 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hong Cao | + 2601 word(s) | 2601 | 2021-10-14 05:51:24 | | | |
| 2 | Peter Tang | Meta information modification | 2601 | 2021-10-22 08:25:06 | | |
Video Upload Options
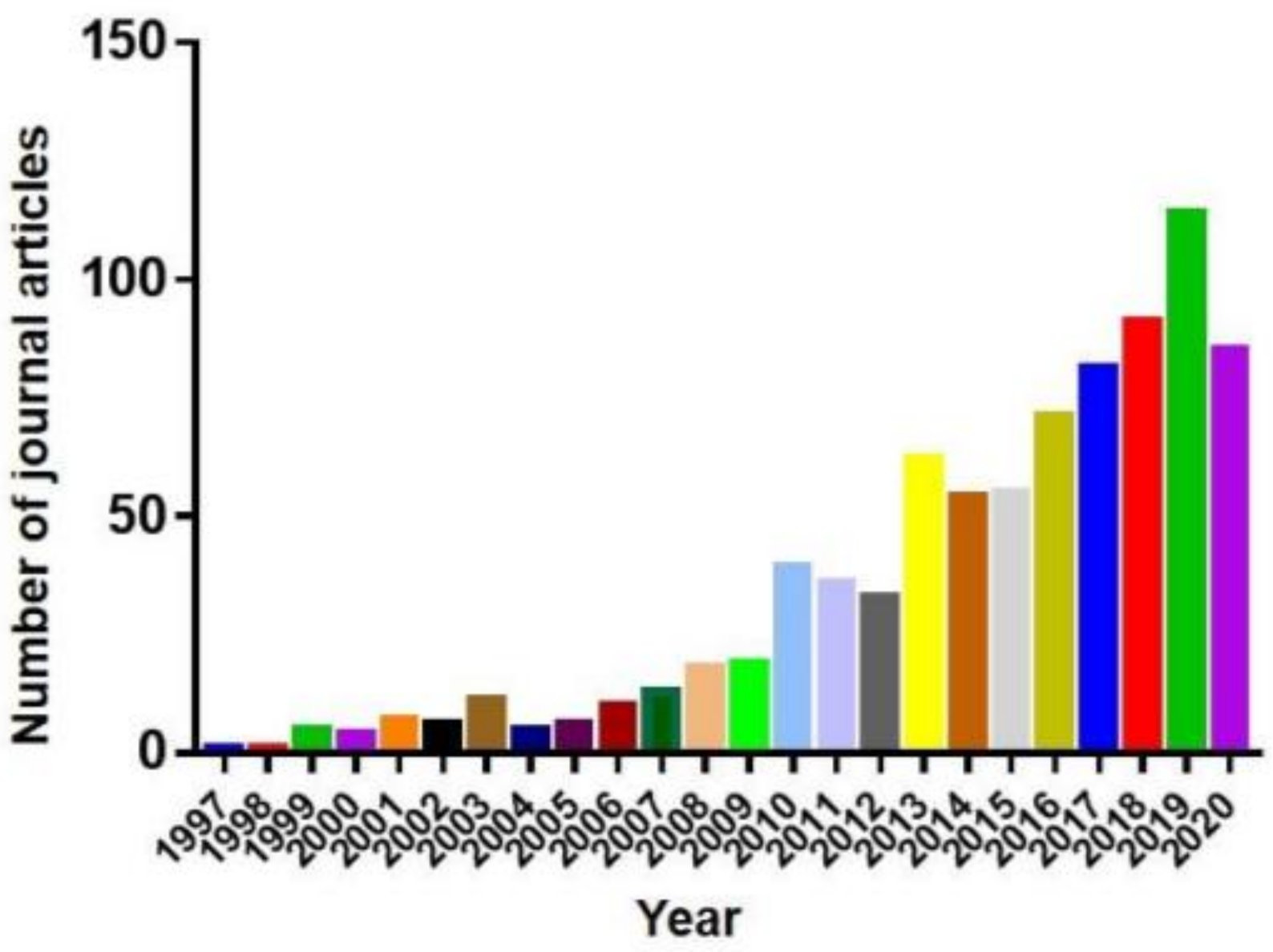
Animal models of human neurodegenerative disease have been investigated for several decades. In recent years, zebrafish (Danio rerio) and medaka (Oryzias latipes) have become popular in pathogenic and therapeutic studies about human neurodegenerative diseases due to their small size, the optical clarity of embryos, their fast development, and their suitability to large-scale therapeutic screening.
1. Introduction
2. Parkinson’s Disease Models
|
Method |
Phenotype |
Results |
Reference |
|---|---|---|---|
|
MPTP induced |
Motor impairments and weakened touch sensory |
Reduction of locomotor activity and dopaminergic neuron, over-expression of synuclein in the optic tectum |
|
|
6-OHDA induced |
Motor impairments and anxiety |
Reduction of dopaminergic neurons and morphological alternations |
|
|
Paraquat induced |
Motor impairments, various developmental anomalies |
The paraquat-treated zebrafish did not recapitulate PD pathology |
|
|
Rotenone induced |
Motor impairments, anxiety, and olfactory dysfunction |
In addition to motor impairments, they also show Olfactory dysfunction, which is a typical non-motor symptom of PD |
|
|
PARK2 Morpholino |
No abnormalities in swimming behavior |
Loss of the DA neuron numbers in the diencephalon, whereas no abnormalities in swimming behavior |
|
|
PINK1 Morpholino; Transgenes |
Motor impairment and oxidative stress |
Reduction of dopaminergic neurons, dis-organized diencephalic dopaminergic neurons, and the pink1 gene are sensitive markers of oxidative stress in zebrafish |
|
|
LRRK2 Morpholino |
Motor impairment |
Loss of neuronal cells and synuclein aggregation, similar to the phenotype of PD in humans |
|
|
PARK7 Morpholino; CRISPR/Cas9 |
Motor impairment |
With aging, exhibit lower TH levels, respiratory failure in skeletal muscle, and lower body mass, particularly in the male fish |
|
|
Synuclein Transgenes |
Motor impairment |
Led to cell death in larval zebrafish sensory neurons |
[61] |
|
GBA TALEN |
Motor impairment |
Reduction of the GBA protein, dopaminergic, and noradrenergic neurons |
|
|
PARL Morpholino; CRISPR/Cas9 |
Motor impairment and olfactory dysfunction |
Reduced DA neuronal population and dysregulation of the PINK1/Parkin mitophagy pathway |
3. Alzheimer’s Disease Models
|
Method |
Phenotype |
Results |
Reference |
|---|---|---|---|
|
Amyloid-β42 induced |
Intracellular depositions |
Link between aging, neurogenesis, regenerative, neuroinflammation, and neural stem cell plasticity |
|
|
Okadaic acid induced |
Cognitive and memory impairments, neuroinflammation cholinergic dysfunction, glutamate excitotoxicity, and mitochondrial dysfunction |
Provide all the major molecular hallmarks of AD |
|
|
Cigarette smoke extract induced |
Neurocognitive dysfunction |
Enhancement of the acetylcholinesterase activity |
|
|
Aluminum chloride induced |
Neurocognitive dysfunction, memory impairment |
Impaired locomotor activity, learning, and memory abilities |
[80] |
|
Copper induced |
Memory impairment |
Reduction of the glutathione S-transferase (GST) activity in the gill |
[81] |
|
MnCl2 induced |
Cognition and exploratory behavior |
Impairment of aversive long-term memory and distance traveled movement time |
[82] |
|
MAPT Transgenes |
Motor impairment |
The phenotypic abnormalities at larval stages make it suitable for high-throughput screening |
|
|
PSEN1 ENU-mutagenized |
Motor impairment |
Regulation of histaminergic neuron development |
[85] |
|
BACE1/2 zinc finger nuclease; ENU-mutagenized |
Hypomyelination, supernumery neuromasts, and abnormal pigmentation |
Bace1 and Bace2 are proteases with different physiological functions |
[86] |
4. Huntington’s Disease Models
5. Other Neurodegenerative Disease Models
References
- Heemels, M.T. Neurodegenerative diseases. Nature 2016, 539, 179.
- Gitler, A.D.; Dhillon, P.; Shorter, J. Neurodegenerative disease: Models, mechanisms, and a new hope. Dis. Models Mech. 2017, 10, 499–502.
- Fernaández-Trapero, M.; Espejo-Porras, F.; Rodrıguez-Cueto, C.; Coates, J.R.; Peérez-Díaz, C.; De Lago, E.; Fernaández-Ruiz, J. Upregulation of CB2 receptors in reactive astrocytes in canine degenerative myelopathy, a disease model of amyotrophic lateral sclerosis. Dis. Model Mech. 2017, 10, 551–558.
- Snyder, B.R.; Chan, A.W.S. Progress in Developing Transgenic Monkey Model for Huntington’s Disease. J. Neural Transm. 2018, 125, 401–417.
- Story, B.D.; Miller, M.E.; Bradbury, A.M.; Million, E.D.; Duan, D.; Taghian, T.; Faissler, D.; Fernau, D.; Beecy, S.J.; Gray-Edwards, H.L. Canine Models of Inherited Musculoskeletal and Neurodegenerative Diseases. Front. Vet. Sci. 2020, 7, 80.
- Bolus, H.; Crocker, K.; Boekhoff-Falk, G.; Chtarbanova, S. Modeling Neurodegenerative Disorders in Drosophila melanogaster. Int. J. Mol. Sci. 2020, 21, 3055.
- Horzmann, K.A.; Freeman, J.L. Making waves: New developments in toxicology with the zebrafish. Toxicol. Sci. 2018, 163, 5–12.
- Paone, C.; Diofano, F.; Park, D.D.; Rottbauer, W.; Just, S. Genetics of cardiovascular disease: Fishing for causality. Front. Cardiovasc. Med. 2018, 5, 60.
- Vaz, R.L.; Outeiro, T.F.; Ferreira, J.J. Zebrafish as an animal model for drug discovery in parkinson’s disease and other movement disorders: A systematic review. Front. Neurol. 2018, 9, 347.
- Saleem, S.; Kannan, R.R. Zebrafish: An emerging real-time model system to study Alzheimer’s disease and neurospecific drug discovery. Cell Death Discov. 2018, 4, 45.
- Furutani-Seiki, M.; Wittbrodt, J. Medaka and zebrafish, an evolutionary twin study. Mech. Dev. 2004, 121, 629–637.
- Dang, M.; Henderson, R.E.; Garraway, L.A.; Zon, L.I. Long-term drug administration in the adult zebrafish using oral gavage for cancer preclinical studies. Dis. Model Mech. 2016, 9, 811–820.
- Matsuzaki, Y.; Hosokai, H.; Mizuguchi, Y.; Fukamachi, S.; Shimizu, A.; Saya, H. Establishment of HRAS(G12V) transgenic medaka as a stable tumor model for in vivo screening of anticancer drugs. PLoS ONE 2013, 8, e54424.
- Fior, R.; Póvoa, V.; Mendes, R.V.; Carvalho, T.; Gomes, A.; Figueiredo, N.; Ferreira, M.G. Single-cell functional and chemosensitive profiling of combinatorial colorectal therapy in zebrafish xenografts. Proc. Natl. Acad. Sci. USA 2017, 114, E8234–E8243.
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503.
- Apfeld, J.; Alper, S. What Can We Learn About Human Disease from the Nematode C. elegans? Methods Mol. Biol. 2018, 1706, 53–75.
- Yamaguchi, M.; Yoshida, H. Drosophila as a Model Organism. Adv. Exp. Med. Biol. 2018, 1076, 1–10.
- Kim, S.S.; Im, S.H.; Yang, J.Y.; Lee, Y.R.; Kim, G.R.; Chae, J.S.; Shin, D.S.; Song, J.S.; Ahn, S.; Lee, B.H.; et al. Zebrafish as a Screening Model for Testing the Permeability of Blood-Brain Barrier to Small Molecules. Zebrafish 2017, 14, 322–330.
- Panula, P.; Chen, Y.C.; Priyadarshini, M.; Kudo, H.; Semenova, S.; Sundvik, M.; Sallinen, V. The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases. Neurobiol. Dis. 2010, 40, 46–57.
- Matsui, H.; Uemura, N.; Yamakado, H.; Takeda, S.; Takahashi, R. Exploring the pathogenetic mechanisms underlying Parkinson’s disease in medaka fish. J. Parkinsons Dis. 2014, 4, 301–310.
- Wittbrodt, J.; Shima, A.; Schartl, M. Medaka—a model organism from the Far East. Nat. Rev. Genet. 2002, 3, 53–64.
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueff, A.V. Zebrafish models for translational neuroscience research: From tank to bedside. Trends Neurosci. 2014, 37, 264–278.
- GBD 2016 Parkinson’s Disease Collaborators. Global, regional, and national burden of Parkinson’s disease, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953.
- Savica, R.; Grossardt, B.R.; Bower, J.H.; Ahlskog, J.E.; Boeve, B.F.; Graff-Radford, J.; Rocca, W.A.; Mielke, M.M. Survival and Causes of Death among People with Clinically Diagnosed Synucleinopathies with Parkinsonism: A Population-Based Study. JAMA Neurol. 2017, 74, 839–846.
- Darweesh, S.K.L.; Raphael, K.G.; Brundin, P.; Matthews, H.; Wyse, R.K.; Chen, H.; Bloem, B.R. Parkinson Matters. J. Parkinsons Dis. 2018, 8, 495–498.
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858.
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, Regional, and National Age-Sex-Specific Mortality for 282 Causes of Death in 195 Countries and Territories, 1980–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788.
- Rocca, W.A. The burden of Parkinson’s disease: A worldwide perspective. Lancet Neurol. 2018, 17, 928–929.
- Pirtošek, Z.; Bajenaru, O.; Kovács, N.; Milanov, I.; Relja, M.; Skorvanek, M. Update on the Management of Parkinson’s Disease for General Neurologists. Parkinson’s Dis. 2020, 2020, 9131474.
- Hoo, J.Y.; Kumari, Y.; Shaikh, M.F.; Hue, S.M.; Goh, B.H. Zebrafish: A Versatile Animal Model for Fertility Research. BioMed Res. Int. 2016, 2016, 9732780.
- Rahman Khan, F.; Sulaiman Alhewairini, S. Zebrafish (Danio rerio) as a Model Organism. Curr. Trends Cancer Manag. 2019, 2019, 81517.
- Matsui, H.; Takahashi, R. Parkinson’s disease pathogenesis from the viewpoint of small fish models. J. Neural Transm. 2018, 125, 25–33.
- Barnhill, L.M.; Murata, H.; Bronstein, J.M. Studying the pathophysiology of Parkinson’s disease using zebrafish. Biomedicines 2020, 8, 8070197.
- Lam, C.S.; Korzh, V.; Strahle, U. Zebrafish embryos are susceptible to the dopaminergic neurotoxin MPTP. Eur. J. Neurosci. 2005, 21, 1758–1762.
- Wasel, O.; Freeman, J.L. Chemical and genetic zebrafish models to define mechanisms of and treatments for dopaminergic neurodegeneration. Int. J. Mol. Sci. 2020, 21, 5981.
- Sarath, B.N.; Murthy, C.H.L.; Kakara, S.; Sharma, R.; Brahmendra, S.C.V.; Idris, M.M. 1-Methyl-4-phenyl-1,2,3,6-tetrahydro-pyridine induced Parkinson’s disease in zebrafish. Proteomics 2016, 16, 1407–1420.
- Vijayanathan, Y.; Lim, F.T.; Lim, S.M.; Long, C.M.; Tan, M.P.; Majeed, A.B.A.; Ramasamy, K. 6-OHDA-Lesioned Adult Zebrafish as a Useful Parkinson’s Disease Model for Dopaminergic Neuroregeneration. Neurotox. Res. 2017, 32, 496–508.
- Vaz, R.L.; Sousa, S.; Chapela, D.; van der Linde, H.C.; Willemsen, R.; Correia, A.D.; Outeiro, T.F.; Afonso, N.D. Identification of antiparkinsonian drugs in the 6-hydroxydopamine zebrafish model. Pharmacol. Biochem. Behav. 2020, 189, 172828.
- Li, M.; Zhou, F.; Xu, T.; Song, H.; Lu, B. Acteoside protects against 6-OHDA-induced dopaminergic neuron damage via Nrf2-ARE signaling pathway. Food Chem. Toxicol. 2018, 119, 6–13.
- Cronin, A.; Grealy, M. Neuroprotective and neuro-restorative effects of minocycline and rasagiline in a zebrafish 6-hydroxydopamine model of Parkinson’s disease. Neuroscience 2017, 367, 34–46.
- Wang, X.H.; Souders, C.L.; Zhao, Y.H.; Martyniuk, C.J. Paraquat affects mitochondrial bioenergetics, dopamine system expression, and locomotor activity in zebrafish (Danio rerio). Chemosphere 2018, 191, 106–117.
- Nellore, J.P.N. Paraquat exposure induces behavioral deficits in larval zebrafish during the window of dopamine neurogenesis. Toxicol. Rep. 2015, 2, 950–956.
- Bortolotto, J.W.; Cognato, G.P.; Christoff, R.R.; Roesler, L.N.; Leite, C.E.; Kist, L.W.; Bogo, M.R.; Vianna, M.R.; Bonan, C.D. Long-term exposure to paraquat alters behavioral parameters and dopamine levels in adult zebrafish (Danio rerio). Zebrafish 2014, 11, 142–153.
- Müller, T.E.; Nunes, M.E.; Menezes, C.C.; Marins, A.T.; Leitemperger, J.; Gressler, A.C.L.; Carvalho, F.B.; de Freitas, C.M.; Quadros, V.A.; Fachinetto, R.; et al. Sodium Selenite Prevents Paraquat-Induced Neurotoxicity in Zebrafish. Mol. Neurobiol. 2018, 55, 1928–1941.
- Lv, D.J.; Li, L.X.; Chen, J.; Wei, S.Z.; Wang, F.; Hu, H.; Xie, A.M.; Liu, C.F. Sleep deprivation caused a memory defects and emotional changes in a rotenone-based zebrafish model of Parkinson’s disease. Behav. Brain Res. 2019, 372, 112031.
- Ramli, M.D.B.C.; Hashim, N.H.B.; Uzid, M.B.M.; Weinheimeri, A.Z. Zebrafish parkinson’s model: The effects of tocotrienol rich fraction towards rotenone induced zebrafish. Int. J. Med. Toxicol. Legal Med. 2020, 23, 1–7.
- Ünal, İ.; Üstündağ, Ü.V.; Ateş, P.S.; Eğilmezer, G.; Alturfan, A.A.; Yiğitbaşı, T.; Emekli-Alturfan, E. Rotenone impairs oxidant/antioxidant balance both in brain and intestines in zebrafish. Int. J. Neurosci. 2019, 129, 363–368.
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s disease and its potential as therapeutic target. Trans. Neurodegener. 2015, 4, 40–42.
- Ge, P.; Dawson, V.L.; Dawson, T.M. PINK1 and Parkin mitochondrial quality control: A source of regional vulnerability in Parkinson’s disease. Mole Neurodegener. 2020, 15, 7.
- Flinn, L.; Mortiboys, H.; Volkmann, K.; Kster, R.W.; Ingham, P.W.; Bandmann, O. Complex I deficiency and dopaminergic neuronal cell loss in parkin-deficient zebrafish (Danio rerio). Brain 2009, 132, 1613–1623.
- Sallinen, V.; Kolehmainen, J.; Priyadarshini, M.; Toleikyte, G.; Chen, Y.C.; Panula, P. Dopaminergic cell damage and vulnerability to MPTP in PINK1 knockdown zebrafish. Neurobiol. Dis. 2010, 40, 93–101.
- Xi, Y.; Ryan, J.; Noble, S.; Yu, M.; Yilbas, A.E.; Ekker, M. Impaired dopaminergic neuron development and locomotor function in zebrafish with loss of pink1 function. Eur. J. Neurosci. 2010, 31, 623–633.
- Sloan, M.; Alegre-Abarrategui, J.; Wade-Martins, R. Insights into LRRK2 function and dysfunction from transgenic and knockout rodent models. Biochem. Soc. Trans. 2012, 40, 1080–1085.
- Prabhudesai, S.; Bensabeur, F.Z.; Abdullah, R.; Basak, I.; Baez, S.; Alves, G.; Holtzman, N.G.; Larsen, J.P.; Møller, S.G. LRRK2 knockdown in zebrafish causes developmental defects, neuronal loss, and synuclein aggregation. J. Neurosci. Res. 2016, 94, 17–35.
- Sheng, D.; Qu, D.; Kwok, K.H.; Ng, S.S.; Lim, A.Y.; Aw, S.S.; Lee, C.W.; Sung, W.K.; Tan, E.K.; Lufkin, T.; et al. Deletion of the WD40 domain of LRRK2 in Zebrafish causes Parkinsonism-like loss of neurons and locomotive defect. PLoS Genet. 2010, 6, e1000914.
- Seegobin, S.P.; Heaton, G.R.; Liang, D.; Choi, I.; Blanca Ramirez, M.; Tang, B.; Yue, Z. Progress in LRRK2-Associated Parkinson’s Disease Animal Models. Front. Neurosci. 2020, 14, 674.
- Hughes, G.L.; Lones, M.A.; Bedder, M.; Currie, P.D.; Smith, S.L.; Pownall, M.E. Machine learning discriminates a movement disorder in a zebrafish model of Parkinson’s disease. Dis. Model Mech. 2020, 13, dmm045815.
- Bai, Q.; Mullett, S.J.; Garver, J.A.; Hinkle, D.A.; Burton, E.A. Zebrafish DJ-1 is evolutionarily conserved and expressed in dopaminergic neurons. Brain Res. 2006, 1113, 33–44.
- Bretaud, S.; Allen, C.; Ingham, P.W.; Bandmann, O. p53-dependent neuronal cell death in a DJ-1-deficient zebrafish model of Parkinson’s disease. J. Neurochem. 2007, 100, 1626–1635.
- Edson, A.J.; Hushagen, H.A.; Frøyset, A.K.; Elda, I.; Khan, E.A.; Di Stefano, A.; Fladmark, K.E. Dysregulation in the Brain Protein Profile of Zebrafish Lacking the Parkinson’s Disease-Related Protein DJ-1. Mol. Neurobiol. 2019, 56, 8306–8322.
- O’Donnell, K.C.; Lulla, A.; Stahl, M.C.; Wheat, N.D.; Bronstein, J.M.; Sagasti, A. Axon degeneration and PGC-1a-mediated protection in a zebrafish model of a-synuclein toxicity. Dis. Models Mech. 2014, 7, 571–582.
- Keatinge, M.; Bui, H.; Menke, A.; Chen, Y.C.; Sokol, A.M.; Bai, Q.; Ellett, F.; Da Costa, M.; Burke, D.; Gegg, M.; et al. Glucocerebrosidase 1 deficient Danio rerio mirror key pathological aspects of human Gaucher disease and provide evidence of early microglial activation preceding alpha-synuclein-independent neuronal cell death. Hum. Mol. Genet. 2015, 24, 6640–6652.
- Matsui, H.; Ito, J.; Matsui, N.; Uechi, T.; Onodera, O.; Kakita, A. Cytosolic dsDNA of mitochondrial origin induces cytotoxicity and neurodegeneration in cellular and zebrafish models of Parkinson’s disease. Nat. Commun. 2021, 12, 3101.
- Noble, S.; Ismail, A.; Godoy, R.; Xi, Y.; Ekker, M. Zebrafish Parla- and Parlb-deficiency affects dopaminergic neuron patterning and embryonic survival. J. Neurochem. 2012, 122, 196–207.
- Merhi, R.; Kalyn, M.; Zhu-Pawlowsky, A.; Ekker, M. Loss of parla Function Results in Inactivity, Olfactory Impairment, and Dopamine Neuron Loss in Zebrafish. Biomedicines 2021, 9, 205.
- Alzheimer’s Association. 2020 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2020, 16, 391–460.
- Alzheimer’s Disease International. World Alzheimer Report 2019: Attitudes to Dementia; Alzheimer’s Disease International (ADI): London, UK, 2019.
- Vaz, M.; Silvestre, S. Alzheimer’s disease: Recent treatment strategies. Eur. J. Pharmacol. 2020, 887, 173554.
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789.
- Cummings, J.L.; Cohen, S.; van Dyck, C.H.; Brody, M.; Curtis, C.; Cho, W.; Ward, M.; Friesenhahn, M.; Rabe, C.; Brunstein, F.; et al. ABBY: A phase 2 randomized trial of crenezumab in mild to moderate Alzheimer disease. Neurology 2018, 90, e1889–e1897.
- Alzheimer’s Association. 2018 Alzheimer’s disease facts and figures. Alzheimers Dement. 2018, 14, 367–429.
- Ruhl, T.; Jonas, A.; Seidel, N.I.; Prinz, N.; Albayram, O.; Bilkei-Gorzo, A.; von der Emde, G. Oxidation and Cognitive Impairment in the Aging Zebrafish. Gerontology 2015, 62, 47–57.
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Zhang, Y.; Kizil, C. Modeling Amyloid-β42 Toxicity and Neurodegeneration in Adult Zebrafish Brain. J. Vis. Exp. 2017, 128, 56014.
- Bhattarai, P.; Thomas, A.K.; Zhang, Y.; Kizil, C. The effects of aging on Amyloid-β42-induced neurodegeneration and regeneration in adult zebrafish brain. Neurogenesis 2017, 4, e1322666.
- Rudrabhatla, P.; Pant, H.C. Role of protein phosphatase 2A in Alzheimer’s disease. Curr. Alzheimer Res. 2011, 8, 623–632.
- Nada, S.E.; Williams, F.E.; Shah, Z.A. Development of a Novel and Robust Pharmacological Model of Okadaic Acid-induced Alzheimer’s Disease in Zebrafish. CNS Neurol. Disord. Drug Targets 2016, 15, 86–94.
- Koehler, D.; Williams, F.E. Utilizing zebrafish and okadaic acid to study Alzheimer’s disease. Neural Regen. Res. 2018, 13, 1538–1541.
- Muthuraman, A.; Thilagavathi, L.; Jabeen, S.; Ravishankar, S.B.; Ahmed, S.S.; George, T.; Rishitha, N.; Paramakrishnan, N. Curcumin prevents cigarette smoke extract induced cognitive impairment. Front. Biosci. 2019, 11, 109–120.
- Muthuraman, A.; Nafisa, K.; Sowmya, M.S.; Arpitha, B.M.; Choedon, N.; Sandy, C.D.; Rishitha, N.; Johurul, I. Role of ambrisentan (selective endothelin-A receptor antagonist) on cigarette smoke exposure induced cognitive impairment in Danio rerio. Life Sci. 2019, 222, 133–139.
- He, X.; Zhong, Z.M.; Che, Y. Locomotor activity and learning and memory abilities in Alzheimer’s disease induced by aluminum in an acid environment in zebrafish. Dongwuxue Yanjiu 2012, 33, 231–236.
- Acosta, D.D.S.; Danielle, N.M.; Altenhofen, S.; Luzardo, M.D.; Costa, P.G.; Bianchini, A.; Bonan, C.D.; da Silva, R.S.; Dafre, A.L. Copper at low levels impairs memory of adult zebrafish (Danio rerio) and affects swimming performance of larvae. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 185–186, 122–130.
- Altenhofen, S.; Wiprich, M.T.; Nery, L.R.; Leite, C.E.; Vianna, M.R.M.R.; Bonan, C.D. Manganese(II) chloride alters behavioral and neurochemical parameters in larvae and adult zebrafish. Aquat. Toxicol. 2017, 182, 172–183.
- Bai, Q.; Garver, J.A.; Hukriede, N.A.; Burton, E.A. Generation of a transgenic zebrafish model of Tauopathy using a novel promoter element derived from the zebrafish eno2 gene. Nucleic Acids Res. 2007, 35, 6501–6516.
- Paquet, D.; Bhat, R.; Sydow, A.; Mandelkow, E.M.; Berg, S.; Hellberg, S.; Fälting, J.; Distel, M.; Köster, R.W.; Schmid, B.; et al. A zebrafish model of tauopathy allows in vivo imaging of neuronal cell death and drug evaluation. J. Clin. Investig. 2009, 119, 1382–1395.
- Sundvik, M.; Chen, Y.C.; Panula, P. Presenilin1 regulates histamine neuron development and behavior in zebrafish, danio rerio. J. Neurosci. 2013, 33, 1589–1597.
- van Bebber, F.; Hruscha, A.; Willem, M.; Schmid, B.; Haass, C. Loss of Bace2 in zebrafish affects melanocyte migration and is distinct from Bace1 knock out phenotypes. J. Neurochem. 2013, 127, 471–481.
- Shannon, K.M. Huntington’s disease—Clinical signs, symptoms, presymptomatic diagnosis, and diagnosis. Handb. Clin. Neurol. 2011, 100, 3–13.
- Ross, C.A.; Aylward, E.H.; Wild, E.J.; Langbehn, D.R.; Long, J.D.; Warner, J.H.; Scahill, R.I.; Leavitt, B.R.; Stout, J.C.; Paulsen, J.S.; et al. Huntington disease: Natural history, biomarkers and prospects for therapeutics. Nat. Rev. Neurol. 2014, 10, 204–216.
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228.
- The Huntington’s Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983.
- Duyao, M.P.; Auerbach, A.B.; Ryan, A.; Persichetti, F.; Barnes, G.T.; McNeil, S.M.; Ge, P.; Vonsattel, J.P.; Gusella, J.F.; Joyner, A.L.; et al. Inactivation of the mouse Huntington’s disease gene homolog Hdh. Science 1995, 269, 407–410.
- Zeitlin, S.; Liu, J.P.; Chapman, D.L.; Papaioannou, V.E.; Efstratiadis, A. Increased apoptosis and early embryonic lethality in mice nullizygous for the Huntington’s disease gene homologue. Nat. Genet. 1995, 11, 155–163.
- Nasir, J.; Floresco, S.B.; O’Kusky, J.R.; Diewert, V.M.; Richman, J.M.; Zeisler, J.; Borowski, A.; Marth, J.D.; Phillips, A.G.; Hayden, M.R. Targeted disruption of the Huntington’s disease gene results in embryonic lethality and behavioral and morphological changes in heterozygotes. Cell 1995, 81, 811–823.
- Karlovich, C.A.; John, R.M.; Ramirez, L.; Stainier, D.Y.; Myers, R.M. Characterization of the Huntington’s disease (HD) gene homologue in the zebrafish Danio rerio. Gene 1998, 217, 117–125.
- Lumsden, A.L.; Henshall, T.L.; Dayan, S.; Lardelli, M.T.; Richards, R.I. Huntingtin-deficient zebrafish exhibit defects in iron utilization and development. Hum. Mol. Genet. 2007, 16, 1905–1920.
- Henshall, T.L.; Tucker, B.; Lumsden, A.L.; Nornes, S.; Lardelli, M.T.; Richards, R.I. Selective neuronal requirement for huntingtin in the developing zebrafish. Hum. Mol. Genet. 2009, 18, 4830–4842.
- Diekmann, H.; Anichtchik, O.; Fleming, A.; Futter, M.; Goldsmith, P.; Roach, A.; Rubinsztein, D.C. Decreased BDNF levels are a major contributor to the embryonic phenotype of huntingtin knockdown zebrafish. J. Neurosci. 2009, 29, 1343–1349.
- Schiffer, N.W.; Broadley, S.A.; Hirschberger, T.; Tavan, P.; Kretzschmar, H.A.; Giese, A.; Haass, C.; Hartl, F.U.; Schmid, B. Identification of anti-prion compounds as efficient inhibitors of polyglutamine protein aggregation in a zebrafish model. J. Biol. Chem. 2007, 282, 9195–9203.
- Williams, A.; Sarkar, S.; Cuddon, P.; Ttofi, E.K.; Saiki, S.; Siddiqi, F.H.; Jahreiss, L.; Fleming, A.; Pask, D.; Goldsmith, P.; et al. Novel targets for Huntington’s disease in an mTOR-independent autophagy pathway. Nat. Chem. Biol. 2008, 4, 295–305.
- Veldman, M.B.; Rios-Galdamez, Y.; Lu, X.H.; Gu, X.; Qin, W.; Li, S.; Yang, X.W.; Lin, S. The N17 domain mitigates nuclear toxicity in a novel zebrafish Huntington’s disease model. Mol. Neurodegener. 2015, 10, 67.
- VerPlank, J.J.S.; Tyrkalska, S.D.; Fleming, A.; Rubinsztein, D.C.; Goldberg, A.L. cGMP via PKG activates 26S proteasomes and enhances degradation of proteins, including ones that cause neurodegenerative diseases. Proc. Natl. Acad. Sci. USA 2020, 117, 14220–14230.
- Renton, A.E.; Majounie, E.; Waite, A.; Simon-Sanchez, J.; Rollinson, S.; Gibbs, J.R.; Schymick, J.C.; Laaksovirta, H.; van Swieten, J.C.; Myllykangas, L.; et al. A Hexanucleotide Repeat Expansion in C9ORF72 Is the Cause of Chromosome 9p21-Linked ALS-FTD. Neuron 2011, 72, 257–268.
- Sullivan, P.M.; Zhou, X.; Robins, A.M.; Paushter, D.H.; Kim, D.; Smolka, M.B.; Hu, F. The ALS/FTLD associated protein C9orf72 associates with SMCR8 and WDR41 to regulate the autophagy-lysosome pathway. Acta Neuropathol. Commun. 2016, 4, 51.
- Shaw, M.P.; Higginbottom, A.; McGown, A.; Castelli, L.M.; James, E.; Hautbergue, G.M.; Shaw, P.J.; Ramesh, T.M. Stable transgenic C9orf72 zebrafish model key aspects of the ALS/FTD phenotype and reveal novel pathological features. Acta Neuropathol. Commun. 2018, 6, 125.
- Ohki, Y.; Wenninger-Weinzierl, A.; Hruscha, A.; Asakawa, K.; Kawakami, K.; Haass, C.; Edbauer, D.; Schmid, B. Glycine-alanine dipeptide repeat protein contributes to toxicity in a zebrafish model of C9orf72 associated neurodegeneration. Mol. Neurodegener. 2017, 12, 6.
- Yeh, T.H.; Liu, H.F.; Li, Y.W.; Lu, C.S.; Shih, H.Y.; Chiu, C.C.; Lin, S.J.; Huang, Y.C.; Cheng, Y.C. C9orf72 is essential for neurodevelopment and motility mediated by Cyclin G1. Exp. Neurol. 2018, 304, 114–124.
- McGown, A.; Shaw, D.P.J.; Ramesh, T. ZNStress: A high-throughput drug screening protocol for identification of compounds modulating neuronal stress in the transgenic mutant sod1G93R zebrafish model of amyotrophic lateral sclerosis. Mol. Neurodegener. 2016, 11, 56.
- Namikawa, K.; Dorigo, A.; Zagrebelsky, M.; Russo, G.; Kirmann, T.; Fahr, W.; Dübel, S.; Korte, M.; Köster, R.W. Modeling Neurodegenerative Spinocerebellar Ataxia Type 13 in Zebrafish Using a Purkinje Neuron Specific Tunable Coexpression System. J. Neurosci. 2019, 39, 3948–3969.
- Namikawa, K.; Dorigo, A.; Köster, R.W. Neurological Disease Modelling for Spinocerebellar Ataxia Using Zebrafish. J. Exp. Neurosci. 2019, 13, 1179069519880515.
- Lin, Y.; Cai, X.; Wang, G.; Ouyang, G.; Cao, H. Model construction of Niemann-Pick type C disease in zebrafish. Biol. Chem. 2018, 399, 903–910.
- Uemura, N.; Koike, M.; Ansai, S.; Kinoshita, M.; Ishikawa-Fujiwara, T.; Matsui, H.; Naruse, K.; Sakamoto, N.; Uchiyama, Y.; Todo, T.; et al. Viable neuronopathic Gaucher disease model in Medaka (Oryzias latipes) displays axonal accumulation of alpha-synuclein. PLoS Genet. 2015, 11, e1005065.


