1000/1000
Hot
Most Recent
 +1 point
+1 point
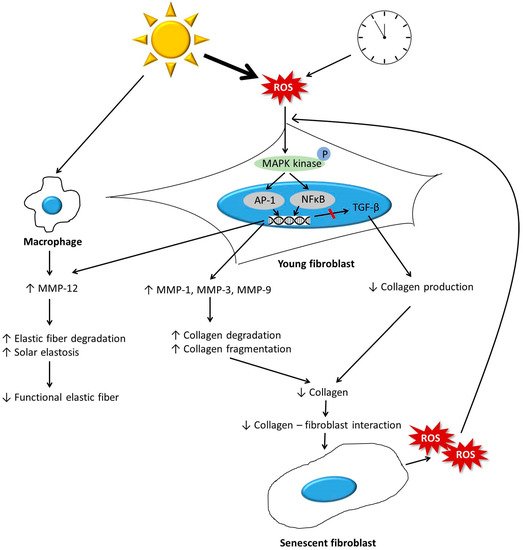
The dermis is primarily composed of the extracellular matrix (ECM) and fibroblasts. During the aging process, the dermis undergoes significant changes. Collagen, which is a major component of ECM, becomes fragmented and coarsely distributed, and its total amount decreases. This is mainly due to increased activity of matrix metalloproteinases, and impaired transforming growth factor-β signaling induced by reactive oxygen species generated during aging. The reduction in the amount of collagen hinders the mechanical interaction between fibroblasts and the ECM, and consequently leads to the deterioration of fibroblast function and further decrease in the amount of dermal collagen. Other ECM components, including elastic fibers, glycosaminglycans (GAGs), and proteoglycans (PGs), also change during aging, ultimately leading to a reduction in the amount of functional components. Elastic fibers decrease in intrinsically aged skin, but accumulate abnormally in photoaged skin. A reduction in the levels of functional dermal components results in the emergence of clinical aging features, such as wrinkles and reduced elasticity. Various antiaging approaches, including topicals, energy-based procedures, and dermal fillers, can restore the molecular features of dermal aging with clinical efficacy.
|
Photoaging |
Components |
Intrinsic Aging |
|---|---|---|
|
Decreased and fragmented |
Collagen |
Decreased and fragmented |
|
Abnormally accumulated (SE) |
Elastic fiber |
Decreased |
|
Increased in SE region |
Hyaluronic acid |
Not changed |
|
Increased |
Total sulfated GAGs |
Decreased |
|
Increased in SE region |
Versican |
Not changed? |
|
Not changed |
Biglycan |
Decreased |
|
Decreased in SE region |
Decorin |
Not changed? |
GAG = glycosaminoglycan, SE = solar elastosis.
|
Modalities |
Mechanisms of Action |
Clinical Efficacies |
|---|---|---|
|
Topicals |
||
|
Retinoid acid (RA) |
Acts through RARs and RXRs [47] Increases type I, III, and VII collagens [48] Reorganizes elastic fiber [50] Normalizes GAG deposition [51] |
Application of 0.05% RA for 6 months improved fine and coarse wrinkles, roughness, and skin laxity [52]. Application of 0.025% RA for 3 months improved rough and fine wrinkles, skin firmness, and roughness (Ho ET) |
|
Ascorbic acid |
Reduces ROS [53] Acts as a cofactor in the biosynthesis of procollagen and elastin [54] Induces collagen synthesis in human skin fibroblasts and increase dermal thickness [55][56] |
Application of 5% ascorbic acid for 6 months led to a clinical improvement of the photodamaged skin [57]. |
|
Glycolic acid |
Stimulates the production of GAGs and collagen in the dermis [58] Improves histologic quality of elastic fibers [58] |
Application of 25% glycolic acid for 6 months increased skin thickness [58]. |
|
Peptides |
Regulate fibroblasts and control the production of ECM [59][60]. |
Application of Pal-KTTKS for 3 months reduced wrinkles [61]. Application of copper–GHK reduced the depth and length of wrinkles and made skin smoother [62]. |
|
Energy-based devices |
||
|
Fractional lasers (FL) |
Heat the dermis and stimulate matrix remodeling by deeply penetrating columns of laser energy [63] Induce biosynthesis of type I and III protocollagens [63][64] |
Two or three treatments with CO2 fractional laser improved skin texture, laxity, and overall cosmetic outcome [65]. Two treatments with 2790 nm Er:YSGG laser improved wrinkle and skin texture [66]. Three treatments of fractional 1550 nm erbium-doped fiber laser improved wrinkles [67]. |
|
Ablative FL |
||
|
Nonablative FL |
||
|
Radiofrequency (RF) |
Causes direct collagen contraction and immediate skin tightening [68][69] Reorganizes collagen bundles [70] Induces increase in types I and III collagens [71] Improves the quality of elastic fibers and solar elastosis [71] |
Three treatments with fractional bipolar RF improved wrinkles and skin texture [72]. Six treatments with monopolar RF improved laxity, texture, and wrinkles [73]. |
|
High-intensity focused ultrasound (HIFU) |
Creates precision microwounds in the dermis [74] Induces the higher level of neocollagenesis and neoelastogenesis in the deep reticular dermis [75] |
Single treatment with HIFU improved skin laxity of lower face and neck [76]. Single treatment with HIFU improved skin laxity of face and upper neck [77]. |
|
Fillers |
||
|
Restore the contractile properties and elongation of aged fibroblasts [78][79] Induce type I collagen synthesis [78] |
Further investigation is needed. |
ECM = extracellular matrix, ER:YSGG = erbium:yttrium scandium gallium garnet, GAG = glycosaminoglycan, GHK = glycyl-L-histidyl-L-lysyl, MMP = matrix metalloproteinase, pal-KTTKS = palmitoyl pentapeptide palmitoyl-lysine-threonine-threonine-lysine-serine, RAR = retinoic acid receptor, RXR = retinoid X receptors.





