1000/1000
Hot
Most Recent
 +1 point
+1 point
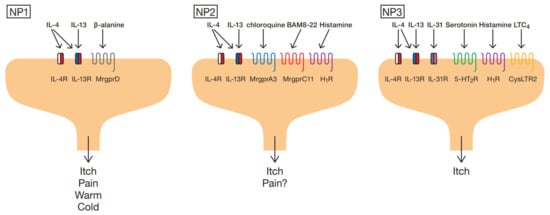
Although histamine is a well-known itch mediator, histamine H1-receptor blockers often lack efficacy in chronic itch. Recent molecular and cellular based studies have shown that non-histaminergic mediators, such as proteases, neuropeptides and cytokines, along with their cognate receptors, are involved in evocation and modulation of itch sensation. Many of these molecules are produced and secreted by immune cells, which act on sensory nerve fibers distributed in the skin to cause itching and sensitization. This understanding of the connections between immune cell-derived mediators and sensory nerve fibers has led to the development of new treatments for itch.
| Category | Pruritogens | Receptors | Therapeutic Methods | Reference |
|---|---|---|---|---|
| Amines | Histamine | H1R/H4R | Anti-histamine/Anti-inflammatory, immuno-modulatory topical and systemic therapy (Cyclosporine A, Pimecrolimus, Tacrolimus and Corticosteroids) | [6][14] |
| Serotonin | 5-HT2 receptor | Sertraline | [15] | |
| Proteases | Tryptase | PAR-2 | Anti-histamine/Cyclosporine A/Pimecrolimus/Tacrolimus/Corticosteroids | [6] |
| Chymase | PAR-2 | ONO-WH-236/Anti-histamine/Cyclosporine A/Pimecrolimus/Tacrolimus/Corticosteroids | [6][16] | |
| Cathepsin S | PAR-2/PAR-4 | LHVS/Anti-histamine/Cyclosporine A/Pimecrolimus/Tacrolimus/Corticosteroids | [6][17] | |
| Peptides | Substance P | NK-1R | Serlopitant/Gabapentin/Pregabalin/Capsaicin | [6][18] |
| Endothelin-1 | ETA | Bosentan | [19] | |
| cytokines | IL-2 | IL-2R | Cyclosporine A/Delgocitinib/Baricitinib/Abrocitinib | [14][20][21][22][23] |
| IL-4 | IL-4Rα/γC | Dupilumab/Delgocitinib/Baricitinib/Abrocitinib | [14][24][25][26][23] | |
| IL-4Rα/IL-13Rα1 | ||||
| IL-13 | IL-4Rα/IL-13Rα1 | Dupilumab/Tralokinumab/Lebrikizumab | [14][24][25] | |
| IL-17 | IL-17RA/IL-17RC | Brodalumab | [27] | |
| IL-23 | IL-12Rβ1/IL-23R | Delgocitinib/Baricitinib | [14][28][29][23] | |
| IL-31 | IL-31RA/OSMR | Nemolizumab/Delgocitinib/Baricitinib/Abrocitinib | [14][30][31][32][33][23][34] | |
| IL-33 | ST2/IL-1RAcP | Etokimab/Delgocitinib/Baricitinib | [14][35][23] | |
| TSLP | TSLPR | Tezepelumab/Delgocitinib/Baricitinib/Abrocitinib | [14][36][37][23][38] | |
| Lipid mediators | PAF | PAFR | PAF antagonist | [39][40] |
| LTB4 | BLT1/BLT2 | CMHVA | [41][42] | |
| LTC4 | CysLTR1/CysLTR2 | CysLTR2 antagonist | [43] |
| Ligands | Receptors | Source | Modulation |
|---|---|---|---|
| SLIGRL-NH2 | PAR-2 | mast cells, basophils | Enhances CQ and BAM8-22 induced itch |
| IL-4 | IL-4Rα/γC IL-4Rα/IL-13Rα1 |
Th2, Tfh, ILC2, mast cells, basophils, eosinophils | Enhanced neuronal responsiveness to histamine, CQ, TSLP and IL-31 |
| IL-13 | IL-13Rα1/IL-13Rα2 | Th2, ILC2, mast cells, basophils, eosinophils | May enhance neuronal responsiveness to histamine, CQ, TSLP and IL-31, as well as IL-4 |
| IL-23 | IL-12Rβ1/IL-23R | DCs, macrophages | Reduced histamine-induced itch |
| IL-33 | ST2/IL-1RAcP | DCs, macrophages, mast cells | Enhanced CQ evoked calcium responses |





