; Wang, B.; Hunt, J.F.; Bonfield, T.L. Conjugation of nitrated acetaminophen to Der p1 amplifies peripheral blood monocyte response to Der p1. PLoS ONE 2017, 12, e0188614.
- Genç, D.; Zibandeh, N.; Nain, E.; Arığ, Ü.; Göker, K.; Aydıner, E.K.; Akkoç, T.U.N.Ç. IFN-γ stimulation of dental follicle mesenchymal stem cells modulates immune response of CD4+ T lymphocytes in Der p1+ asthmatic patients in vitro. Allergol. Immunopathol. (Madr.) 2019, 47, 467–476.
- Yu, S.; Jin, L.; Che, N.; Zhang, R.; Xu, F.; Han, B. Dendritic cells modified with Der p1 antigen as a therapeutic potential for allergic rhinitis in a murine model via regulatory effects on IL-4, IL-10 and IL-13. Int. Immunopharmacol. 2019, 70, 216–224.
- Deb, R.; Shakib, F.; Reid, K.; Clark, H. Major house dust mite allergens Dermatophagoides pteronyssinus 1 and Dermatophagoides farinae 1 degrade and inactivate lung surfactant proteins A and D. J. Biol. Chem. 2007, 282, 36808–36819.
- Furmonaviciene, R.; Ghaemmaghami, A.M.; Boyd, S.E.; Jones, N.S.; Bailey, K.; Willis, A.C.; Sewell, H.F.; Mitchell, D.A.; Shakib, F. The protease allergen Der p 1 cleaves cell surface DC-SIGN and DC-SIGNR: Experimental analysis of in silico substrate identification and implications in allergic responses. Clin. Exp. Allergy 2007, 37, 231–242.
- Schulz, O.; Sutton, B.J.; Beavil, R.L.; Shi, J.; Sewell, H.F.; Gould, H.J.; Laing, P.; Shakib, F. Cleavage of the low-affinity receptor for human IgE (CD23) by a mite cysteine protease: Nature of the cleaved fragment in relation to the structure and function of CD23. Eur. J. Immunol. 1997, 27, 584–588.
- Schulz, O.; Sewell, H.F.; Shakib, F. Proteolytic cleavage of CD25, the alpha subunit of the human T cell interleukin 2 receptor, by Der p 1, a major mite allergen with cysteine protease activity. J. Exp. Med. 1998, 187, 271–275.
- Hara, K.; Iijima, K.; Elias, M.K.; Seno, S.; Tojima, I.; Kobayashi, T.; Kita, H. Airway uric acid is a sensor of inhaled protease allergens and initiates type 2 immune responses in respiratory mucosa. J. Immunol. 2014, 192, 4032–4042.
- Kato, T.; Takai, T.; Fujimura, T.; Matsuoka, H.; Ogawa, T.; Murayama, K.; Ishii, A.; Ikeda, S.; Okumura, K.; Ogawa, H. Mite serine protease activates protease-activated receptor-2 and induces cytokine release in human keratinocytes. Allergy 2009, 64, 1366–1374.
- Kalsheker, N.A.; Deam, S.; Chambers, L.; Sreedharan, S.; Brocklehurst, K.; Lomas, D.A. The house dust mite allergen Der p1 catalytically inactivates alpha 1-antitrypsin by specific reactive centre loop cleavage: A mechanism that promotes airway inflammation and asthma. Biochem. Biophys. Res. Commun. 1996, 221, 59–61.
- Cunningham, P.T.; Elliot, C.E.; Lenzo, J.C.; Jarnicki, A.G.; Larcombe, A.N.; Zosky, G.R.; Holt, P.G.; Thomas, W.R. Sensitizing and Th2 adjuvant activity of cysteine protease allergens. Int. Arch. Allergy Immunol. 2012, 158, 347–358.
- Kubo, M. Innate and adaptive type 2 immunity in lung allergic inflammation. Immunol. Rev. 2017, 278, 162–172.
- Cayrol, C.; Duval, A.; Schmitt, P.; Roga, S.; Camus, M.; Stella, A.; Burlet-Schiltz, O.; Gonzalez-de-Peredo, A.; Girard, J.P. Environmental allergens induce allergic inflammation through proteolytic maturation of IL-33. Nat. Immunol. 2018, 19, 375–385.
- Zhang, J.; Chen, J.; Allen-Philbey, K.; Perera Baruhupolage, C.; Tachie-Menson, T.; Mangat, S.C.; Garrod, D.R.; Robinson, C. Innate generation of thrombin and intracellular oxidants in airway epithelium by allergen Der p 1. J. Allergy Clin. Immunol. 2016, 138, 1224–1227.
- Jacquet, A.; Campisi, V.; Szpakowska, M.; Dumez, M.-E.; Galleni, M.; Chevigné, A. Profiling the Extended Cleavage Specificity of the House Dust Mite Protease Allergens Der p 1, Der p 3 and Der p 6 for the Prediction of New Cell Surface Protein Substrates. Int. J. Mol. Sci. 2017, 18, 1373.
- Klimek, L.; Bachert, C.; Pfaar, O.; Becker, S.; Bieber, T.; Brehler, R.; Buhl, R.; Casper, I.; Chaker, A.; Czech, W.; et al. ARIA guideline 2019: Treatment of allergic rhinitis in the German health system. Allergol. Select 2019, 3, 22–50.
- Global Initiative for Asthma—GINA. Available online: (accessed on 25 December 2020).
- D’alessandro, M.; Bergantini, L.; Perrone, A.; Beltrami, V.; Cameli, P.; Flori, L.; Saletti, M.; Vietri, L.; Sestini, P.; Bargagli, E. A real-life experience with ImmunoCAP ISAC: The advantages of a new diagnostic method. Minerva Med. 2020.
- Chapman, M.D.; Platts-Mills, T.A. Purification and characterization of the major allergen from Dermatophagoides pteronyssinus-antigen P1. J. Immunol. 1980, 125, 587–592.


