Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Evangelos Zoidis | + 7972 word(s) | 7972 | 2021-07-29 10:35:12 | | | |
| 2 | Vivi Li | Meta information modification | 7972 | 2021-09-15 04:32:42 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Zoidis, E. Selenium/Selenoproteins in Male Reproductive Function. Encyclopedia. Available online: https://encyclopedia.pub/entry/14175 (accessed on 07 February 2026).
Zoidis E. Selenium/Selenoproteins in Male Reproductive Function. Encyclopedia. Available at: https://encyclopedia.pub/entry/14175. Accessed February 07, 2026.
Zoidis, Evangelos. "Selenium/Selenoproteins in Male Reproductive Function" Encyclopedia, https://encyclopedia.pub/entry/14175 (accessed February 07, 2026).
Zoidis, E. (2021, September 14). Selenium/Selenoproteins in Male Reproductive Function. In Encyclopedia. https://encyclopedia.pub/entry/14175
Zoidis, Evangelos. "Selenium/Selenoproteins in Male Reproductive Function." Encyclopedia. Web. 14 September, 2021.
Copy Citation
Selenium (Se) is an important trace mineral having many essential roles at the cellular and organismal levels in animal and human health. The biological effects of Se are mainly carried out by selenoproteins (encoded by 25 genes in humans and 24 in mice). As an essential component of selenoproteins, Se performs structural and enzymic roles; in the latter context it is well known for its catalytic and antioxidative functions. Studies involving different animal models have added great value to our understanding regarding the potential implications of Se and selenoproteins in mammalian fertility and reproduction.
male fertility
mammalian reproduction
selenium
selenoproteins
spermatogenesis
1. Introduction
1.1. Background
In 1817, Se was first identified by Jöns Jacob Berzelius, who was conducting an investigation on chemicals responsible for the outbreaks of ill health amongst the workers in a sulfuric acid plant in Sweden [1]. Its basic importance in both normal growth and reproduction in animals was not discovered until the 1950s [2]. However, the definitive manifestation for the importance of Se in humans came through the findings of large-scale trials in China that demonstrated the ameliorative effects of Se supplementation on children and young adults suffering from the Keshan’s disease (characterized by cardiomyopathy); once endemic in areas with low soil Se levels [3][4].
1.2. Implication of Se in Mammalian Reproduction: An Overview
The optimal reproductive efficiency of mammals depends on many factors, such as genetics, nutrition, management, and environmental determinants [5]. Among these, trace mineral nutrition is vital for various biological functions such as, normal growth, development, and reproduction [6]. Furthermore, even the narrow variations in the levels of micronutrients (such as trace minerals) may have considerable bearing on vital biological processes including reproductive health and performance [5][6]. Similarly, as for the Se, there is a constringe window between inadequacy and nimiety and its essential and toxic levels are clearly defined [7][8]. In order to sustain the optimum Se concentrations within cells, enormously controlled mechanisms should be in place [8]. Ideally, for investigating whether or not the Se supplementation could ameliorate the fertility, the reference range for Se adequacy/inadequacy should be pre-defined [7]. The Se concentration, for the most part, depends on the tissue being analyzed [7]. As a matter of fact, it is also unclear which biological fluid viz. blood, serum, seminal plasma, sperm cells, follicular fluid provides the most precise picture of Se concentration with regards to its role in reproduction [7]. Apparently, there is a dearth of scientific information elucidating the relation between serum Se and the levels in the reproductive tissues [7]. Nevertheless, few past reports on mammalian models have demonstrated the relationship between Se status and reproductive performance in both males [5][9] and females [5][10]. The adequate traces of Se in the male reproductive organs are essentially required for normal spermatogenesis, sperm maturation, and sperm motility and overall function [11][12][13][14]. Increased dietary Se intake has also been implicated in enhancing the antioxidant glutathione peroxidase (GPX) activity, thereby improving fertility in male [15].
2. Important Selenoproteins Relevant to Male Reproduction
Eight different enzymatic isoforms of glutathione peroxidase (GPX) family (GPX 1–8) play a vital role in several redox responses. Of note, GPX1 to 4 and GPX6 are selenoproteins, while the other isoforms are Cys-containing analogs instead. Moreover, GPX6 is the lone exception and a selenoprotein in man [16]. The principal role of these enzymatic isoforms is to safeguard and protect the cells from oxidative stress by catalyzing the reduction (via glutathione) of hydrogen peroxide (H2O2), organic hydroperoxides and lipid peroxides. All these are tissue specific and apparently have a sexually dimorphic expression [17].
Glutathione peroxidase 4, also known as PHGPx (phospholipid hydroperoxide GSH Px), is a classical case. It is distinctly expressed in testes and has both an antioxidant as well as a structural role; the latter context is evident from a fact that it constitutes over 50% of mitochondrial capsule (as an oxidatively inactivated protein) in midpiece of mature sperm [9]. In an early stage of spermatogenesis, GPX4 is believed to protect the developing sperm from oxidative stress-induced DNA damage, however, in the later phase, through cross linkage with proteins in midpiece region, it provides the integrity to the sperm midpiece by becoming a structural component of mitochondrial sheath circumventing the flagellum, which is an essential component for sperm stability and motility [18][19][20][21]. Three isoforms with different N-terminal amino acid sequences viz. mitochondrial (mGPX4) and cytosolic (cGPX4) and nuclear variant (sperm nucleus glutathione peroxidase 4, snGPX4) are encoded by a single GPX4 gene. These three isoforms are specifically localized in the mitochondria, cytosol, and nucleus [22]. Following complete targeted disruption of Gpx4, early embryonic lethality in homozygous Gpx4 knockout (KO) mice has been witnessed; however, on the other hand, in spite of their decreased Gpx4 mRNA and protein levels, the heterozygote mice were viable, fertile and appeared normal [22]. Conrad et al. [23] have showed that mice with a directed deletion of snGpx4 gene were not only viable, but also fully fertile, in contrast to the full KO mice. The deletion of cGpx4 variant results in the early developmental defects (reviewed in [24]). On the other hand, Schneider et al. [25] have demonstrated that normal embryonic and postnatal development could be achieved even after targeted deletion of mGpx4 in mouse model; however, this situation leads to infertility in males. Interestingly, resulting infertility was bypassed following intra-cytoplasmic sperm injection of mGpx4−/− sperm and viable offspring were produced [25]. Imai and co-researchers [26] demonstrated the importance of GPX4 for male fertility in humans. They found that GPX4 was abundantly distributed in late spermatocytes and spermatids and localized in the sperm midpiece, particularly in the mitochondria. These authors also report that, among examined subjects, all but 10% of the infertile men demonstrated a striking decline in GPX4 levels in sperm. However, no abnormal expression of GPX4 was observed in sperm from fertile males [26]. These results were, however, confirmed by Foresta and colleagues [9] who reported that both sperm structural integrity and motility were directly related with the concentration of GPX4. With the same token, residual GPX4 activity was significantly reduced in infertile men compared to the controls, and particularly reduced in oligoasthenozoospermic men. This activity showed a direct correlation, most importantly, with the forward motility, but also with the viability and structural integrity [9]. In another study on murine model in 2009, it was further demonstrated that reduced Gpx4 levels lead towards infertility in males [27]. This was further supported by the evidence that mGpx4 KO epididymal sperm were unable to fertilize the oocytes in vitro [27]. Therefore, regardless of cause of reduced GPX4, it might be regarded as the prognostic tool for determining the fertility [9]. In vitro fertilization (IVF) studies have also been extended to assess the general impact of sperm GPX expression on fertilization ability, embryo quality and over all reproductive outcomes. More asymmetric embryos (at day 3) were obtained from sperm samples with a lower GPX4 mRNA expression. However, GPX4 levels showed no effect on the later phase of in vitro development (at days 5 and 6) or on pregnancy rates [28].
In addition to these evidences, recently Parillo and colleagues [29] reported that strong expression signals of GPX4 protein were observed in seminiferous tubules (apical region) of healthy Chianina bulls, where it was localized in the cytoplasmic region of maturing sperm i.e., spermatogonia, round spermatids, and elongated spermatids. Similarly, immuno-signals were also observed in the epididymal and ejaculated sperm particularly in acrosome region. These dynamic strong signals at different stages of sperm maturation conform to the earlier biochemical observations indicating that GPX4 is essential for the optimal development and function of bovine sperm similar to other mammalian species [29].
Like GPX4, SELENOP is also believed to play an essential role in male reproductive functions. It serves as a transport protein for Se and is also expressed in vesicle like structures in the basal region of the Sertoli cells [30]. Besides, Selenop mRNA was also expressed in Leydig cells of rats (discussed later in 3.1). Noticeable reduction in fertility, reduced levels of Se and a lower Gpx activity have been reported in male Selenop KO mice [31]. Meanwhile, a diet containing high Se content could not restitute the testis Se levels or normal sperm phenotype in these mice [30]. X-ray fluorescence microscopic observations in mGpx4 KO mice models have revealed a 60% decline in Se levels, and a resultant greatly impaired spermatogenesis. However, the largest loss of Se was manifested in Selenop KO models; in which a 77% decrease of Se was observed [32]. In addition to these two peculiar and canonical selenoproteins (GPX4, SELENOP), other selenoproteins such as GPX1 and GPX3 were also expressed in the male reproductive tissues and secretions, and have been implicated in male fertility. GPX1 and GPX3 are well represented and characterized and are located in the epididymal epithelia and sperm. These selenoproteins are reported to protect the epididymal parenchyma and maturing sperm from oxidative stress (reviewed in ref. [33]).
The foregoing evidences highlight that Se performs significant functions in the male reproductive system which are regulated by selenoproteins, especially GPX4 and SELENOP. Therefore, it is advisable to perform more studies focusing on the elucidation of additional roles played by these (GPX4 and SELENOP) selenoproteins in male reproductive functions. Nevertheless, search for the elucidation of additional putative mechanisms potentially modulated by other biologically relevant selenoproteins should also be included in the scope of future studies. In any case, selected mammalian selenoproteins (expressed in mouse model) having potential implications in male reproductive function are summarized in Table 1.
Table 1. Brief description of known mammalian selenoproteins relevant to male reproduction.
| Selenoprotein Gene * | Symbol [12] | General Description/Function [8][34][35][36][37][38] |
mRNA * | Protein * | Relevance to Male Reproductive Function |
|---|---|---|---|---|---|
| Glutathione peroxidase 4 | Gpx4 | Detoxification of lipid hydroperoxides, Antioxidant in membranes, functions as structural protein in sperm, also implicated in apoptosis | ++++ | ++++ | Structural protein of sperm midpiece mitochondrial sheath and involved in sperm chromatin condensation [39]. Implication in male fertility [27]. |
| Thioredoxin-glutathione reductase | Txnrd3 (TGR) | Part of the thioredoxin system, Antioxidant function, redox regulation, cell signaling | + | + | Implicated in formation of disulfide bond and sperm maturation process [40]. Expressed in post-pubertal testis, particularly abundant in elongated spermatids at the site of mitochondrial sheath formation [39]. |
| Selenoprotein P | Selenop | Primarily responsible for Se transport and also performs antioxidative role. Considered as a major contributor to plasma Se and a reliable biomarker of Se status. Its deficiency causes infertility characterized by abnormal sperm in mice |
+ | + | Implicated in male fertility [41][42]. Implicated in transport of Se to spermatogenic cells [30]. Essential for sperm development in mice [43]. |
| Selenoprotein V | Selenov | Largely unidentified, potential role in redox regulation | + | n.d. | Specifically expressed in rodent testes [44]. In situ hybridization trials have demonstrated the expression of Selenov mRNA in seminiferous tubules in mouse, however, its precise function in spermatogenesis is largely unexplored [45][44]. |
| Selenoprotein W | Selenow | Antioxidant protection | + | + | n.d. * |
| Selenoprotein K | Selenok | Possible antioxidant protection in cardiomyocytes, Endoplasmic reticulum transmembrane protein | ++ | n.d. | n.d. * |
| Selenoprotein F | Selenof | Role in cell apoptosis and mediation of chemo-preventive effects of Se | + | n.d. | n.d. * |
| Selenoprotein S | Selenos | Cellular redox balance, Possible influence in inflammatory response |
+ | n.d. | n.d. * |
| Selenophosphate synthetase 2 | Sephs2 | Required for biosynthesis of selenophosphate, a precursor of selenocysteine, and thus for selenoprotein synthesis | + | n.d. | n.d. |
n.d.: not defined. Relative expression levels in mouse testis: ++++, very high; ++, modest; +, low. Note: The contents tagged with “*” are adapted by permission from “Springer Nature Customer Service Centre GmbH: Springer Nature, New York, USA” by [39].
3. Role of Selenium in Male Reproduction
In past, Behne and colleagues conducted a set of studies to observe Se regulation and its importance in male rats; these authors reported that, in conditions of deficient Se intake its supply to testes was prioritized over other tissues and organs. Subsequently, results of such studies allowed to assume that Se is potentially involved in testosterone biosynthesis [11][46].
3.1. Role of Selenium in Steroidogenesis and Spermatogenesis
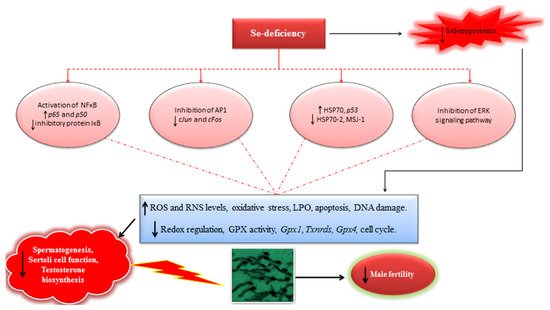
The optimal concentrations of primary sex hormone (testosterone) are essential for normal development of sperm cells. During the process of testosterone biosynthesis, ROS are generated and their excessive production contributes to male infertility [47]. Interestingly, almost two decades ago, Selenop mRNA was selectively identified in Leydig cells of rats [47][48]. It was suggested that, in addition to its role as a plasma selenoprotein transporting Se, Selenop may also play a role as an intracellular antioxidant in Leydig cells [48]. Besides, cytosolic Gpx was also implicated in counteracting the H2O2 generated as a result of testosterone biosynthesis, but its expression in testis was relatively lower [47]. However, as for the effects of Selenop on steroid biosynthesis in Leydig cells, it was proposed that, in addition to its intracellular antioxidant role, Selenop might also act as an extracellular antioxidant protecting Leydig cells from oxidative damage. Subsequently, in 2001, using the mouse and rat models, a physiological function of Selenop in testosterone production was reported in Leydig cells [47]. It was suggested that Selenop plays an important antioxidant role in protecting Leydig cells from the oxidative damage resulting from testosterone biosynthesis pathway [47][48].
In any case, the growing body of literature also suggests that Se is also important for the biosynthesis of testosterone. It has been suggested that blood testosterone concentrations have positive correlation with concentrations of Se [49][50]. Similarly, dietary Se supplementation has been demonstrated to ameliorate the testosterone level and quality of semen in different animal species [12][51][52][53]. However, the exact underlying mechanisms regulating the testosterone production are yet unclear and require further understanding. Since testosterone is produced by Leydig cells, they could serve as a potential target model to further investigate such mechanisms of Se-related modulation of testosterone synthesis [54]. It has been demonstrated that Se has a potential regulatory role in some essential cellular functions. This role is reportedly modulated by activating the extracellular-signal-regulated kinase (ERK) signaling pathways [55]. However, the exact role of Se in biosynthesis of testosterone via modulation of ERK signaling pathway and the fate of Leydig cells throughout the process of sperm development is incompletely understood [54]. In this connection, in an in vitro study, Shi et al. [54] revealed that Se could modulate the proliferative and apoptosis-related cellular events in ovine Leydig cells. These effects were principally achieved via regulation of oxidative stress, cell cycle, and apoptosis-associated biomarkers. The lowest ROS content and the highest GPX activity were found in the 2.0 μmol/L group compared to the control (without Se) and high Se-treated (4.0 and 8.0 μmol/L) groups. They further reported that Se could also increase testosterone biosynthesis in Leydig cells via activation of ERK signaling pathway and expression of its downstream genes i.e., steroidogenic acute regulatory (StAR) and 3 beta-hydroxysteroid dehydrogenase (3β-HSD). These biomarkers are thought to have a close relation with the regulatory implications of Se in fertility and gametogenesis in males. Optimal levels of Se (2.0 μmol/L) triggered a significant up-regulation in the expression of these genes and improved testosterone biosynthesis. However, when the concentration of Se was increased (8.0 μmol/L) in culture medium, the proliferative capacity of Leydig cells and the expression of cell cycle-related biomarkers were reduced, and the ratio of cells undergoing apoptosis was also significantly raised. Intriguingly, latter findings were coherent with the relative expression of pro-apoptosis-related markers [54]. Interestingly, maternal dietary Se supplementation (0.5, 2.0 mg Se/kg dry matter (DM)) led to the statistically higher indices related to testes histomorphology, and significantly higher (compared to the controls) density of spermatogenic cell lines and Leydig cells was recorded in testicular parenchyma of young “Taihang Black” male goats [56]. In contrast, these indices (testicular weight and volume) were significantly decreased in Se-excess (4.0 mg/kg) group. Improved levels of testosterone in testicular tissue and serum, and increased expression of testosterone biosynthesis-related biomarkers were observed in Se-treated groups. The relative mRNA expression of StAR, 3β-HSD and cytochrome P450 family 11 subfamily A member 1 (CYP11A1) was reduced when the level of Se was increased in the maternal diet. Supplementation of Se in maternal diet could also influence the expression of androgenic receptor protein in testes of resulting progeny [56]. Furthermore, in an interesting comparative transcriptomic study on testes of new born bovine calves, Cerny and colleagues [57] reported that the form of Se (35 ppm organic Se, 35 ppm inorganic Se, and 50:50 mix of both (MIX)) fed to cows during gestation differentially impacted the expression of several mRNAs putatively regulating the encoding proteins implicated in steroidogenesis and/or spermatogenesis. Relatively pronounced and desirable transcriptional expression of mRNAs implicated in steroidogenesis and spermatogenesis were observed compared to the calves born to cows fed inorganic Se during gestation [57]. Of note, testes of neonatal calves born to cows fed MIX (50:50 organic and inorganic Se) showed an independent transcriptomic phenotype that was not intermediate between the organic and inorganic groups [57]. It is worthwhile to mention that all claves used in this analysis were of high/adequate Se status and were born to the dams of high/adequate Se status. However, at present, the mechanistic and physiological bases of these differential transcriptomic events are incompletely understood and future well- powered studies with adequate number of animals are awaited. Nevertheless, previously it has been demonstrated that organic Se resulted in the higher blood and liver tissue levels of Se compared to the inorganic supplemented cows [57][58][59].
Similar to its role in steroidogenesis, Se has also been implicated to play an important role in spermatogenesis. In one of the classical studies on the role Se in male fertility and reproduction, Watanabe and Endo [60] examined the morphology of sperm and spermatocyte chromosomes in mice fed Se-deficient diet. They reported that the ratio of abnormal sperm was high (6.8% to 49.6%) in Se-deficient group compared to the control group (4.0% to 15.0%). The morphological defects were more pronounced in sperm head compared to other regions i.e., midpiece and tail. However, the frequency of chromosomal abnormalities in spermatocytes (MI stage) was comparable between Se-insufficient and the control groups [60]. Besides, in another study, Se-deficient (yeast-based Se 0.02 ppm) feed significantly reduced the number of spermatogenic cell line i.e., pachytene spermatocytes, spermatids and maturing sperm in mice [61]. Taken together, these findings corroborate that Se might have strong implication in the process of spermatogenesis in males.
Recently, in an in vitro cell culture study on Sertoli cells of newborn bovine calves, it has been shown that Se supplementation (inorganic Se; 0.25, 0.50, 0.75, and 1.00 mg/L) in culture media could influence the cell viability and expression of essential protein components (occludin, connexin-43, zonula occluden, E-cadherin) of blood–testis-barrier. In fact, this effect of Se was dose-dependent and exhibited the U-shaped response i.e., optimal ameliorative effects were observed at moderate doses (0.25, 0.50), and temporary cytotoxic effects were evident when the dose was increased to 0.75, and 1.00 mg/L [62]. Interestingly, the mechanistic basis for these effects of Se was later elucidated by the same group of researchers [63], where they demonstrated that Se-pretreatment could ameliorate microcystin-LR (MC-LR)-triggered cytotoxicity in bovine Sertoli cells. Pre-treatment with inorganic Se (0.50 mg/L) inhibited the nuclear factor kappa B (NFκB) activation in cultured Sertoli cells. Selenium also exhibited some immunomodulatory roles, which were evident by the inhibition of inflammatory cytokine activation in MC-LR-exposed Sertoli cells [63]. Besides, Se-pretreatment also modulated the expression of mitochondria-related genes such as Cytochrome c oxidase subunit I (COX-1), Cytochrome c oxidase subunit 2 (COX-2), Acetyl-CoA acetyltransferase 1 (ACAT1), Mitochondrial transcription factor A (mtTFA) and Subunit 2 of NADH dehydrogenase (MT-ND2). Se-treatment also modulated the apoptotic events in Sertoli cells, which were highlighted by the inhibitory effects on apoptosis induction via cytochrome-c release and expression of caspase-3. Mitophagy and mislocalization of components of blood–testis-barrier were also inhibited in Se-treated Sertoli cells compared to the MC-LR-exposed group. Intriguingly, the peculiar antioxidant effects of Se were also observed, which were evident by an increased GPX4 activity in cultured Sertoli cells [63]. Since, NFκB and mitochondrial signaling pathways, and blood–testis barrier (an essential measure for protecting the testicular parenchyma) are implicated in delicate cellular functions in testicular parenchyma, results of these studies reasonably provide the novel and enticing evidence that Se, at least in part, could help ameliorate the Sertoli cells-related perturbations is spermatogenesis and male fertility. Nevertheless, both in vitro and in vivo studies involving other animal models will be interesting, and have the potential to provide concreate evidence with regards to ameliorative roles of Se in Sertoli cell function and spermatogenesis.
There is also an evidence (in vivo) that Se might have an implication in modulating the process of spermatogenesis via regulation of expression of genes related to cell cycle and apoptosis [64][65][66]; however, the precise mechanistic basis for this Se-mediated modulation is still unclear [54]. It has been demonstrated that components (cjun and cfos) of redox active transcription factor, Activator protein 1 (AP-1), modulate the proliferation and differentiation of cells, and they are also implicated in modulating the signal transduction pathway, and also play a regulatory role in steroidogenesis and spermatogenesis. It has also been shown that the expression of transcription factor (AP-1), cfos and cjun, is stage specific and increased during the phase of active spermatogenesis [67]. It was reported that when mice were fed a Se-deficient diet (yeast-based diet with 0.02 ppm Se), a significant decline in testis Se-levels was observed compared to those who received a Se-adequate (0.2 ppm inorganic Se) diet, and a significant increase was observed in Se-excess group (1 ppm in organic Se) compared to the Se-adequate and Se-deficient groups. GPX activity and lipid peroxidation (LPO) levels were also decreased and increased, respectively in Se-deficient group. Interestingly, a significant increase was also observed in the mRNA and protein expression of cfos and cjun in Se-deficient mice [67]. Of note, 8-weeks-long Se-deficiency lead to a significant reduction in the number of spermatogenic cells i.e., pachytene spermatocytes, late and mature spermatids. Intriguingly, the effect of Se-deficiency was less intense on germ cell kinetics at the early stages, suggesting a potential implication of Se-deficiency in affecting the overall sperm quality parameters observed in other similar studies [67]. It has been suggested that loss of AP-1 (cJun/cFos) expression during the meiotic stages might have an involvement in preventing the progression of these gem cells to differentiate into spermatids [68]. It has also been demonstrated that Se-deficiency, and to a lesser degree, Se-excess perturb these regulatory processes. As for the impact of high Se levels on modulation of AP-1, it has been suggested that Se at higher levels react with GSH and thiols present in proteins, contributing to the constitution of -S-S- and GS-Se-SG. Since the regulation of Fos and Jun is mediated by the redox status of a critical cysteine residue, thus the oxidation by excessive levels of Se may preclude the binding of cJun homodimers at the promoter sites and may lead towards the decreased cjun mRNA [68]. In addition, mitogen-activated protein kinase (MAPK) has been implicated in modulating cfos gene expression, and high Se levels have been linked with inhibition of MAPK/c-Jun amino-terminal kinase (JNK) pathways. Therefore, these two mechanisms are suggested to have an involvement in decreasing the expression of cjun and cfos in condition of excess Se [61]. Conversely, the reduced expression of these transcription factors in limiting Se conditions may be attributed to the increased ROS levels and subsequent oxidative damage at the promoter sites in these transcription factors, since the binding to AP-1 control sequence is reduced in Se-deficient conditions [68].
Similarly, NFκB, a widely known redox-regulated transcription factor reportedly performs an important function in spermatogenesis. It has been shown that an increased relative expression of both p65 and p50 genes (components of NFκB) was observed in male mice consuming Se-deficient diet (0.02 ppm inorganic Se) [66]. Besides, the levels of inducible nitric oxide and expression of inhibitory protein IκB were increased and decreased, respectively in Se-deficient mice [66]. Previously, it has also been suggested that during conditions of oxidative stress in testis, NFκB proteins paly a pro-apoptotic role in spermatogenic cells, which elicits the hypothesis that an increased expression of NFκB might lead to an exuberant germ cell death and reduced fertility. Therefore; these findings clearly demonstrate that the decreased Se supply has potential negative implication on reproductive efficiency and spermatogenesis in mice via modulating the expression and activation of NFκB in testes [66]. Intriguingly, it has been demonstrated that Se-mediated oxidative stress also has a potential implication in the modulation of expression of HSP70, HSP70-2, and MSJ-1 (a chaperone partner of spermatogenic cell-specific HSP70-2), contributing to an impaired spermatogenesis and reduced fertility in mice [69]. Previously it has also been demonstrated that the alterations in Se-levels (deficiency or excess) could lead to significantly increased apoptosis (p53-meidated) in spermatogenic cells in mice [65]. Therefore, these findings and others described before lend reasonable insights and add new dimensions to the understanding of underlying Se-mediated molecular mechanisms implicated in impairing the process of spermatogenesis and male fertility. However, the exact mechanisms behind such detrimental effects of alterations in Se-levels are still incompletely understood and require further elucidation.
In light of available evidences, it could be inferred that Se-deficiency has a strong implication in modulation of redox signaling and intracellular oxidative stress, triggering the expression of several redox-sensitive transcription and proliferation factors, and ultimately affecting the process of spermatogenesis and male fertility (Figure 1). Therefore, well-powered studies focusing on the elucidation of Se-mediated modulation of transcription and proliferation factors should be continued in future. Besides, it is also envisaged that like GPX4 and SELENOP, other selenoproteins may also have certain complementary roles in steroidogenesis and spermatogenesis in males. For instance, in situ hybridization experiments have demonstrated high levels of Selenov mRNA in seminiferous tubules in mice, but its exact function in spermatogenesis still remains largely unexplored. Recently it has been shown that selenoprotein U (absent in mouse and human) has a potential implication in modulation of phosphatidylinositide 3-kinases–AKT–mechanistic (or mammalian) target of rapamycin (PI3K–AKT–mTOR) signaling pathway in chicken Sertoli cells (SC) and could perform a novel regulatory role in the SC-mediated spermatogenesis [70]. Therefore, similar studies focusing on the possible physiological and regulatory functions of other selenoproteins such as, selenoprotein K (SELNOK), selenoprotein F (SELENOF), selenoprotein S (SELENOS), SELENOW, and others in mammalian counterparts may also provide enticing results and improve our understanding regarding Se and selenoprotein-mediated modulation of reproductive function in mammalian males. Moreover, it has been reported that seasonal variations could also influence the production of sperm. Therefore, it is highly recommended that replication of these findings in adequately powered studies (involving higher domestic animals) are needed to gain confidence with regards to the interpretation and comparison of evidences currently at hand. Meanwhile, such studies should also focus on an adequate (multiple) number of spermatogenesis cycles for ensuring the optimal effects of Se supplementation on steroidogenesis and spermatogenesis.
Figure 1. Schematic illustrating the implication of Se-deficiency in steroidogenesis, spermatogenesis, and male fertility (For details see the text in Section 3.1).
3.2. Implication of Se on Male Fertility-Related Parameters
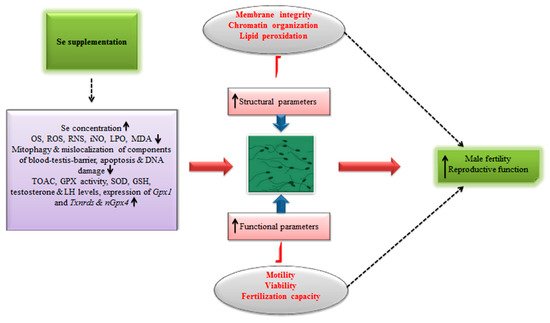
It has been reported that sperm maturation process has a strong association with the sperm and ejaculate quality, and overall reproductive efficiency in males. Therefore, any abnormality in such processes might result in ejaculates of inadequate and poorer quality and declined fertility in males [14]. Similarly, increased Se dietary intake has been implicated in enhancing the antioxidant GPX activity, thereby improving the fertility in males [15]. Meanwhile, as discussed before, an excessive supplementation could also result in no therapeutic advantage or even deleterious effects on overall reproductive outcome in males.
It is also worthwhile to mention that, even though the overproduction of ROS results in oxidative stress-induced DNA damage and/or apoptosis, membrane peroxidation and decreased sperm motility, an optimal level is necessary to carry some vital sperm functions viz. capacitation and acrosome reactions [7]. Therefore, a delicate balance in the redox regulation is necessary for the optimal functioning of cells. It has been shown that the alterations in Se-levels may perturb the redox status and could lead to oxidative stress, adversely affecting male fertility by altering the expression of biologically important markers and activity of antioxidant enzymes. To this effect, Kaushal and Bansal [69], reported that the alterations in Se-levels perturb the expression profiles of HSP70 proteins (heat shock proteins) and induce the oxidative stress. When mice were fed Se-deficient (0.02 ppm inorganic Se) and Se-excess (1.0 ppm inorganic Se) diets, a significant increase was observed in the levels of oxidative stress-related markers such as LPO, malondialdehyde (MDA) and free radicals (reactive oxygen species). Besides, a decreased level of GPX was noticed in Se-deficient mice. Conversely, Se-excess group showed relatively higher levels of GPX. Interestingly, the overall fertility-related markers were significantly diminished in both groups [69]. In another similar study, increased LPO, elevated oxidative stress, and reduced levels of GPX were observed in male mice consuming Se-deficient diet [66]. Recently, it has been demonstrated that two months long dietary supplementation of organic Se (0.3 mg per kg body weight) significantly improved the gross morphological and histomorphologcal indices in testes of young male goats. The enzymatic activity of GPX and superoxide dismutase (SOD) in serum and testicular tissue were also significantly ameliorated in young male goats treated with Se compared to the control group [71].
In addition, Stefanov and colleagues [72] studied the effects of organic (1.83g L-SeMet [Sel-Plex] per animal per day) and inorganic (4.0 mg sodium selenite per animal per day) Se supplementation on semen parameters in Bulgarian Merino rams. After 45 days of Se dietary supplementation, improvements were observed in semen volume per ejaculate, sperm motility and overall sperm survival rate in treated rams [72]. Besides, in a very recent study on a mouse model, Asri-Rezaei and colleagues [73] observed that an intraperitoneal injection of sodium selenite (0.50 mg per kg body weight) and Se nanoparticles (0.50 mg per kg body weight) for seven consecutive days adequately improved the tissue Se concentration in testis (as observed on day 28 post-injection). Similarly, the enzymic activities of antioxidant biomarkers were also significantly improved following Se treatment. In addition, the sperm quality parameters such as total count and motility were also improved compared to the control group [73]. In general, these results corroborate that Se supplementation can produce beneficial effects and counter the oxidative stress.
Besides, recently in an interesting cost–benefit analysis [74], it has been shown that boars fed organic Se-based (0.5 mg per kg) diet produced around 23% higher doses of semen per week compared to those maintained on inorganic Se diets (0.5 mg per kg). A significant increase in revenue (26%) was also observed in this study [74]. In any case, additional animal studies reporting the effects of Se supplementation on the male reproductive efficiency in different animal models are presented in Table 2. Meanwhile, Figure 2 illustrates the implication of Se supplementation in ameliorating the male fertility and reproduction.
Figure 2. Schematic illustrating the implication of Se supplementation in ameliorating fertility and reproductive efficiency in males.
Table 2. Animal studies reporting the effects of selenium supplementation on male reproductive efficiency.
| Model | Treatment | Key Observations Reported | Ref. |
|---|---|---|---|
| Sprague-Dawley rats | Se nanoparticles at supranutritional levels (0.2, 0.4, or 0.8 mg Se per kg body weight) | Sperm parameters such as, sperm concentration, motility, and morphological features were all improved at supranutritional levels. However, these parameters were significantly affected when rats were supplemented with higher levels (nonlethal level) of Se nanoparticles i.e., at 2.0, 4.0, or 8.0 mg Se per kg body weight. | [75] |
| Sprague-Dawley rats | Treated with inorganic Se [0.01(deficient); 0.25 (adequate); 3 (excess); or 5 (excess) mg per kg] for four weeks | The U-shaped response of dietary Se was observed on DNA damage and sperm quality. Se deficiency showed a lower expression of sensitive antioxidant selenoproteins (Gpx1 and Txnrds). However, excessive doses of Se impaired sperm quality and this was linked with reduced mRNA expression of nGpx4. | [76] |
| Mouse | Se-supplement (inorganic Se (0.3 μg/g Se) or organic Se-enriched probiotics (containing 0.3 μg/g Se) given for 75 days | Organic Se co-supplemented with probiotics significantly improved male fertility in mice. The ameliorated fertility index included the parameters such as, reduced testicular tissue injury, increased levels of serum testosterone, and improved sperm indices in Se-supplemented group. As such, these improved fertility-related parameters were ascribed to be the result of the antioxidant function of Se. | [77] |
| Mouse | 0.2 ppm sodium selenite; 1.0 ppm sodium selenite |
Mice in both groups showed an increased occurrence of mitochondria- and plasma membrane-related defects, and DNA damage in sperm. However, these damages were more pronounced in mice exposed to Se-deficient feed. | [78] |
| Mouse | Se-deficient diet (0.02 ppm) Se-sufficient (0.2 ppm); organic Se |
Sperm from Se-deficient mice demonstrated vitiated chromatin condensation, declined in vitro fertilization ability and increased lipid peroxidation (LPO) in both testes and sperm compared to the Se-sufficient mice. | [79] |
| Mouse | Se-deficient (0.02 ppm) Se-excess (0.2 ppm); yeast-based Se. Mice were fed for 4 months |
Se concentration and GPX activity (in testis) were significantly reduced. The fertility percentage and size of litter were both reduced in Se-deficient group. | [61] |
| Aged mice | Inorganic Se 0.2 mg/kg body weight | Improved sperm parameters and increased expression of CatSper genes were observed in Se-treated group. | [80] |
| Rabbit | Treated with Se nanoparticles (400 μg/kg) for 60 days | Improved serum testosterone levels were recorded in Se-treated group compared to the control. Besides, improved ejaculate volume and sperm quality parameters such as, sperm morphology, viability were observed. | [81] |
| Ram | 0.5 ppm organic Se; 0.2 ppm organic Se | A significantly higher concentration of Se and improved ejaculate and sperm quality were observed in seminal plasma of rams exposed to a feed containing 0.5 ppm organic Se compared to those who received 0.2 ppm organic Se. | [82] |
| Boar | Organic Se (0.2 mg per kg); Inorganic Se (0.2 mg per kg) |
Ejaculate quality and sperm parameters were significantly improved in boars following dietary supplementation of organic Se (0.2 mg per kg) compared to those treated with sodium selenite at the same dose. | [83] |
| Aardi buck | Sodium selenite 0.1 mg/kg, Sodium selenite 0.05 mg/kg |
Improved sperm count and motility was observed in both Se-treated groups. However, relatively better outcomes were observed in 0.1 mg/kg group. | [84] |
| Boar | 0.5 ppm organic Se | Following 11 weeks of feeding trail, organic Se supplementation increased glutathione peroxidase 4 (GPX4) activity (raw semen) and number of seminal doses in boars. | [85] |
| Boar | 0.3 ppm organic Se; 0.3 ppm inorganic Se |
Following 12 weeks of Se supplementation, Se content and GPX activity were increased in semen of boars treated with organic and inorganic Se. Besides, semen quality parameters namely semen concentration and progressive motility of sperm were improved compared to the control group without Se. Improved resistance of liquid stored semen to hypo-osmotic shock and thermal tests, and improved fertility rates were observed in semen of boars treated with Se. All mentioned indices were slightly higher in the organic Se group compared to the inorganic group. | [86] |
| Buffalo bulls | 10 mg organic Se/animal twice a week; and 10 mg inorganic Se/animal twice a week |
Three months long Se supplementation significantly improved the sperm quality parameters (ejaculate volume, sperm motility, concentration, and morphology) in buffalo bulls. Besides, testosterone concentrations were also increased in Se-treated groups. | [87] |
| Saanen bucks | Inorganic Se 0.34 mg/kg body weight supplemented at ten-day intervals for three months | Se supplementation improved the testicular biometry and sperm parameters. GPX activity, plasma testosterone and LH levels significantly increased in Se-treated group from days 40 to 80 compared to the control group. These indices reached peak reached peak at day 80 of the trial. | [88] |
| Bovine bull | In vitro fertilization (IVF) medium supplemented with Se (100 ng/mL) | A significant increase in sperm mitochondrial activity was observed after 1 h of incubation in Se-supplemented IVF medium. Moreover, Se supplementation after 2 h of incubation showed an increase in HOST-positive (hypo-osmotic swelling test) sperm and sperm acrosome integrity. Increased number of sperm bound to zona pellucida (ZP) was observed in Se-treated group compared to the control. | [89] |
It is reiterated that sperm quality parameters such as, sperm concentration, vitality, progressive movement, and overall sperm morphology are regarded as the essential clinical markers for the assessment of reproduction efficiency. Therefore, perturbations in these markers may lead towards sub- and infertility in males. Of note, oxidative stress has been implicated as an important factor contributing towards male infertility [90]. Intriguingly, it is believed that sperm are more sensitive to oxidative damages; this is largely because of the biochemical composition of sperm i.e., it contains higher ratio of polyunsaturated fatty acids and low concentrations of cytoplasmic antioxidant enzymes compared to the somatic cells [91]. Therefore, in conditions of increased ROS-induced oxidative stress, the plasma membrane integrity is affected due to the triggering of LPO cascades. Similarly increased oxidative stress is also implicated in inducing DNA damage in sperm by causing perturbations in chromatin condensation, and the process of chromatin condensation is an essential step for both sperm maturation and fertilization capacity [79]. In this connection, it has been demonstrated that Se deficiency could lead to the impaired chromatin condensation and reorganization processes via elicitation of oxidative stress, and lead to the impaired sperm quality and reduced fertilization capacity in males [79]. Nevertheless, in addition to aforementioned observations of previous studies, further studies focusing on the elucidation of potential markers of DNA integrity will be of high value and should lend encouraging insights with regards to the role of Se in process of spermatogenesis and overall male fertility. Besides, in-depth evaluation of sperm chromatin condensation and reorganization is also biologically more relevant because it is completely reorganized during the later phases of spermatogenesis when histones are substituted by protamines [79]. Further adequately powered studies on Se-deficient models would also improve our understanding in this domain. As demonstrated in previous studies, Se deficiency could impair the sperm quality parameters and antioxidant status in the male reproductive organs, therefore it is reasonable to infer that Se-deficiency potentially perturbs the biosynthesis of selenoproteins having essential roles in redox regulation, leading towards oxidative insult and resultant reduced male fertility parameters. Therefore, in addition to already known mechanisms, more focus should be centered on elucidation of other putative mechanisms implicated in impairing the selenoprotein-mediated redox homeostasis in sperm/ejaculates.
The results of relevant studies with regards to the form of Se used are comparable and encouraging i.e., it is fairly evident that both organic and inorganic forms have the potential to ameliorate male fertility parameters in different animal models (also see Table 2). In order to get a more precise picture, it is also reasonable to conduct more adequately powered studies focusing on the comparative ameliorative effects of different forms of Se. It should also be considered that the effects of Se (especially the organic form) might be impacted by the duration of supplementation; therefore, for obtaining the desirable outcomes with regards to the improved fertility in different animal species, future studies should also include this aspect within their scope. Se-nanoparticles are also reported to possess more bioavailability and less toxicity potential compared to other conventional forms of Se. However, the level of evidence regarding the effects Se-nanoparticles in ameliorating male fertility is still insufficient, and more multifaceted studies are awaited to lend the concrete evidence.
Selenium in Seminal Plasma and its Implication in Male Fertility
Seminal plasma is considered to play many essential roles in motility, viability and sustentation of fertilizing ability of mammalian sperm [92]. Villaverde et al. [50] studied the correlation between the serum concentrations of Se and sperm quality traits, testosterone levels, and testes morphology in domestic cats. Se concentrations showed no influence on testosterone levels and sperm production. However, Se concentrations in blood and seminal plasma showed a correlation with sperm quality parameters. It was demonstrated that Se concentrations in seminal plasma were negatively correlated with total testicular weight and sperm morphological traits such as total head defects. Besides, Se concentrations in serum were also negatively correlated with some sperm motility-related parameters including average path velocity, straight line velocity, and curvilinear velocity [50]. These observations, to some extent, highlight that Se (in seminal plasma) may have a potential implication in male fertility and could be used as an important marker in ameliorating the overall male reproductive biology of domestic cats [50]. However, it should be noted that a small number of cats (n = 6) was used in this investigation; therefore, inclusion of Se concentration (as a biomarker) in studies focusing on evaluation of potential association between this trace element and sperm quality and fertility in cats should be very cautiously assessed. In addition, for getting reasonable evidences, more data should be gathered from well-powered functional/mechanistic studies using the cat models. Similarly, Bertelsmann and colleagues [93] determined the levels of Se in semen, seminal plasma and sperm, and their association with overall sperm quality and fertility in male horse. It was reported that Se level/concentration in sperm was correlated with sperm quality parameters such as, membrane integrity, progressive motility of sperm, positive acrosomal status and pregnancy rate per estrus cycle. Moreover, Se was also linked with ameliorated sperm quality and fertility in horse. These findings therefore suggest that evaluation of an optimal Se status for equine male reproduction necessitates the analysis of Se in sperm [93].
It is worthwhile to mention that Se in seminal plasma originates from secretion of glandular epithelium of accessory sex glands i.e., prostate gland, seminal vesicles, and epididymis, therefore, alterations in seminal plasma Se-levels indicate that supply of Se to these sex glands changes with dietary Se. It was also demonstrated that effects of Se on sperm motility are likely arbitrated by secretions from these sex glands, either during the process of sperm maturation or at the time of ejaculation [94]. Therefore, it is also reasonable to conduct more research on elucidation of potential functions of Se in seminal plasma and accessory sex glands. This will also improve our current understanding regarding underlying mechanisms and physiological basis of relationship between seminal plasma-Se concentrations and male fertility.
3.3. Combinatorial Effects of Se (as a Part of Micronutrient Supplement) on Male Fertility Outcomes (Animal Studies)
Recently, very few studies have demonstrated that Se in combination with other trace minerals and micronutrients can ameliorate the fertility-related outcomes in different animal species. To this effect, in one recent clinical study, Domosławska and colleagues [95] demonstrated that Se-yeast (6 μg per kg) combined with vitamin E (5 mg per kg) supplement (treated for 60 days) significantly ameliorated the semen quality parameters and antioxidant status in clinically healthy dogs with reduced fertility. In Se-supplemented group, significant improvements were observed in parameters such as, blood Se concentration, mean sperm concentration, mean total sperm count, sperm motility, and percentage of live and normal sperm. Furthermore, sperm GPX activity and total antioxidant capacity (TAOC) were also increased significantly [95]. Similar findings were also observed by the same group of researchers in 2015 [96], where the dietary supplementation of Se-yeast (6 μg per kg) in combination with vitamin E (5 mg per kg) for 60 days resulted in the improved sperm quality parameters such as total sperm count, concentration, morphology, and motility scores in clinically healthy dogs with reduced fertility [96]. Similarly, Butt et al. [97] demonstrated that Se (supplemented as 3g Selemax®) in combination with vitamin E significantly improved the ejaculate and sperm quality parameters, and increased the concentration of testosterone in Holstein Friesian bulls maintained in a hot and humid environment [97]. Additionally, Ghorbani et al. [98] evaluated the effects of dietary Se (0.3 mg/kg), Zn (40 mg/kg) and their combination on reproductive performance in Sanjabi rams during the breeding season. They reported a significant improvement in sperm concentration, total sperm count, sperm motility following a 120 days long dietary supplementation. However, the concentration of testosterone remained unaltered [98]. In another randomized, double-blinded study, Kirchhoff and colleagues [99] failed to elucidate a clear trend in the extent to which a three months long supplementation of Se/vitamin E (tablets containing 0.1 mg organic Se; capsules containing 100 mg vitamin E) influences the semen qualitative traits in normospermic Cairn Terriers. Nevertheless, an effect of supplementation (head sperm defects) and a substantial interaction between time and treatment was observed for some semen parameters such as, percentage of viable sperm, sperm head irregularities, defective acrosome, and proximal cytoplasmic droplets. Nevertheless, it is worthwhile to mention that the number of dogs (n = 9; three dogs per treatment group) used in this randomized study was too low and the subjects included were normospermic i.e., without any apparent semen quality defects. In any case, results of three recent studies (2016–2019) reporting the effects of combination treatment of Se and other micronutrients are summarized in Table 3.
Table 3. Recent animal studies reporting the implication of selenium supplementation (in combination with other micronutrients) on male fertility outcomes.
| Animal Model and Number | Treatment Regime and Duration | Key Findings | Ref. |
|---|---|---|---|
| Male CD-1 mice (n = 12 per experimental group) |
Fertilix® (CellOxess, Princeton, NJ, USA) was supplemented for two months. (Se 55 μg, zinc 7.5–11 mg, Full spectrum natural vitamin E 104–290 mg, Lycopene 7.5–15 mg Carnitine blend 200–800 mg Folic acid 400–500 mg Vitamin C 30–90 mg). |
Eight weeks long pretreatment with the antioxidant formulation completely protected oxidative stress-induced DNA damage in Gpx5 KO mice sperm. In mouse models of scrotal heat stress, only 35% (19/54) of female mice became pregnant resulting in 169 fetuses with 18% fetal resorption (30/169). Conversely, in antioxidant pretreated group 74% (42/57) of female mice became pregnant, resulting in 427 fetuses with 9% fetal resorption (38/427). | [91] |
| Four infertile male dogs with low blood Se levels (86.0–165.0 μg/L) | Organic Se 0.6 mg/kg and vitamin E (5 mg/kg) orally supplemented for 60 days. | Treated dogs showed improved sperm parameters. Increase in blood Se concentration (401 μg/L) was observed at the end of trial. When these dogs were used for matting purpose, bitches successfully conceived and gave birth to 4–6 pups. | [100] |
| Sixteen healthy normospermic dogs (two patients were excluded after adaptation period) | A supplement comprising of Se 0.27 mg/kg vitamin E 250 mg/kg, vitamin B9 1.5 mg/kg, zinc 180 mg/kg, and n-3 PUFA 0.5%, given for 90 days. | In treated group, sperm quality parameters i.e., total sperm count, concentration, sperm vitality and membrane integrity were significantly improved compared to the control group. | [101] |
In general, the results of these studies are encouraging and suggest that Se in combination with other essential micronutrients could improve the reproductive efficiency in males. However, the level of concrete evidence is still insufficient and somehow inconsistent. Therefore, well-powered randomized studies will be of high value for building the solid scientific evidence in this regard. It should also be noted that these ameliorative effects of integrated mixture of essential micronutrients might be a result of augmentative and camouflaged effects of each essential micronutrient [101]. Currently, the underlying mechanisms of such combinatorial effects are largely unclear and should be considered in future investigations [35]. Nevertheless, such studies will also lend reasonable basis for determining the proper and adequate remedy protocols aimed at ameliorating male factor sub- and infertility in mammalian species, particularly in domestic and companion animals.
3.4. Selenium and Sperm Cryopreservation
Despite significant progress made towards optimizing the cryopreservation technology, confounding factors, and underlying mechanisms related to cryoinjury and freeze tolerance of mammalian sperm remain incompletely understood. Cryopreservation is believed to result in structural and functional losses, and could also lead to apoptosis and reduced fertilization rates. Recently, it has been demonstrated that cryopreservation leads to the differential expression of several mRNAs, long non-coding RNAs, and microRNAs implicated in various essential signaling pathways such as, PI3K-AKT, p53, cAMP, cell adhesion, MAPK, calcium signaling pathways, environment stimuli, apoptosis, and metabolic activities [102][103]. These detrimental effects are linked to the excessive production of ROS and subsequent oxidative insult leading to the overall reduced sperm quality and fertilization rates. Use of antioxidants in semen extenders has been reported to ameliorate these damages and improve IVF outcomes to some extent; however, much remains to be studied in this regard.
One recent study [104] reported that when Se (2 mM) was added in semen extender, sperm quality parameters such as, motility, concentration, and membrane integrity were significantly improved following freezing-thawing of buffalo bull semen. Surprisingly, authors have reported that conception rate was significantly higher (60% vs. 30%) in buffaloes which were subsequently inseminated with Se-treated semen [104]. However, it should be noted that this study seemed to be relatively weakly designed and supporting data do not lend reasonable evidence with respect to conception rate following insemination. Therefore, there is a definitive need to validate these findings in well controlled and properly powered studies in future. In comparison, the study of Khalil et al. [105] seems to be more powered and results are more reasonable. These authors supplemented the semen extender (Tris-yolk fructose) with Se nanoparticles at different doses, and report that 1.0 mg/mL Se supplementation adequately improved the semen and sperm quality parameters including membrane integrity and sperm viability post-thawing in Holstein bulls. Besides, antioxidant biomarkers (in seminal plasma) such as TAOC and MDA were also significantly ameliorated in Se-treated group compared to the control. The rate of apoptosis and necrosis in sperm was also significantly reduced in two Se-treated groups (0.5 and 1.0 mg/mL), however, the lowest rates were observed in the 1.0 mg/mL Se group. Conversely, a higher dose of Se (1.5 mg/mL) resulted in some deleterious effects on sperm quality parameters and increased the ratio of apoptosis in sperm. Interestingly, the conception rate was significantly improved (90% vs. 59%) when cows were inseminated with cryopreserved semen from Se-treated (1.0 mg/mL) group [105]. Albeit, the quality and level of such evidence is still low, these findings, to some degree, support the notion that Se supplementation, particularly in nano-form, could be integrated in sperm cryopreservation protocols, and subsequently, improved fertility rates could be achieved in farm animals following insemination with cryopreserved semen.
References
- Oldfield, J.E. The two faces of selenium. J. Nutr. 1987, 117, 2002–2008.
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293.
- O’Dell, B.L.; Sunde, R.A. Handbook of Nutritionally Essential Mineral Elements; CRC Press: New York, NY, USA; Basel, Switzerland; Hong Kong, China, 1997.
- Ceko, M.J.; O’leary, S.; Harris, H.H.; Hummitzsch, K.; Rodgers, R.J. Trace elements in ovaries: Measurement and physiology. Biol. Reprod. 2016, 94, 86.
- Xiong, X.; Lan, D.; Li, J.; Lin, Y.; Li, M. Selenium supplementation during in vitro maturation enhances meiosis and developmental capacity of yak oocytes. Anim. Sci. J. 2018, 89, 298–306.
- Hedaoo, M.; Khllare, K.; Meshram, M.; Sahatpure, S.; Patil, M. Study of some serum trace minerals in cyclic and non-cyclic surti buffaloes. Vet. World 2008, 1, 71.
- Mirone, M.; Giannetta, E.; Isidori, A. Selenium and reproductive function. A systematic review. J. Endocrinol. Investig. 2013, 36, 28–36.
- Pappas, A.; Zoidis, E.; Surai, P.; Zervas, G. Selenoproteins and maternal nutrition. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 361–372.
- Foresta, C.; Flohé, L.; Garolla, A.; Roveri, A.; Ursini, F.; Maiorino, M. Male fertility is linked to the selenoprotein phospholipid hydroperoxide glutathione peroxidase. Biol. Reprod. 2002, 67, 967–971.
- Kommisrud, E.; Østerås, O.; Vatn, T. Blood selenium associated with health and fertility in Norwegian dairy herds. Acta Vet. Scand. 2005, 46, 229.
- Behne, D.; Höfer, T.; von Berswordt-Wallrabe, R.; Elger, W. Selenium in the testis of the rat: Studies on its regulation and its importance for the organism. J. Nutr. 1982, 112, 1682–1687.
- Behne, D.; Weiler, H.; Kyriakopoulos, A. Effects of selenium deficiency on testicular morphology and function in rats. J. Reprod. Fertil. 1996, 106, 291–297.
- Flohe, L. Selenium in mammalian spermiogenesis. Biol. Chem. 2007, 388, 987–995.
- Ahsan, U.; Kamran, Z.; Raza, I.; Ahmad, S.; Babar, W.; Riaz, M.; Iqbal, Z. Role of selenium in male reproduction—A review. Anim. Reprod. Sci. 2014, 146, 55–62.
- Irvine, D.S. Glutathione as a treatment for male infertility. Rev. Reprod. 1996, 1, 6–12.
- Conrad, M.; Schweizer, U. Mouse Models that Target Individual Selenoproteins. In Selenium: Its Molecular Biology and Role in Human Health; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 567–578.
- Riese, C.; Michaelis, M.; Mentrup, B.; Gotz, F.; Kohrle, J.; Schweizer, U.; Schomburg, L. Selenium-dependent pre-and posttranscriptional mechanisms are responsible for sexual dimorphic expression of selenoproteins in murine tissues. Endocrinology 2006, 147, 5883–5892.
- Beckett, G.J.; Arthur, J.R. Selenium and endocrine systems. J. Endocrinol. 2005, 184, 455–465.
- Knapen, M.F.; Zusterzeel, P.L.; Peters, W.H.; Steegers, E.A. Glutathione and glutathione-related enzymes in reproduction: A review. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 82, 171–184.
- Ursini, F.; Heim, S.; Kiess, M.; Maiorino, M.; Roveri, A.; Wissing, J.; Flohé, L. Dual function of the selenoprotein PHGPx during sperm maturation. Science 1999, 285, 1393–1396.
- Safarinejad, M.R.; Safarinejad, S. Efficacy of selenium and/or N-acetyl-cysteine for improving semen parameters in infertile men: A double-blind, placebo controlled, randomized study. J. Urol. 2009, 181, 741–751.
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286.
- Conrad, M.; Moreno, S.; Sinowatz, F.; Ursini, F.; Kölle, S.; Roveri, A.; Brielmeier, M.; Wurst, W.; Maiorino, M.; Bornkamm, G. The nuclear form of phospholipid hydroperoxide glutathione peroxidase is a protein thiol peroxidase contributing to sperm chromatin stability. Mol. Cell. Biol. 2005, 25, 7637–7644.
- Chabory, E.; Damon, C.; Lenoir, A.; Henry-Berger, J.; Vernet, P.; Cadet, R.; Saez, F.; Drevet, J. Mammalian glutathione peroxidases control acquisition and maintenance of spermatozoa integrity 1. J. Anim. Sci. 2010, 88, 1321–1331.
- Schneider, M.; Förster, H.; Boersma, A.; Seiler, A.; Wehnes, H.; Sinowatz, F.; Neumüller, C.; Deutsch, M.J.; Walch, A.; de Angelis, M.H. Mitochondrial glutathione peroxidase 4 disruption causes male infertility. FASEB J. 2009, 23, 3233–3242.
- Imai, H.; Suzuki, K.; Ishizaka, K.; Ichinose, S.; Oshima, H.; Okayasu, I.; Emoto, K.; Umeda, M.; Nakagawa, Y. Failure of the expression of phospholipid hydroperoxide glutathione peroxidase in the spermatozoa of human infertile males. Biol. Reprod. 2001, 64, 674–683.
- Imai, H.; Hakkaku, N.; Iwamoto, R.; Suzuki, J.; Suzuki, T.; Tajima, Y.; Konishi, K.; Minami, S.; Ichinose, S.; Ishizaka, K. Depletion of selenoprotein GPx4 in spermatocytes causes male infertility in mice. J. Biol. Chem. 2009, 284, 32522–32532.
- Meseguer, M.; Maria, J.; Simón, C.; Pellicer, A.; Remohí, J.; Garrido, N. Effect of sperm glutathione peroxidases 1 and 4 on embryo asymmetry and blastocyst quality in oocyte donation cycles. Fertil. Steril. 2006, 86, 1376–1385.
- Parillo, F.; Sylla, L.; Palombi, C.; Monaci, M.; Stradaioli, G. Immunocytochemical Localisation of Phospholipid Hydroperoxide Glutathione Peroxidase in Bull’s Spermatogenic Cells. Ital. J. Anim. Sci. 2014, 13, 3483.
- Olson, G.E.; Winfrey, V.P.; NagDas, S.K.; Hill, K.E.; Burk, R.F. Apolipoprotein E receptor-2 (ApoER2) mediates selenium uptake from selenoprotein P by the mouse testis. J. Biol. Chem. 2007, 282, 12290–12297.
- Hill, K.E.; Zhou, J.; McMahan, W.J.; Motley, A.K.; Atkins, J.F.; Gesteland, R.F.; Burk, R.F. Deletion of selenoprotein P alters distribution of selenium in the mouse. J. Biol. Chem. 2003, 278, 13640–13646.
- Kehr, S.; Malinouski, M.; Finney, L.; Vogt, S.; Labunskyy, V.M.; Kasaikina, M.V.; Carlson, B.A.; Zhou, Y.; Hatfield, D.L.; Gladyshev, V.N. X-ray fluorescence microscopy reveals the role of selenium in spermatogenesis. J. Mol. Biol. 2009, 389, 808–818.
- Noblanc, A.; Kocer, A.; Chabory, E.; Vernet, P.; Saez, F.; Cadet, R.; Conrad, M.; Drevet, J.R. Glutathione peroxidases at work on epididymal spermatozoa: An example of the dual effect of reactive oxygen species on mammalian male fertilizing ability. J. Androl. 2011, 32, 641–650.
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268.
- Qazi, I.H.; Angel, C.; Yang, H.; Pan, B.; Zoidis, E.; Zeng, C.J.; Han, H.; Zhou, G.B. Selenium, Selenoproteins, and Female Reproduction: A Review. Molecules 2018, 23, 3053.
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241.
- Gladyshev, V.N.; Arnér, E.S.; Berry, M.J.; Brigelius-Flohé, R.; Bruford, E.A.; Burk, R.F.; Carlson, B.A.; Castellano, S.; Chavatte, L.; Conrad, M. Selenoprotein gene nomenclature. J. Biol. Chem. 2016, 291, 24036–24040.
- Zoidis, E.; Pappas, A. The health effects of selenoproteins. In Selenium: Sources, Functions and Health Effects; Aomori, C., Hokkaido, M., Eds.; Nova Science Publishers: New York, NY, USA, 2012.
- Boitani, C.; Puglisi, R. Selenium, a key element in spermatogenesis and male fertility. In Molecular Mechanisms in Spermatogenesis; Springer: Berlin/Heidelberg, Germany, 2009; pp. 65–73.
- Su, D.; Novoselov, S.V.; Sun, Q.-A.; Moustafa, M.E.; Zhou, Y.; Oko, R.; Hatfield, D.L.; Gladyshev, V.N. Mammalian selenoprotein thioredoxin-glutathione reductase roles in disulfide bond formation and sperm maturation. J. Biol. Chem. 2005, 280, 26491–26498.
- Michaelis, M.; Gralla, O.; Behrends, T.; Scharpf, M.; Endermann, T.; Rijntjes, E.; Pietschmann, N.; Hollenbach, B.; Schomburg, L. Selenoprotein P in seminal fluid is a novel biomarker of sperm quality. Biochem. Biophys. Res. Commun. 2014, 443, 905–910.
- Renko, K.; Werner, M.; Renner-Muller, I.; Cooper, T.G.; Yeung, C.H.; Hollenbach, B.; Scharpf, M.; Kohrle, J.; Schomburg, L.; Schweizer, U. Hepatic selenoprotein P (SePP) expression restores selenium transport and prevents infertility and motor-incoordination in Sepp-knockout mice. Biochem. J. 2008, 409, 741–749.
- Olson, G.E.; Winfrey, V.P.; NagDas, S.K.; Hill, K.E.; Burk, R.F. Selenoprotein P is required for mouse sperm development. Biol. Reprod. 2005, 73, 201–211.
- Turanov, A.A.; Malinouski, M.; Gladyshev, V.N. Selenium and male reproduction. In Selenium: Its Molecular Biology and Role in Human Health; Hatfield, D.L., Berry, M.J., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2012; pp. 409–417.
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443.
- Behne, D.; Duk, M.; Elger, W. Selenium content and glutathione peroxidase activity in the testis of the maturing rat. J. Nutr. 1986, 116, 1442–1447.
- Nishimura, K.; Matsumiya, K.; Tsujimura, A.; Koga, M.; Kitamura, M.; Okuyama, A. Association of selenoprotein P with testosterone production in cultured Leydig cells. Arch. Androl. 2001, 47, 67–76.
- Koga, M.; Tanaka, H.; Yomogida, K.; Tsuchida, J.; Uchida, K.; Kitamura, M.; Sakoda, S.; Matsumiya, K.; Okuyama, A.; Nishimune, Y. Expression of selenoprotein-P messenger ribonucleic acid in the rat testis. Biol. Reprod. 1998, 58, 261–265.
- Oluboyo, A.; Adijeh, R.; Onyenekwe, C.; Oluboyo, B.; Mbaeri, T.; Odiegwu, C.; Chukwuma, G.; Onwuasoanya, U. Relationship between serum levels of testosterone, zinc and selenium in infertile males attending fertility clinic in Nnewi, south east Nigeria. Afr. J. Med. Med. Sci. 2012, 41, 51–54.
- Villaverde, A.I.S.; Fioratti, E.G.; Ramos, R.S.; Neves, R.C.; Ferreira, J.C.P.; Cardoso, G.S.; Padilha, P.M.; Lopes, M.D. Blood and seminal plasma concentrations of selenium, zinc and testosterone and their relationship to sperm quality and testicular biometry in domestic cats. Anim. Reprod. Sci. 2014, 150, 50–55.
- Richards, J.; Hallford, D.; Duff, G. Serum luteinizing hormone, testosterone, and thyroxine and growth responses of ram lambs fed locoweed (Oxvtropis sericea) and treated with vitamin e/selenium. Theriogenology 1999, 52, 1055–1066.
- Shi, L.; Zhang, C.; Yue, W.; Shi, L.; Zhu, X.; Lei, F. Short-term effect of dietary selenium-enriched yeast on semen parameters, antioxidant status and Se concentration in goat seminal plasma. Anim. Feed Sci. Technol. 2010, 157, 104–108.
- Ren, X.-M.; Wang, G.-G.; Xu, D.-Q.; Luo, K.; Liu, Y.-X.; Zhong, Y.-H.; Cai, Y.-Q. The protection of selenium on cadmium-induced inhibition of spermatogenesis via activating testosterone synthesis in mice. Food Chem. Toxicol. 2012, 50, 3521–3529.
- Shi, L.; Song, R.; Yao, X.; Ren, Y. Effects of selenium on the proliferation, apoptosis and testosterone production of sheep Leydig cells in vitro. Theriogenology 2017, 93, 24–32.
- Liu, H.; Li, X.; Qin, F.; Huang, K. Selenium suppresses oxidative-stress-enhanced vascular smooth muscle cell calcification by inhibiting the activation of the PI3K/AKT and ERK signaling pathways and endoplasmic reticulum stress. JBIC J. Biol. Inorg. Chem. 2014, 19, 375–388.
- Shi, L.; Song, R.; Yao, X.; Duan, Y.; Ren, Y.; Zhang, C.; Yue, W.; Lei, F. Effects of maternal dietary selenium (Se-enriched yeast) on testis development, testosterone level and testicular steroidogenesis-related gene expression of their male kids in Taihang Black Goats. Theriogenology 2018, 114, 95–102.
- Cerny, K.; Garbacik, S.; Skees, C.; Burris, W.; Matthews, J.; Bridges, P. Gestational form of selenium in free-choice mineral mixes affects transcriptome profiles of the neonatal calf testis, including those of steroidogenic and spermatogenic pathways. Biol. Trace Elem. Res. 2016, 169, 56–68.
- Brennan, K.M.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Selenium content in blood fractions and liver of beef heifers is greater with a mix of inorganic/organic or organic versus inorganic supplemental selenium but the time required for maximal assimilation is tissue-specific. Biol. Trace Elem. Res. 2011, 144, 504–516.
- Liao, S.F.; Brown, K.R.; Stromberg, A.J.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Dietary supplementation of selenium in inorganic and organic forms differentially and commonly alters blood and liver selenium concentrations and liver gene expression profiles of growing beef heifers. Biol. Trace Elem. Res. 2011, 140, 151–169.
- Watanabe, T.; Endo, A. Effects of selenium deficiency on sperm morphology and spermatocyte chromosomes in mice. Mutat. Res. Lett. 1991, 262, 93–99.
- Kaur, P.; Bansal, M.P. Effect of selenium-induced oxidative stress on the cell kinetics in testis and reproductive ability of male mice. Nutrition 2005, 21, 351–357.
- Adegoke, E.; Wang, X.; Wang, H.; Wang, C.; Zhang, H.; Zhang, G. Selenium (Na 2 SeO 3) Upregulates Expression of Immune Genes and Blood–Testis Barrier Constituent Proteins of Bovine Sertoli Cell In Vitro. Biol. Trace Elem. Res. 2018, 185, 332–343.
- Adegoke, E.; Xue, W.; Machebe, N.; Adeniran, S.; Hao, W.; Chen, W.; Han, Z.; Guixue, Z.; Peng, Z. Sodium Selenite inhibits mitophagy, downregulation and mislocalization of blood–testis barrier proteins of bovine Sertoli cell exposed to microcystin-leucine arginine (MC-LR) via TLR4/NF-kB and mitochondrial signaling pathways blockage. Ecotoxicol. Environ. Saf. 2018, 166, 165–175.
- Kaushal, N.; Bansal, M. Inhibition of CDC2/Cyclin B1 in response to selenium-induced oxidative stress during spermatogenesis: Potential role of Cdc25c and p21. Mol. Cell. Biochem. 2007, 298, 139–150.
- Kaushal, N.; Bansal, M. Selenium variation induced oxidative stress regulates p53 dependent germ cell apoptosis: Plausible involvement of HSP70-2. Eur. J. Nutr. 2009, 48, 221–227.
- Shalini, S.; Bansal, M. Alterations in selenium status influences reproductive potential of male mice by modulation of transcription factor NFκB. Biometals 2007, 20, 49–59.
- Shalini, S.; Bansal, M.P. Role of selenium in spermatogenesis: Differential expression of cjun and cfos in tubular cells of mice testis. Mol. Cell. Biochem. 2006, 292, 27–38.
- Shalini, S.; Bansal, M. Role of selenium in regulation of spermatogenesis: Involvement of activator protein 1. Biofactors 2005, 23, 151–162.
- Kaushal, N.; Bansal, M. Diminished reproductive potential of male mice in response to selenium-induced oxidative stress: Involvement of HSP70, HSP70-2, and MSJ-1. J. Biochem. Mol. Toxicol. 2009, 23, 125–136.
- Sattar, H.; Yang, J.; Zhao, X.; Cai, J.; Liu, Q.; Ishfaq, M.; Yang, Z.; Chen, M.; Zhang, Z.; Xu, S. Selenoprotein-U (SelU) knockdown triggers autophagy through PI3K–Akt–mTOR pathway inhibition in rooster Sertoli cells. Metallomics 2018, 10, 929–940.
- Bano, I.; Malhi, M.; Soomro, S.; Kandhro, S.; Awais, M.; Baloch, S.; Perveen, S.; Sajjad, H. Effect of Dietary Selenium Supplementation on Morphology and Antioxidant Status in Testes of Goat. J. Basic Appl. Sci. 2018, 14, 53–61.
- Stefanov, R.; Chervenkov, M.; Anev, G.; Maksimović, N.; Andreeva, M.; Ivanova, T.; Milovanović, A. Effect of supplementation with inorganic and organic selenium on sperm quality and quantity in north-east Bulgarian merino rams. Biotechnol. Anim. Husb. 2018, 34, 69–81.
- Asri-Rezaei, S.; Nourian, A.; Shalizar-Jalali, A.; Najafi, G.; Nazarizadeh, A.; Koohestani, M.; Karimi, A. Selenium supplementation in the form of selenium nanoparticles and selenite sodium improves mature male mice reproductive performances. Iran. J. Basic Med Sci. 2018, 21, 577.
- Martins, S.M.M.K.; Afonso, E.R.; Parazzi, L.J.; Andrade, A.F.C.D.; Leal, D.F.; Gameiro, A.H.; Moretti, A.D.S.A.; Arruda, R.P.D. Organic selenium supplementation is cost-effective for increasing the number of seminal doses produced by sexually mature boars. Rev. Bras. Zootec. 2018, 47.
- Liu, L.; He, Y.; Xiao, Z.; Tao, W.; Zhu, J.; Wang, B.; Liu, Z.; Wang, M. Effects of selenium nanoparticles on reproductive performance of male Sprague-Dawley rats at supranutritional and nonlethal levels. Biol. Trace Elem. Res. 2017, 180, 81–89.
- Zhou, J.-C.; Zheng, S.; Mo, J.; Liang, X.; Xu, Y.; Zhang, H.; Gong, C.; Liu, X.-L.; Lei, X.G. Dietary selenium deficiency or excess reduces sperm quality and testicular mRNA abundance of nuclear glutathione peroxidase 4 in rats. J. Nutr. 2017, 147, 1947–1953.
- Ibrahim, H.A.; Zhu, Y.; Wu, C.; Lu, C.; Ezekwe, M.O.; Liao, S.F.; Haung, K. Selenium-enriched probiotics improves murine male fertility compromised by high fat diet. Biol. Trace Elem. Res. 2012, 147, 251–260.
- Shalini, S.; Bansal, M. Dietary selenium deficiency as well as excess supplementation induces multiple defects in mouse epididymal spermatozoa: Understanding the role of selenium in male fertility. Int. J. Androl. 2008, 31, 438–449.
- Sánchez-Gutiérrez, M.; García-Montalvo, E.; Izquierdo-Vega, J.; Del Razo, L. Effect of dietary selenium deficiency on the in vitro fertilizing ability of mice spermatozoa. Cell Biol. Toxicol. 2008, 24, 321–329.
- Mohammadi, S.; Movahedin, M.; Mowla, S.J. Up-regulation of CatSper genes family by selenium. Reprod. Biol. Endocrinol. 2009, 7, 126.
- Abdel-Wareth, A.; Ahmed, A.; Hassan, H.; El-Sadek, M.A.; Ghazalah, A.; Lohakare, J. Nutritional impact of nano-selenium, garlic oil, and their combination on growth and reproductive performance of male Californian rabbits. Anim. Feed Sci. Technol. 2019, 249, 37–45.
- Baiomy, A.; Mohamed, A.; Mottelib, A. Effect of dietary selenium and vitamin E supplementation on productive and reproductive performance in rams. In Proceedings of the 14th International Congress of the International Society for Animal Hygiene (ISAH), Vechta, Germany, 9–23 July 2009; pp. 43–46.
- Jacyno, E.; Kawecka, M.; Kamyczek, M. Influence of inorganic Se+ vitamin E and organic Se+ vitamin E on reproductive performance of young boars. Agric. Food Sci. 2002, 11, 175–184.
- Hajalshaikh, A.; Al-Hassan, M.; Mohamed, H. The influence of injectable sodium selenite on semen characteristics and testosterone concentration in Aardi goats. Indian J. Anim. Res. 2015, 49, 793–797.
- Martins, S.M.M.K.; De Andrade, A.; Zaffalon, F.G.; Parazzi, L.J.; Bressan, F.F.; Pugine, S.M.P.; Melo, M.P.d.; Chiaratti, M.; Marino, C.T.; Afonso, E.R. Organic selenium increases PHGPx, but does not affect quality sperm in raw boar semen. Livest. Sci. 2014, 164, 175–178.
- Petrujkić, B.; Šefer, D.; Jovanović, I.; Jovičin, M.; Janković, S.; Jakovljević, G.; Beier, R.; Anderson, R. Effects of commercial selenium products on glutathione peroxidase activity and semen quality in stud boars. Anim. Feed Sci. Technol. 2014, 197, 194–205.
- El-Sharawy, M.; Eid, E.; Darwish, S.; Abdel-Razek, I.; Islam, M.R.; Kubota, K.; Yamauchi, N.; El-Shamaa, I. Effect of organic and inorganic selenium supplementation on semen quality and blood enzymes in buffalo bulls. Anim. Sci. J. 2017, 88, 999–1005.
- Lukusa, K.; Lehloenya, K. Selenium supplementation improves testicular characteristics and semen quality of Saanen bucks. Small Rumin. Res. 2017, 151, 52–58.
- Anchordoquy, J.P.; Anchordoquy, J.M.; Lizarraga, R.M.; Nikoloff, N.; Pascua, A.M.; Furnus, C.C. The importance of trace minerals copper, manganese, selenium and zinc in bovine sperm–zona pellucida binding. Zygote 2019, 1–8.
- Roychoudhury, S.; Sharma, R.; Sikka, S.; Agarwal, A. Diagnostic application of total antioxidant capacity in seminal plasma to assess oxidative stress in male factor infertility. J. Assist. Reprod. Genet. 2016, 33, 627–635.
- Gharagozloo, P.; Gutiérrez-Adán, A.; Champroux, A.; Noblanc, A.; Kocer, A.; Calle, A.; Pérez-Cerezales, S.; Pericuesta, E.; Polhemus, A.; Moazamian, A. A novel antioxidant formulation designed to treat male infertility associated with oxidative stress: Promising preclinical evidence from animal models. Hum. Reprod. 2016, 31, 252–262.
- Anel-López, L.; Ortega-Ferrusola, C.; Martínez-Rodríguez, C.; Álvarez, M.; Borragán, S.; Chamorro, C.; Peña, F.; Anel, L.; de Paz, P. Analysis of seminal plasma from brown bear (Ursus arctos) during the breeding season: Its relationship with testosterone levels. PLoS ONE 2017, 12, e0181776.
- Bertelsmann, H.; Keppler, S.; Höltershinken, M.; Bollwein, H.; Behne, D.; Alber, D.; Bukalis, G.; Kyriakopoulos, A.; Sieme, H. Selenium in blood, semen, seminal plasma and spermatozoa of stallions and its relationship to sperm quality. Reprod. Fertil. Dev. 2010, 22, 886–891.
- Hawkes, W.C.; Turek, P.J. Effects of dietary selenium on sperm motility in healthy men. J. Androl. 2001, 22, 764–772.
- Domosławska, A.; Zdunczyk, S.; Franczyk, M.; Kankofer, M.; Janowski, T. Selenium and vitamin E supplementation enhances the antioxidant status of spermatozoa and improves semen quality in male dogs with lowered fertility. Andrologia 2018, e13023.
- Domosławska, A.; Zduńczyk, S.; Niżański, W.; Jurczak, A.; Janowski, T. Effect of selenium and vitamin E supplementation on semen quality in dogs with lowered fertility. Bull. Vet. Inst. Pulawy 2015, 59, 85–90.
- Butt, M.A.; Shahid, M.Q.; Bhatti, J.A.; Khalique, A. Effect of Dietary Vitamin E and Selenium Supplementation on Physiological Responses and Reproductive Performance in Holstein Friesian Bulls during Humid Hot Summer. Pak. Vet. J. 2019, 1–5.
- Ghorbani, A.; Moeini, M.M.; Souri, M.; Hajarian, H. Influences of dietary selenium, zinc and their combination on semen characteristics and testosterone concentration in mature rams during breeding season. J. Appl. Anim. Res. 2018, 46, 813–819.
- Kirchhoff, K.; Failing, K.; Goericke-Pesch, S. Effect of dietary vitamin E and selenium supplementation on semen quality in Cairn Terriers with normospermia. Reprod. Domest. Anim. 2017, 52, 945–952.
- Domosławska, A.; Zduńczyk, S.; Janowski, T. Improvement of sperm motility within one month under selenium and vitamin E supplementation in four infertile dogs with low selenium status. J. Vet. Res. 2019.
- Alonge, S.; Melandri, M.; Leoci, R.; Lacalandra, G.M.; Caira, M.; Aiudi, G.G. The Effect of Dietary Supplementation of Vitamin E, Selenium, Zinc, Folic Acid, and N-3 Polyunsaturated Fatty Acids on Sperm Motility and Membrane Properties in Dogs. Animals 2019, 9, 34.
- Ran, M.-X.; Li, Y.; Zhang, Y.; Liang, K.; Ren, Y.-N.; Zhang, M.; Zhou, G.-B.; Zhou, Y.-M.; Wu, K.; Wang, C.-D. Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm. Int. J. Mol. Sci. 2018, 19, 3066.
- Dai, D.-H.; Qazi, I.H.; Ran, M.-X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.-B.; Angel, C.; Zeng, C.-J. Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing. Int. J. Mol. Sci. 2019, 20, 802.
- Jamali, N.U.; Kaka, A.; Khatri, P.; Malhi, M.; Naeem, M.; Memon, A.A.; Kaleri, R.R.; Janyaro, H.; Kalhoro, D.H. Effect of in vitro Selenium Addition to the Semen Extender on the Spermatozoa Characteristics before and after Freezing in Kundhi Buffalo Bull and in vivo Fertility Rate. Pak. J. Zool. 2019, 51, 317–323.
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.; Hassan, M.A. Impact of selenium nano-particles in semen extender on bull sperm quality after cryopreservation. Theriogenology 2019, 126, 121–127.
More
Information
Subjects:
Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.5K
Revisions:
2 times
(View History)
Update Date:
15 Sep 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No