 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Katarzyna Bogunia-Kubik | + 4508 word(s) | 4508 | 2021-06-16 07:43:58 | | | |
| 2 | Nora Tang | + 940 word(s) | 5448 | 2021-07-02 08:30:04 | | |
Video Upload Options
The functionality of NK cells is preserved by the set of activating and inhibitory receptors. Under normal conditions, the appropriate level of major histocompatibility complex (MHC) class I molecules is supervised by inhibitory receptors, which keep NK cells silenced. The “missing self” mechanism considers reduced levels of MHC I (Ia and Ib) molecules when NK cells become activated.
1. Introduction
Natural killer (NK) cells are the first line of defense of the innate immune system, essential for protection against viral infections and detection of malignant cells [1]. They are responsible for foreign cell recognition and play a key role in immune surveillance as well as in antimicrobial, antiviral and antitumor responses [2]. NK cells are large granular lymphocytes present in lymphoid organs and non-lymphoid peripheral tissues [3]. Their main task is to destroy transformed or infected cells without affecting normal host cells.
The functionality of NK cells is preserved by the set of activating and inhibitory receptors. I molecules is supervised by inhibitory receptors, which keep NK cells silenced. The “missing self” mechanism considers reduced levels of MHC I (Ia and Ib) molecules when NK cells become activated [4]. These activated NK cells produce cytokines and gain cytotoxic properties that make them fully functional [5][6].
Their activity was first observed in peripheral blood mononuclear cells (PBMCs) In humans, there are two subsets of NK cells—CD56bright and CD56dim—depending on expression of a surface antigen CD56 (neural cell adhesion molecule) [7]. CD56dim cells express killer cell immunoglobulin-like receptors and have a strong cytolytic activity, whereas CD56bright cells secrete large quantities of cytokines [7][8][9][10]. The CD56dim cells make up to 90% of all blood NK cells, which makes them the major subset of NK cells, while CD56bright cells are present in lymphoid tissue [8].
The functionality of NK cells is preserved by the set of activating and inhibitory receptors localized on their surface, which are responsible for interacting with ligands expressed on transformed, infected or stressed cells. In humans, normal and healthy cells express human leukocyte antigen (HLA) class I molecules, inducing activity of inhibiting receptors. On the other hand, transformed or infected cells are characterized by decreased expression of the HLA molecules, which affects NK cell activation through activating receptors and their potential to kill the target cells [7][11][12].
NK cells have no T cell receptors (TCRs) or Natural Killer T (NKT) receptors [13]. Killer cell immunoglobulin-like receptors (KIRs) are one of the major NK cell receptor groups in humans, consisting of both inhibitory and activating receptors [11]. As they can potentially suppress T cell responses and proliferation, they are thought to have a role in allograft tolerance during transplantation and cancer [14][15]. They are responsible for recognizing tumor cells and inducing cytotoxic effects.
withC-type lectin-like extracellular domain, transmembrane segment and cytoplasmic tail. They are inhibiting receptors containing two ITIMs in their cytoplasmic tails. With an exception of NKG2D and NKG2F, all NKG2 receptors form heterodimers with invariant chains of CD94 by disulfide bonds [16]. They are still poorly understood, but studies suggest that NKG2H is expressed on effector cells more likely [17].
In humans, it is expressed on almost all NK cells and some subsets of T cells such as NKT cells, CD8+ TCR-αβ T, CD4+ T, TCR-γδ T cells, while in mice it is expressed on NK cells and activated macrophages [18][19]. Its ligands (NKG2DLs) are commonly overexpressed in transformed and infected cells or in cells with damaged DNA [20][21]. NKG2D triggers NK cell cytotoxicity and cytokine secretion [22]. Human NKG2D can associate only to DAP10 protein, while in mice, there are two isoforms of NKG2D, long and short; therefore, it can bind with two types of adaptor molecules—
I molecules [18]. NKG2D ligands are mostly expressed on the cells exposed to a stress factor, such as heat shock or chemical stimuli, while they are absent or expressed at a very low level in normal cells. Expression of NKG2DLs is related to sensitivity of target cells to NK cells cytotoxicity [23]. One of the hypotheses for NKG2DL diversity is that each ligand is associated with different stress signal types (various oncogenes, infections, tumor suppressors), while another suggests that ligand diversity reduces the chances of escaping immunosurveillance by transformed cells [24].
Predominantly, NKG2DLs are not present in normal human or murine cells; however, human MICA and ULBP3 molecules are expressed on bone marrows stem cells [25]. This discovery was the very first evidence of the stress cell detection by the NKG2D receptors. Furthermore, MICA and MICB are often expressed in epithelial tumors (e.g., lung, breast, kidney, prostate, ovarian and colon cancers), and they can act like stress-induced antigens in intestinal epithelium cells. The receptor’s involvement affects NK cell response and TCR-dependent T cell activation [26].
2. Variability of the NKG2D Gene—Its Functional Effect and Clinical Associations
The gene that encodes NKG2D protein is known asNKG2DorKLRK1(Killer Cell Lectin-Like Receptor K1). TheKLRK1gene is located on the human 12p12-p13 chromosome and belongs to the NK gene complex (NKC), which is a 270 kb size cluster of 19 genes encodingC-type lectins NKG2 and CD94 [27].KLRK1consists of eight exons and seven introns. A review of published reports demonstrates that polymorphisms in theKLRK1gene influence the natural cytotoxic activity, predisposing to infectious diseases, cancers, autoimmune disorders, pregnancy miscarriages or transplant-related mortality (Table 1).
| SNP ID | Localization [58] | Main Findings/Associations | Ref. |
|---|---|---|---|
| rs1049174 (G > C) |
KLRK1 exon 8 (3’UTR) | G allele associated with reduced risk of colorectal carcinoma (colon, rectal, both) in Japanese patients | [57] |
| G allele associated with reduced risk of aerodigestive cancer (head, neck, esophageal) among Japanese never smokers and never drinkers |
[59] | ||
| CC genotype associated with higher breast cancer risk in Iranian women | [60] | ||
| GG and GC genotypes associated with increased risk of gastric cancer in Chinese patients | [61] | ||
| GG homozygosity in chronic myeloid leukemia Japanese patients associated with better response to dasatinib treatment, compared to other CML patients’ genotypes; G allele affected the incidence of skin rash after Das treatment | [62] | ||
| CC homozygosity in Caucasians protected against spondyloarthritis | [63] | ||
| CC polymorphism associated with inefficient anti-TNF therapy in Polish rheumatoid arthritis patients; presence of C allele correlated with worse EULAR response |
[64] | ||
| G allele associated with rheumatoid arthritis lower disease activity scores among Greek patients | [65] | ||
| C allele associated with increased HPV+ cervical, anal and vaginal cancers risk in Vietnamese patients |
[66] | ||
| CC homozygosity associated with higher risk of EBV-infected nasopharyngeal carcinoma in Vietnamese patients |
[67] | ||
| GG genotype in chronic hepatitis C Iranian patients demonstrated higher response to Peginterferon Alfa-2a/Ribavirin therapy against HCV infection | [68] | ||
| No association between Caucasian alleles and CMV infection in the first year after kidney transplantation |
[69] | ||
| Donor GG genotype associated with improved overall survival and transplant related mortality in Japanese patients with standard risk disease after HLA-fully matched unrelated BM transplantation; no impact on disease relapse or development of aGvHD, cGvHD | [70] | ||
| C allele associated with decreased risk of recurrent miscarriages among Iranian women | [71] | ||
| rs2255336 (A, C > G, T) |
KLRK1 exon 4 (region encoding transmembrane part of receptor) |
A allele in Caucasian patients associated with lower incidence of systemic lupus erythematosus | [72] |
| GG genotype associated with systemic lupus erythematosus risk in Caucasian patients; reduced stimulatory effect of CD4+ NKG2D+ T cells carrying the GG variant | [73] | ||
| AA genotype associated with susceptibility to rheumatoid arthritis in Korean patients | [74] | ||
| GG genotype associated with inefficient anti-TNF therapy in rheumatoid arthritis Caucasian patients | [64] | ||
| TT genotype in Taiwanese chronic hepatitis B patients associated with a decreased risk of delayed spontaneous HBeAg seroconversion | [75] | ||
| Higher A allele frequency in Japanese congenital CMV infection cases symptomatic at birth than in asymptomatic cases | [76] | ||
| AA and GA genotypes associated with reduced HPV-related cancer risk; protective effect of A allele against the progression to advanced stages of cancer | [77] | ||
| rs2617160 (A > T) |
Intergenic KLRC4-KLRK1 | TT genotype associated with increased risk of chronic hepatitis B in Chinese patients | [78] |
| A allele associated with lower frequency of retinopathy—vascular complication in sickle cell disease in Sub-Saharan African and French West Indian patients | [79] | ||
| rs2246809 (A > G) |
Intergenic KLRC4-KLRK1 | A allele frequency higher in South Indian rheumatoid arthritis patients with deformities—marker of severe disease | [80] |
| A allele associated with lower frequency of retinopathy—vascular complication in sickle cell disease in Sub-Saharan African and French West Indian patients | [79] | ||
| rs2617169 (A > T) |
KLRC4 intron 3 | T allele frequency higher in South Indian rheumatoid arthritis patients with deformities—marker of severe disease | [80] |
| A allele associated with lower frequency of retinopathy—vascular complication in sickle cell disease in Sub-Saharan African and French West Indian patients | [79] | ||
| rs2617170 (T > C) | KLRC4 exon 3 | TT homozygosity associated with lower risk of recurrent miscarriage among North African women | [81] |
| rs7397310 (T > C) |
3’ flanking region of KLRK1 | No association between alleles frequency and cholangiocarcinoma risk in Caucasian individuals | [82] |
| rs2900420 (G > A) | 3’ flanking region of KLRK1 | A allele associated with better survival time in advanced-stage non-small cell lung cancer Non-Hispanic Whites patients treated with first-line chemotherapy |
[83] |
| rs10772271 (G > A) |
3’ flanking region of KLRK1 | A allele associated with decreased NKG2D expression on NK cells in Finnish infants, and increased viral bronchiolitis susceptibility |
[84] |
| rs11053781 (G > A) |
KLRK1 intron 3 | Higher G allele frequency in PSC Scandinavian patients with cholangiocarcinoma | [85] |
| rs7972757 (A > G) | Intergenic KLRC4-KLRK1 | Higher A allele frequency in placental malaria affected African patients | [86] |
| rs728010 (G > A) | Intergenic KLRC4-KLRK1 | Higher GG genotype frequency in placental malaria affected African patients | [86] |
| rs12821887 (T > C) |
Intergenic KLRC4-KLRK1 | Higher TT genotype frequency in placental malaria affected African patients | [86] |
| rs2617167 (G > A) | Intergenic KLRC4-KLRK1 | Higher A allele frequency in PSC Scandinavian patients with cholangiocarcinoma | [85] |
| rs1154831 (A > C) |
Intergenic KLRC4-KLRK1 | CC genotype associated with lower disease activity scores in rheumatoid arthritis Greek patients | [65] |
| CCGGGCA > CCAGGCG (rs1049174, rs2255336, rs11053781, rs12819494, rs728010, rs2617165, rs2617167) | Haplotypes associated with increasing/decreasing cholangiocarcinoma risk in PSC Scandinavian patients |
[85] | |
| GAT (rs2255336, rs224809, rs2617169) | Haplotype associated with higher risk for cutaneous melanoma in Caucasian patients | [87] | |
| GCAGATCC (rs1049174, rs2255336, rs2617160, rs2246809, rs2617169, rs2617170, rs2617171, rs1983526) | Higher haplotype frequency in rheumatoid arthritis among South Indian patients | [80] | |
| CAT > GGA (rs1049174, rs2255336, rs2617160) | Haplotypes associated with higher recurrent miscarriage risk among North African women |
[81] | |
| CAA (rs1049174, rs2255336, rs2617160) | Haplotypes associated with higher recurrent miscarriage risk among North African women | [81] | |
| GTTG > ATTC (rs2246809, rs2617169, rs2617170, rs2617171) | Haplotypes associated with increasing/decreasing recurrent miscarriage risk among North African women | [81] | |
| (ATG, TTC) > (TTC, TTG) (rs1746123, rs10431294, rs1049174) | KLRK1 haplotypes associated with decreasing/increasing gastric cancer risk in Chinese patients | [61] | |
One of the earliest studies describingKLRK1polymorphisms was based on previous indications of the relationship between natural cytotoxic activity of peripheral blood lymphocytes and cancer susceptibility in the Saitama population [28]. Comparison of the allele frequencies in groups with high and low natural cytotoxic activity showed an association of eight SNPs with natural cytotoxic activity: two of them were located within theKLRK1gene (rs1049174 The examined alleles formed LNK1, HNK1 and LNK2, HNK2 haplotypes with strong linkage disequilibrium (LD), related to low and high natural cytotoxic activity phenotypes, respectively. /LNK1 haplotypes is associated with significantly increased NKG2D protein surface expression on NK cells and reduced overall cancer risk, compared to the LNK1/LNK1 haplotype [27][29].
To the best of our knowledge, among all of the genetic substitutions located within theKLRK1gene, two of them have been uncovered as functional polymorphisms: rs1049174 and rs2255336. transcribed from the rs1049174 G variant, resulting in higher NKG2D expression in vitro [30]. (A > G), is the only non-synonymous polymorphism located in the transmembrane region in exon 4, close to the binding site of the adapter protein DAP10 [31], and near the intron–exon junction region with potential to affect splicing [32]. The SNP does not affect the expression ofKLRK1gene [31]; nonetheless, proteins encoded by theKLRK1rs2255336 variants may differ in affinity to DAP10 binding, hence transmitting different signal strength inside activated NK cells upon bonding NKG2D ligands.
Reports summarized in Table 1 clearly showed a positive effect of increased cytotoxicity mediated by HNK genotypes on cancer immunosurveillance, virus eradication, and a protective effect against viral-induced cancer development. Some alleles can be used as markers of severe clinical presentation of the disease [33][34][35].KLRK1gene polymorphism may represent a potential biomarker for prediction of treatment outcome in patients with cancers, autoimmune diseases or virus induced diseases [36][32][37][38][39]. Thanks to these, novel importantKLRK1SNPs that do not originate from the HNK/LNK genotypes were detected recently [39][40][41].
3. Multiple Functions of the Activating NKG2D Receptor—Clinical Implications and Application for Treatment
The NKG2D receptor is a well-known molecule with varied functions and features (Figure 1). In the next paragraphs, its role in cancer immunosurveillance, autoimmune diseases, transplantation, infections together with the defense mechanisms of target cells to avoid the cytotoxic effect of NK cells as well as NKG2D-based treatment options will be presented and discussed.
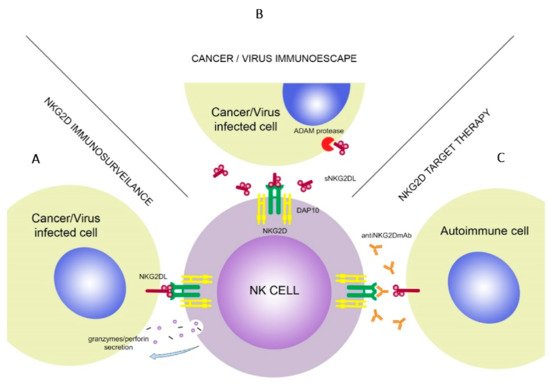
NK cells have a crucial role in tumor immunosurveillance through their ability to detect changes in MHC class I expression. During the transformation, expression of various surface molecules changes, i.e., downregulation of “self” MHC class I or upregulation of stress-induced molecules, which can be detected by NKG2D. Long term follow-up study showed that lack of cytotoxic activity of NK cells can be connected with increased risk of cancer occurrence [13][27][42].
One inconsistent finding indicated that GG and GC genotypes of rs1049174 are related to gastric cancer incidence [43], although it can be assumed that rs1049174 G allele has a protective effect on cancer occurrence in different populations. A study conducted on non-PSC Caucasian patients failed to reveal any correlation between NKG2D genetic variation and CC development risk [44]. The contribution of NK cells’ cytotoxic activity in the mechanisms of malignant transformation may be more significant in the inflamed biliary ducts than in non-inflammatory conditions. Higher risk of melanoma development was observed in individuals from south-eastern Spain, bearing the rs2255336 G, rs2617169 T, rs2246809 A, and haplotype, which refers to the low NK activity LNK2 haplotype, except of rs2246809 A, a major variant of the HNK2 haplotype [45].
The NKG2D receptor could be used as an independent prognostic indicator for gastric cancer. level is significantly associated with death risk reduction in patients with gastric cancer. Treatment with chimeric NKG2D-expressing T cells is a potential immunotherapy for gastric cancers with peritoneal metastasis [46][47]. Paradoxically, in certain advanced cancers, tumor progression is connected with high levels of NKG2D despite overexpression of NKG2DLs on the surface of cancer cells [48].
NKG2D binding with its ligands on the cell surface may start several signaling pathways such as phosphatidylinositol 3-hydroxy kinase (PI3K), phospholipase C Gamma 2 (PLCG2), c-Jun-NH(2)-terminal kinase (JNK) [49]. This results in increased antitumor properties of the NK cells due to enhancing Antibody-Dependent Cellular Cytotoxicity (ADCC) effects, secreting cytokines and initiating apoptosis. However, on the other hand, studies on mice developing HCC showed that NKG2D-deficient mice had an increased survival rate compared with the wild type [50]. This suggests that NKG2D receptors may promote HCC progression, which should be considered in HCC immunotherapy.
NKG2DL expression can be modified by advanced cancer cells to develop various mechanisms helping them to escape the immune system. Overexpression of strongly limited NKG2DLs was observed in transformed cells, which makes them more prone to cytotoxic activity of NK cells, although high levels of ULBP2 and ULBP4 expression are often connected with bad prognosis in ovarian cancer [51]. Tissue isolated from patients suffering from different types of tumors (e.g., breast, lung, prostate, kidney), melanoma or leukemia showed increased MICA expression [52]. The MICA level is also increased in various epithelial cancers such as breast [53], intestinal or stomach cancers [54]; however, in the meta-analysis performed by Zhao et al., a high cell-surface MICA and MICAB expression was found to be associated with increased survival in cancers of the digestive system [26].
Cancer cells have the ability to avoid detection by the immune system. One of the immunoescaping mechanisms is shedding soluble forms of NKG2D ligands, which is often associated with poor outcomes in different types of cancer. The level of soluble forms of MICA and MICB in serum are associated with the state of disease and survival rates in oral squamous cell carcinoma patients [55]. Soluble ULBP2 molecules are related to poor prognosis in patients with lung cancer [56], melanoma [57] and chronic lymphocytic leukemia (CLL) [58].
It has been shown that ADAM10 and ADAM17 are upregulated in breast cancer. Inhibition of ADAMs may decrease the cleavage of CD16 receptors, which are expressed in CD56dim cells. Studies in vitro proved that inhibition of ADAM17 enhances the ADCC activity of NK cells against breast cancer cell lines treated with trastuzumab. Activation of ADAM17 results in CD16A cleavage, which decreases attachment of NK cells to their targets.
The NKG2D-NKG2DL signaling pathway blockade results in activation of the T cells reacting to cancer cells. As studies showed, absence of NKG2D signaling promotes differentiation of aberrant memory T cells. Temporary blockade of NKG2D signaling in the effector phase resulted in development of altered defective memory CD8+ T cells, which means that uninterrupted NKG2D signaling in the effector phase promotes development of fully functional memory CD8+ T cells. This is a new feature of the NKG2D receptor [59].
CAR-T cell therapy has become a revolutionary strategy of cancer treatment. T cells isolated from patients can be modified to express synthetic, chimeric antigen receptor (CAR) binding to cancer antigen which will result in cancer cell death. so it would be difficult to use this treatment in patients with rapid development of disease. Due to these limitations, modified therapy—the CAR-NK cell therapy—is now being considered as an alternative treatment (Table 3).
As studies showed, NK cells are able to trace and kill lymphohematopoietic host cells, which take part in the process of donor cell rejection [60]. In the first clinical study [61], autologous T cells were isolated from the cancer patients’ blood and electroporated to suppress TCR receptors and reduce apoptosis. The first study on CAR-NK-92 cell modified therapy against prostate cancer resulted in cells expressing prostate-specific membrane antigen (PSMA), which made an effective prostate cancer-specific therapeutic tool [62]. Thus, genetic modification of these cells with CARs may be a good alternative to standard CAR-T cell therapy for better efficacy and specificity.
This supports the theory that downregulation of the NKG2D expression reduces NK cell cytotoxic activity in leukemia patients [63]. This may relate to the tumor production of immunosuppressive cytokines and other inhibitory factors, especially in the advanced stages of hematological malignancy and multiple myeloma [64]. In multiple myeloma bone marrow mesenchymal stromal cells (BMMSCs) are the microenvironment mediators for immune resistance against NK cells. Studies in vitro, using the KHYG-1 NK cell line, showed that overcoming the effect of BMMSCs may be possible through inhibition of antiapoptotic molecules or increasing the efficacy of killer cells [65].
This study confirms that the NKG2D receptor is able to activate NK cells by inducing their cytotoxicity effect to specific target cell lines. Interestingly, Sandoval-Borrego et al. observed that patients suffering from AML M3 showed decreased NKG2D expression [66]. Earlier studies showed that the level of NKG2D expression is decreased in patients suffering from different types of leukemia, which is caused by shedding of a soluble form of NKG2D ligands [67]. Chemotherapy resistant cells in AML patients, leukemic stem cells (LSCs), have the ability to induce leukemia in immunocompromised mice [68].
Overexpression of NKG2D ligands is reported in various types of malignant cells but the level of their expression is different depending on cancer type. It has been reported that cancer cells in solid tumors have more than one type of ligand which makes them more susceptible to the immune system while blood cancer cells can avoid its detection. One of the strategies of AML treatment is increasing the level of NKG2DLs expression on cells’ surface which may help in detection of cancer cells and prevent their immunoescape. It has been shown that the treatment of cancer cells with demethylating factors increases the level of NKG2DLs expression.
Prolonged treatment of anti-NKG2D mAb was necessary to avoid rejection in murine cardiac transplant model CD28−/−. It has been shown that anti-NKG2D mAb treatment was highly effective in preventing rejection of cardiac allografts that are independent from CD28 [69]. In cardiac xenografts (mouse-rat) models an upregulation of soluble form of MICA (sMICA) in acute graft rejection occurring 2h after transplantation was observed indicating that MICA expression may be related to a high risk of acute graft rejection [70]. Impairment of NK cells results in graft rejection in cardiac allograft models [71].
Endogenous RAE-1e desensitizes NK cells and decreases antitumor NK cytotoxicity and its rejection by interaction with NKG2D ligands in murine models [72]. Studies on mice models of allogeneic HSCT show that NKG2D expression by CD8+ T cells mediates graft versus host disease (GvHD) and graft versus tumor (GvT) effects [73]. It has been reported that short NKG2D blockade with anti-NKG2D mAb reduces GvHD while keeping the GvT effect in 75% of studied mice. Treatment with anti-NKG2D mAb prevents transplantation rejection in the F1 generation and enhances engraftment.
Upregulation of NKG2C and NKG2D receptors is observed after HSCT [74]. High expression of these activating receptors may overcome the inhibiting effect caused by NKG2A. Upregulation of NKG2C and NKG2D during engraftment may also be beneficial for infection control in immunosuppressed patients. It has been observed that activating receptors are expressed at a higher level in transplanted donor cells in HSCT. Moreover, NK cells are known to initiate allogeneic cells rejection, like BM allografts [75].
NK cell inhibition is a major factor protecting normal tissues from the autoimmune effects. Donor NK cells may lyse allo-reactive T cells (which are associated with occurrence of acute GvHD). This may help to predict a risk for acute GvHD depending on the presence of NK cells cytotoxic to allo-reactive T cells. Studies showed that ligand expression is higher after chemoradiotherapy treatment, which may suggest that NKG2DL’s induction increases immunogenicity of glioma cells.
The donor and recipientKLRK1gene variations, together with HLA disparity, affect the clinical outcomes of patients undergoing allogeneic myeloablative BM transplantations. Patients with standard risk disease receiving transplants from unrelated, HLA-matched donors with the rs1049174 G gene variant had a significantly decreased risk of transplant-related mortality and a better survival rate [38]. This polymorphism, as well as rs2255336, did not show any influence on the development of either acute or chronic GvHD, or disease relapse in BM transplant patients [38][76].
Ligand recognition by the NKG2D is also crucial in viral infections. Viruses have a variety of defence mechanisms to evade the cytotoxic effect of NK cells mediated by NKG2D receptors. [77], by protein binding and retaining (such as ULP16 binding to ULBP1, ULBP2, ULBP6 and MIC [78][79], or by targeting NKG2DLs mRNA for lysosomal degradation, such as US18 and US20 downregulating MICA expression [80]. Moreover, the viral protein UL16 interferes with the cell surface expression of ULBP1 and blocks interaction with the NKG2D receptor [81][82].
Another mechanism of immune evasion is releasing the soluble form of NKG2DLs, which decreases the NKG2DL level on the cell surface [83]. Decreased expression of NKG2D enables viral immunoescape, which leads to increased pro-inflammatory cytokine expression and enhanced lung pathology. In respiratory syncytial virus (RSV), lower activity of NKG2D is caused by a high expression of sMICA [84]. Infection with Epstein–Barr virus (EBV) leads to HLA class I downregulation and modulation of NKG2D activity
A higher frequency of rs1049174 CC genotype and C allele in Vietnamese patients with Epstein–Barr virus-induced nasopharyngeal carcinoma High levels of TGF-β1 were detected in serum of patients with EBV-induced cancers [85]. CC genotype may affect the progression of viral infection and tumor formation.
Increased cytotoxicity mediated by HNK genotypes has a protective effect on virus eradication and viral-induced cancer development. It was found that rs2617160 TT homozygosity (associated with low NK activity) was more common in Han Chinese patients with chronic HBV than in those who cleared HBV spontaneously [86]. The authors presumed that this variation located within the non-codingKLRK1-KLRC4readthrough region may act as a tag SNP in linkage disequilibrium with functional transcription regulators responsible for the immune response to HBV infection. The rs2255336 TT genotype may increase the progression of HBeAg seroconversion by alternating NK cell cytotoxic properties in Taiwanese chronic hepatic B patients [87].
Induction of TGF-β1 in host cells by E6 and E7 oncoproteins of HPV is a proposed mechanism by which HPV-induced cancer cells escape from immune responses mediated by NKG2D receptors [42]. The impact of the TGF-β1 downregulation signal might be modulated by different affinity of rs1049174 C and G variants within 3′UTR NKG2D mRNA to miR-1245 binding. A study on Polish women showed a protective function of rs2255336 AA and GA genotype against HPV-related cervical cancer [88].
NKG2D gene variability can be regarded as a marker for virus treatment outcome. In Iranian patients suffering from chronic HCV infection, an association with treatment outcome and NKG2D polymorphism was observed [37]. The rs1049174 G allele or GG genotype were predictive factors of sustained virologic response after Peginterferon Alfa-2a/Ribavirin treatment.
It is worthy of note that viral infections are not the only ones capable of NK cell activation, as bacterial infections may also trigger the NK cells. Endocellular bacteria (i.e., mycobacteria) have the ability to induce the expression of NCR or NKG2D ligands, transforming the infected cell into a detectable target for NK cells [89].
People with weakened immune systems suffer from more dangerous conditions compared with healthy ones. This mechanism may lead to suppress NK cells functions, as NK+T cells express ACE2 [90]. Limited research on the NK cell derangement during SARS-CoV-2 infection demonstrate that patients suffering from COVID-19 have an increased amount of NK and CD8+T cells with an exhausted phenotype and high expression of the inhibitory receptor, NKG2A [91]. In this case IVIG may promote NK cytolytic functions, and reduce release of IL-6 by activated inflammatory cells [92].
Target cell recognition by the NK cells may be insufficient virus eradication. This idea uses NK cells modified to express both ACE2 and NKG2D receptors, which are able to target viral S protein and NKG2DL’s on the infected cells’ surface, respectively. Target cells are attacked with the synergistic effect of IL15 superagonist and cytokine release syndrome prevention through GM-CSF neutralizing scFv. SARS CoV-2 is much more dangerous to patients with a weakened immune systems.
NKG2D receptor can be regarded as a fragile link between infection and tumor eradication and progression of autoimmune disorders. NKG2D receptors have a different role in the autoimmune disorders than in cancer progression and viral infections. Hence, NKG2D polymorphisms associated with high NK cytotoxic activity act as risk factors for autoimmune disorders. Another study has shown that not individual SNPs, but particular NKG2D haplotype composed of rs1049174 and rs2255336 variants, can be associated with RA risk [35].
The previous study by Bogunia-Kubik research group determined that NKG2D polymorphisms may act as pharmacogenomic biomarkers of responsiveness to RA therapy [32]. The antitumor necrosis factor (anti-TNF)F treatment failure was found more frequently in Polish patients carrying rs1049174 In both studied polymorphisms, a lack of response to anti-TNF treatment in RA correlated with loss of NK cell activity. The presence of heterozygous genotypes of both SNPs was positively correlated with a better response to anti-TNF treatment, which may imply that balanced NKG2D activity is a relevant feature of successful anti-TNF therapy.
It is worth considering autoimmune diseases cases with viral etiology, such as systemic lupus erythematosus (SLE) induced by chronic viral infections such as EBV [31], in which HNK genotypes were associated with decreased disease susceptibility. The low NK activity rs2255336 GG variants were associated with SLE susceptibility in German and Polish cohorts [93][31]. In the study of Kabalak et al., peripheral blood lymphocytes with the NKG2D 72Ala protein were characterized by lower proliferation than those with the 72Thr variant, when stimulated by CD3+ and NKG2D antibodies.
Experiments on mice showed that blockade of the receptor’s interaction with its ligands may be an efficient form of type I diabetes treatment. This suggests new possibilities of the NKG2D-NKG2DL path blockade usage not only in treatment of cancers or infections but also in autoimmune diseases [94].
Regulation of NKG2D may be beneficial in Crohn’s disease (CD) treatments. NKG2D is present on the surface of mucosal T cells. Its ligands are expressed on the epithelial cells exposed to the inflammatory factors. Blockade of NKG2D with a single dose anti-NKG2D mAb can significantly reduce the Crohn’s disease symptoms after 12 weeks, which suggests a new approach to CD treatment [95].
The resolution of symptoms is connected with lymphocytes cytotoxicity and cytokine release, as well as their migration and stopping proinflammatory cells. Anti-NKG2D mAb can modulate the lymphocyte and intestinal inflammation, which are two major processes in Crohn’s disease treatment [96]. This may decrease microenvironment cytotoxicity and killing of the target cells in the CD. The anti-NKG2D mAb seems to be the most commonly used form of treatment due to the high receptor prevalence and thanks to its limited side effects.
Xu et al. demonstrated that changes in expression of NKG2D on NK cells and NKG2DLs on endometrial cells are associated with pelvic endometriosis [23]. Low expression of ULBP-2 in patient’s endometrial cells results in lack of recognition by NK cells, which provides endometrium back to the pelvic cavity with the menstruation and escape the immune system detection Increased ULBP-2 receptor level in ectopic endometrial cells may be a result of autoimmune response.
Decidual NK cells are involved in uterine vascular remodeling, producing a variety of angiogenic cytokines, such as vascular endothelial growth factor, placental growth factor, TGF-β, angiopoietin 1 and 2 [97]. On the contrary, maternal NK cytotoxic activity prevents excessive trophoblast invasion. However, to avoid killing fetal trophoblast cells, the inhibitory signal must dominate the activating signal [98]. Increased NKG2D expression in RM patients can be the result of increased IL-15 expression in the decidua [99], or due to exposure of extravillous trophoblast cells to the oxidative stress in severely deficient trophoblast invasion, resulting in NKG2L expression [100][101].
The HNK phenotype with increased cytotoxic activity may result in killing trophoblast cells and increasing inflammatory cytokine production, leading to miscarriage. The rs2617170 TT genotype showed a significant association with protective effect against RM in north African fertile women [102]. The frequency of rs1049174 GG genotype and G allele was higher among Iranian women after recurrent miscarriages compared with controls [103]. Successful inhibition of the cytotoxic immune activity might be a therapeutic option, as well as intravenous immunoglobulin method in secondary RM cases, respectively [104].
4. Conclusions
The natural cytotoxic effect of the NK cells may be affected byKLRK1gene variability, showing also some association with the development of various diseases and treatment outcome. Due to its specific binding with ligands expressed on infected, neoplastic or stressed cells, NKG2D appears as a potentially interesting therapeutic agent in the treatment of cancer, autoimmune and viral diseases. As many studies confirmed, the NKG2D gene polymorphisms are related to immunogenetic susceptibility to specific types of cancer and autoimmune or viral-induced diseases. This gene also plays an important role in the transplant outcome and risk of post-transplantation complications development.
Due to its functionality and potential clinical application, NKG2D has been subject to various scientific and clinical studies, which in addition to research related to transplantation or cancer also include pioneering research on viral infections such as SARS-CoV-2. There are many clinical trials focusing on CAR-NK therapy targeted against solid tumors as well as hematological malignancies. New approaches to improve the efficacy and augment the NK cell infiltration of tumor have been made, including genetic manipulations (expressing IL-8 receptor [105] or tumor CD37 enzyme blockade [106]), using human pluripotent stem cells for better availability of the treatment [107] and improving the manufacturing conditions for a future clinical use [108]. In the near future, this novel form of therapy may find a clinical application in various cancers, including hematological malignancies as well as in GvHD and graft rejection treatments.
Despite the many different defense mechanisms of protection against cancer and infected cells, treatment with the NKG2D receptor show promising results. However, further clinical trials are necessary to confirm its efficacy and safety.
References
- Caligiuri, M.A. Human Natural Killer Cells. Blood 2008, 112, 461–469.
- Liu, H.; Wang, S.; Xin, J.; Wang, J.; Yao, C.; Zhang, Z. Role of NKG2D and Its Ligands in Cancer Immunotherapy. Am. J. Cancer Res. 2019, 9, 2064–2078.
- Stewart, C.A.; Vivier, E.; Colonna, M. Strategies of Natural Killer Cell Recognition and Signaling. In Immunobiology of Natural Killer Cell Receptors; Vivier, E., Colonna, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 298, pp. 1–21.
- Ljunggren, H.G.; Karre, K. In Search of the ‘Missing Self’: MHC Molecules and NK Cell Recognition. Immunol. Today 1990, 11, 237–244.
- Cooley, S.; Parham, P.; Miller, J.S. Strategies to Activate NK Cells to Prevent Relapse and Induce Remission Following Hematopoietic Stem Cell Transplantation. Blood 2018, 131, 1053–1062.
- Jelenčić, V.; Lenartić, M.; Wensveen, F.M.; Polić, B. NKG2D: A Versatile Player in the Immune System. Immunol. Lett. 2017, 189, 48–53.
- Vuletić, A.; Jurišić, V.; Jovanić, I.; Milovanović, Z.; Nikolić, S.; Konjević, G. Distribution of Several Activating and Inhibitory Receptors on CD3−CD56+ NK Cells in Regional Lymph Nodes of Melanoma Patients. J. Surg. Res. 2013, 183, 860–868.
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The Biology of Human Natural Killer-Cell Subsets. Trends Immunol. 2001, 22, 633–640.
- Mirjačić Martinović, K.M.; Babović, N.L.; Džodić, R.R.; Jurišić, V.B.; Tanić, N.T.; Konjević, G.M. Decreased Expression of NKG2D, NKp46, DNAM-1 Receptors, and Intracellular Perforin and STAT-1 Effector Molecules in NK Cells and Their Dim and Bright Subsets in Metastatic Melanoma Patients. Melanoma Res. 2014, 24, 295–304.
- Merkt, W.; Salzer, U.; Thiel, J.; Jandova, I.; Bergner, R.; Venhoff, A.C.; Venhoff, N. Blood CD3-(CD56 or 16)+ Natural Killer Cell Distributions Are Heterogeneous in Healthy Adults and Suppressed by Azathioprine in Patients with ANCA-Associated Vasculitides. BMC Immunol. 2021, 22, 26.
- Pende, D.; Falco, M.; Vitale, M.; Cantoni, C.; Vitale, C.; Munari, E.; Bertaina, A.; Moretta, F.; Del Zotto, G.; Pietra, G.; et al. Killer Ig-Like Receptors (KIRs): Their Role in NK Cell Modulation and Developments Leading to Their Clinical Exploitation. Front. Immunol. 2019, 10, 1179.
- Husain, Z.; Alper, C.A.; Yunis, E.J.; Dubey, D.P. Complex Expression of Natural Killer Receptor Genes in Single Natural Killer Cells. Immunology 2002, 106, 373–380.
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869.
- Biassoni, R.; Malnati, M.S. Human Natural Killer Receptors, Co-Receptors, and Their Ligands. Curr. Protoc. Immunol. 2018, 121.
- Brown, D.P.; Jones, D.C.; Anderson, K.J.; Lapaque, N.; Buerki, R.A.; Trowsdale, J.; Allen, R.L. The Inhibitory Receptor LILRB4 (ILT3) Modulates Antigen Presenting Cell Phenotype and, along with LILRB2 (ILT4), Is Upregulated in Response to Salmonella Infection. BMC Immunol. 2009, 10, 56.
- Borrego, F.; Masilamani, M.; Marusina, A.I.; Tang, X.; Coligan, J.E. The CD94/NKG2 Family of Receptors: From Molecules and Cells to Clinical Relevance. IR 2006, 35, 263–278.
- Dukovska, D.; Fernández-Soto, D.; Valés-Gómez, M.; Reyburn, H.T. NKG2H-Expressing T Cells Negatively Regulate Immune Responses. Front. Immunol. 2018, 9, 390.
- Spear, P.; Wu, M.-R.; Sentman, M.-L.; Sentman, C.L. NKG2D Ligands as Therapeutic Targets. Cancer Immun. 2013, 13, 8.
- Wensveen, F.M.; Jelenčić, V.; Polić, B. NKG2D: A Master Regulator of Immune Cell Responsiveness. Front. Immunol. 2018, 9, 441.
- González, S.; López-Soto, A.; Suarez-Alvarez, B.; López-Vázquez, A.; López-Larrea, C. NKG2D Ligands: Key Targets of the Immune Response. Trends Immunol. 2008, 29, 397–403.
- Campos-Silva, C.; Kramer, M.K.; Valés-Gómez, M. NKG2D-Ligands: Putting Everything under the Same Umbrella Can Be Misleading. HLA 2018, 91, 489–500.
- Schmiedel, D.; Mandelboim, O. NKG2D Ligands–Critical Targets for Cancer Immune Escape and Therapy. Front. Immunol. 2018, 9, 2040.
- Xu, H. Expressions of Natural Cytotoxicity Receptor, NKG2D and NKG2D Ligands in Endometriosis. J. Reprod. Immunol. 2019, 136, 102615.
- Diefenbach, A.; Hsia, J.K.; Hsiung, M.-Y.B.; Raulet, D.H. A Novel Ligand for the NKG2D Receptor Activates NK Cells and Macrophages and Induces Tumor Immunity. Eur. J. Immunol. 2003, 33, 381–391.
- Poggi, A.; Prevosto, C.; Massaro, A.-M.; Negrini, S.; Urbani, S.; Pierri, I.; Saccardi, R.; Gobbi, M.; Zocchi, M.R. Interaction between Human NK Cells and Bone Marrow Stromal Cells Induces NK Cell Triggering: Role of NKp30 and NKG2D Receptors. J. Immunol. 2005, 175, 6352–6360.
- Zhao, Y.; Chen, N.; Yu, Y.; Zhou, L.; Niu, C.; Liu, Y.; Tian, H.; Lv, Z.; Han, F.; Cui, J. Prognostic Value of MICA/B in Cancers: A Systematic Review and Meta-Analysis. Oncotarget 2017, 8, 96384–96395.
- Hayashi, T.; Imai, K.; Morishita, Y.; Hayashi, I.; Kusunoki, Y.; Nakachi, K. Identification of the NKG2D Haplotypes Associated with Natural Cytotoxic Activity of Peripheral Blood Lymphocytes and Cancer Immunosurveillance. Cancer Res. 2006, 66, 563–570.
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural Cytotoxic Activity of Peripheral-Blood Lymphocytes and Cancer Incidence: An 11-Year Follow-up Study of a General Population. Lancet 2000, 356, 1795–1799.
- Imai, K.; Hayashi, T.; Yamaoka, M.; Kajimura, J.; Yoshida, K.; Kusunoki, Y.; Nakachi, K. Effects of NKG2D Haplotypes on the Cell-Surface Expression of NKG2D Protein on Natural Killer and CD8 T Cells of Peripheral Blood among Atomic-Bomb Survivors. Hum. Immunol. 2012, 73, 686–691.
- Espinoza, J.L.; Nguyen, V.H.; Ichimura, H.; Pham, T.T.T.; Nguyen, C.H.; Pham, T.V.; Elbadry, M.I.; Yoshioka, K.; Tanaka, J.; Trung, L.Q.; et al. A Functional Polymorphism in the NKG2D Gene Modulates NK-Cell Cytotoxicity and Is Associated with Susceptibility to Human Papilloma Virus-Related Cancers. Sci. Rep. 2016, 6, 39231.
- Kabalak, G.; Thomas, R.M.; Martin, J.; Ortego-Centeno, N.; Jimenez-Alonso, J.; de Ramón, E.; Buyny, S.; Hamsen, S.; Gross, W.L.; Schnarr, S.; et al. Association of an NKG2D Gene Variant with Systemic Lupus Erythematosus in Two Populations. Hum. Immunol. 2010, 71, 74–78.
- Iwaszko, M.; Świerkot, J.; Kolossa, K.; Jeka, S.; Wiland, P.; Bogunia-Kubik, K. Influence of NKG2D Genetic Variants on Response to Anti-TNF Agents in Patients with Rheumatoid Arthritis. Genes 2018, 9, 64.
- Wielińska, J.; Tarassi, K.; Iwaszko, M.; Kościńska, K.; Wysoczańska, B.; Mole, E.; Kitsiou, V.; Świerkot, J.; Kolossa, K.; Kouniaki, D.; et al. Shared epitope and polymorphism of MICA and NKG2D encoding genes in Greek and Polish patients with rheumatoid arthritis. Cent. Eur. J. Immunol. 2021, 46.
- Tozatto-Maio, K.; Girot, R.; Ly, I.D.; Silva Pinto, A.C.; Rocha, V.; Fernandes, F.; Diagne, I.; Benzerara, Y.; Dinardo, C.L.; Soler, J.P.; et al. Polymorphisms in Inflammatory Genes Modulate Clinical Complications in Patients With Sickle Cell Disease. Front. Immunol. 2020, 11, 2041.
- Mariaselvam, C.M.; Tamouza, R.; Krishnamoorthy, R.; Charron, D.; Misra, D.P.; Jain, V.K.; Negi, V.S. Association of NKG2D Gene Variants with Susceptibility and Severity of Rheumatoid Arthritis. Clin. Exp. Immunol. 2017, 187, 369–375.
- Hara, R.; Onizuka, M.; Matsusita, E.; Kikkawa, E.; Nakamura, Y.; Matsushita, H.; Ohgiya, D.; Murayama, H.; Machida, S.; Ohmachi, K.; et al. NKG2D Gene Polymorphisms Are Associated with Disease Control of Chronic Myeloid Leukemia by Dasatinib. Int. J. Hematol. 2017, 106, 666–674.
- Asadi-Saghandi, A.; Shams, A.; Eslami, G.; Mirghanizadeh, S.A.; Eskandari-Nasab, E. Peginterferon Alfa-2a/Ribavirin Treatment Efficacy in Chronic Hepatitis C Patients Is Related to Natural Killer Group 2D Gene Rs1049174 GC Polymorphism. Virusdisease 2016, 27, 369–374.
- Espinoza, J.L.; Takami, A.; Onizuka, M.; Sao, H.; Akiyama, H.; Miyamura, K.; Okamoto, S.; Inoue, M.; Kanda, Y.; Ohtake, S.; et al. NKG2D Gene Polymorphism Has a Significant Impact on Transplant Outcomes after HLA-Fully-Matched Unrelated Bone Marrow Transplantation for Standard Risk Hematologic Malignancies. Haematologica 2009, 94, 1427–1434.
- Pu, X.; Hildebrandt, M.A.; Lu, C.; Roth, J.A.; Stewart, D.J.; Zhao, Y.; Heist, R.S.; Ye, Y.; Chang, D.W.; Su, L.; et al. Inflammation-Related Genetic Variations and Survival in Patients with Advanced Non-Small Cell Lung Cancer Receiving First-Line Chemotherapy. Clin. Pharmacol. Ther. 2014, 96, 360–369.
- Pasanen, A.; Karjalainen, M.K.; Kummola, L.; Waage, J.; Bønnelykke, K.; Ruotsalainen, M.; Piippo-Savolainen, E.; Goksör, E.; Nuolivirta, K.; Chawes, B.; et al. NKG2D Gene Variation and Susceptibility to Viral Bronchiolitis in Childhood. Pediatr. Res. 2018, 84, 451–457.
- Sikora, M.; Laayouni, H.; Menendez, C.; Mayor, A.; Bardaji, A.; Sigauque, B.; Netea, M.G.; Casals, F.; Bertranpetit, J. A Targeted Association Study of Immunity Genes and Networks Suggests Novel Associations with Placental Malaria Infection. PLoS ONE 2011, 6, e24996.
- Peralta-Zaragoza, O.; Bermúdez-Morales, V.; Gutiérrez-Xicotencatl, L.; Alcocer-González, J.; Recillas-Targa, F.; Madrid-Marina, V. E6 and E7 Oncoproteins from Human Papillomavirus Type 16 Induce Activation of Human Transforming Growth Factor Beta1 Promoter throughout Sp1 Recognition Sequence. Viral Immunol. 2006, 19, 468–480.
- Zheng, W.; Li, H.; Liu, B.; Wu, C. Association between the SNPs in Trace Element-Related Metabolic Genes and the Risk of Gastric Cancer: A Case-Control Study in Xianyou of China. J. Genet. 2019, 98.
- Wadsworth, C.A.; Dixon, P.H.; Taylor-Robinson, S.; Kim, J.U.; Zabron, A.A.; Wong, J.H.; Chapman, M.H.; McKay, S.C.; Spalding, D.R.; Wasan, H.S.; et al. Polymorphisms in Natural Killer Cell Receptor Protein 2D (NKG2D) as a Risk Factor for Cholangiocarcinoma. J. Clin. Exp. Hepatol. 2019, 9, 171–175.
- Gimeno, L.; Martínez-Banaclocha, H.; Bernardo, M.V.; Bolarin, J.M.; Marín, L.; López-Hernández, R.; López-Alvarez, M.R.; Moya-Quiles, M.R.; Muro, M.; Frias-Iniesta, J.F.; et al. NKG2D Polymorphism in Melanoma Patients from Southeastern Spain. Cancers 2019, 11, 438.
- Lin, F.; Dai, C.; Ge, X.; Tang, W.; Lin, Y.; Wang, Y.; Li, J. Prognostic Significance and Functional Implication of Immune Activating Receptor NKG2D in Gastric Cancer. Biochem. Biophys. Res. Commun. 2017, 487, 619–624.
- Liu, X.; Sun, M.; Yu, S.; Liu, K.; Li, X.; Shi, H. Potential Therapeutic Strategy for Gastric Cancer Peritoneal Metastasis by NKG2D Ligands-Specific T Cells. OncoTargets Ther. 2015, 8, 3095–3104.
- Sheppard, S.; Ferry, A.; Guedes, J.; Guerra, N. The Paradoxical Role of NKG2D in Cancer Immunity. Front. Immunol. 2018, 9, 1808.
- Wang, J.; Li, C.-D.; Sun, L. Recent Advances in Molecular Mechanisms of the NKG2D Pathway in Hepatocellular Carcinoma. Biomolecules 2020, 10, 301.
- Sheppard, S.; Guedes, J.; Mroz, A.; Zavitsanou, A.-M.; Kudo, H.; Rothery, S.M.; Angelopoulos, P.; Goldin, R.; Guerra, N. The Immunoreceptor NKG2D Promotes Tumour Growth in a Model of Hepatocellular Carcinoma. Nat. Commun. 2017, 8, 13930.
- Huergo-Zapico, L.; Acebes-Huerta, A.; López-Soto, A.; Villa-Álvarez, M.; Gonzalez-Rodriguez, A.P.; Gonzalez, S. Molecular Bases for the Regulation of NKG2D Ligands in Cancer. Front. Immunol. 2014, 5, 106.
- Groh, V.; Rhinehart, R.; Secrist, H.; Bauer, S.; Grabstein, K.H.; Spies, T. Broad Tumor-Associated Expression and Recognition by Tumor-Derived Gamma Delta T Cells of MICA and MICB. Proc. Natl. Acad. Sci. USA 1999, 96, 6879–6884.
- Madjd, Z.; Spendlove, I.; Moss, R.; Bevin, S.; Pinder, S.E.; Watson, N.F.; Ellis, I.; Durrant, L.G. Upregulation of MICA on highgrade invasive operable breast carcinoma. Cancer Immun. 2007, 7, 17.
- Chen, Y.; Lin, W.; Zhu, W.; Lin, J.; Zhou, Z.; Huang, C.; Chen, G.; Shi, Y.; Guo, Z.; Ye, Y. Tumor MICA Status Predicts the Efficacy of Immunotherapy with Cytokine-Induced Killer Cells for Patients with Gastric Cancer. Immunol. Res. 2016, 64, 251–259.
- Liu, C.-J.; Lui, M.-T.; Chen, H.-L.; Lin, S.-C.; Chang, K.-W. MICA and MICB Overexpression in Oral Squamous Cell Carcinoma: MICA and MICB Overexpression in OSCC. J. Oral Pathol. Med. 2006, 36, 43–47.
- Yamaguchi, K.; Chikumi, H.; Shimizu, A.; Takata, M.; Kinoshita, N.; Hashimoto, K.; Nakamoto, M.; Matsunaga, S.; Kurai, J.; Miyake, N.; et al. Diagnostic and Prognostic Impact of Serum-Soluble UL16-Binding Protein 2 in Lung Cancer Patients. Cancer Sci. 2012, 103, 1405–1413.
- Paschen, A.; Sucker, A.; Hill, B.; Moll, I.; Zapatka, M.; Nguyen, X.D.; Sim, G.C.; Gutmann, I.; Hassel, J.; Becker, J.C.; et al. Differential Clinical Significance of Individual NKG2D Ligands in Melanoma: Soluble ULBP2 as an Indicator of Poor Prognosis Superior to S100B. Clin. Cancer Res. 2009, 15, 5208–5215.
- Nückel, H.; Switala, M.; Sellmann, L.; Horn, P.A.; Dürig, J.; Dührsen, U.; Küppers, R.; Grosse-Wilde, H.; Rebmann, V. The Prognostic Significance of Soluble NKG2D Ligands in B-Cell Chronic Lymphocytic Leukemia. Leukemia 2010, 24, 1152–1159.
- Perez, C.; Prajapati, K.; Burke, B.; Plaza-Rojas, L.; Zeleznik-Le, N.J.; Guevara-Patino, J.A. NKG2D Signaling Certifies Effector CD8 T Cells for Memory Formation. J. Immunother. Cancer 2019, 7, 48.
- Ruggeri, L.; Capanni, M.; Urbani, E. Effectiveness of Donor Natural Killer Cell Alloreactivity in Mismatched Hematopoietic Transplants. Science 2002, 295, 2097–2100.
- Stadtmauer, E.A.; Fraietta, J.A.; Davis, M.M.; Cohen, A.D.; Weber, K.L.; Lancaster, E.; Mangan, P.A.; Kulikovskaya, I.; Gupta, M.; Chen, F.; et al. CRISPR-Engineered T Cells in Patients with Refractory Cancer. Science 2020, 367, eaba7365.
- Montagner, I.M.; Penna, A.; Fracasso, G.; Carpanese, D.; Dalla Pietà, A.; Barbieri, V.; Zuccolotto, G.; Rosato, A. Anti-PSMA CAR-Engineered NK-92 Cells: An Off-the-Shelf Cell Therapy for Prostate Cancer. Cells 2020, 9, 1382.
- Huergo-Zapico, L.; Acebes-Huerta, A.; Gonzalez-Rodriguez, A.P.; Contesti, J.; Gonzalez-García, E.; Payer, A.R.; Villa-Alvarez, M.; Fernández-Guizán, A.; López-Soto, A.; Gonzalez, S. Expansion of NK Cells and Reduction of NKG2D Expression in Chronic Lymphocytic Leukemia. Correlation with Progressive Disease. PLoS ONE 2014, 9, e108326.
- Jurisic, V.; Srdic, T.; Konjevic, G.; Markovic, O.; Colovic, M. Clinical Stage-Depending Decrease of NK Cell Activity in Multiple Myeloma Patients. Med. Oncol. 2007, 24, 312–317.
- Holthof, L.C.; Stikvoort, A.; van der Horst, H.J.; Gelderloos, A.T.; Poels, R.; Li, F.; Groen, R.W.J.; Zweegman, S.; van de Donk, N.W.C.J.; O’Dwyer, M.; et al. Bone Marrow Mesenchymal Stromal Cell-Mediated Resistance in Multiple Myeloma Against NK Cells Can Be Overcome by Introduction of CD38-CAR or TRAIL-Variant. HemaSphere 2021, 5, e561.
- Sandoval-Borrego, D.; Moreno-Lafont, M.C.; Vazquez-Sanchez, E.A.; Gutierrez-Hoya, A.; López-Santiago, R.; Montiel-Cervantes, L.A.; Ramírez-Saldaña, M.; Vela-Ojeda, J. Overexpression of CD158 and NKG2A Inhibitory Receptors and Underexpression of NKG2D and NKp46 Activating Receptors on NK Cells in Acute Myeloid Leukemia. Arch. Med. Res. 2016, 47, 55–64.
- Ashiru, O.; Boutet, P.; Fernández-Messina, L.; Agüera-González, S.; Skepper, J.N.; Valés-Gómez, M.; Reyburn, H.T. Natural Killer Cell Cytotoxicity Is Suppressed by Exposure to the Human NKG2D Ligand MICA*008 That Is Shed by Tumor Cells in Exosomes. Cancer Res. 2010, 70, 481–489.
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A Cell Initiating Human Acute Myeloid Leukaemia after Transplantation into SCID Mice. Nature 1994, 367, 645–648.
- Kim, J.; Chang, C.K.; Hayden, T.; Liu, F.-C.; Benjamin, J.; Hamerman, J.A.; Lanier, L.L.; Kang, S.-M. The Activating Immunoreceptor NKG2D and Its Ligands Are Involved in Allograft Transplant Rejection. J. Immunol. 2007, 179, 6416–6420.
- Yu, R.; Xu, S.; Wang, Y.; Cai, H.; Xu, P. Role of MICA Expression, Anti-MICA Antibodies and Serum MICA during Acute Rejection in a Rat-to-Mouse Cardiac Transplantation Model. Int. J. Clin. Exp. Pathol. 2015, 8, 14514–14520.
- Fabritius, C.; Ritschl, P.V.; Resch, T.; Roth, M.; Ebner, S.; Günther, J.; Mellitzer, V.; Nguyen, A.-V.; Pratschke, J.; Sauter, M.; et al. Deletion of the Activating NK Cell Receptor NKG2D Accelerates Rejection of Cardiac Allografts. Am. J. Transplant. 2017, 17, 3199–3209.
- Thompson, T.W.; Kim, A.B.; Li, P.J.; Wang, J.; Jackson, B.T.; Huang, K.T.H.; Zhang, L.; Raulet, D.H. Endothelial Cells Express NKG2D Ligands and Desensitize Antitumor NK Responses. eLife 2017, 6.
- Karimi, M.A.; Bryson, J.L.; Richman, L.P.; Fesnak, A.D.; Leichner, T.M.; Satake, A.; Vonderheide, R.H.; Raulet, D.H.; Reshef, R.; Kambayashi, T. NKG2D Expression by CD8+ T Cells Contributes to GVHD and GVT Effects in a Murine Model of Allogeneic HSCT. Blood 2015, 125, 3655–3663.
- Picardi, A.; Mengarelli, A.; Marino, M.; Gallo, E.; Benevolo, M.; Pescarmona, E.; Cocco, R.; Fraioli, R.; Tremante, E.; Petti, M.C.; et al. Up-Regulation of Activating and Inhibitory NKG2 Receptors in Allogeneic and Autologous Hematopoietic Stem Cell Grafts. J. Exp. Clin. Cancer Res. 2015, 34, 98.
- Koike, J.; Wakao, H.; Ishizuka, Y.; Sato, T.; Hamaoki, M.; Seino, K.; Koseki, H.; Nakayama, T.; Taniguchi, M. Bone Marrow Allograft Rejection Mediated by a Novel Murine NK Receptor, NKG2I. J. Exp. Med. 2004, 199, 137–144.
- Apithy, M.-J.; Charbonnier, A.; Desoutter, J.; Diouf, M.; Morel, P.; Garçon, L.; Marolleau, J.-P.; Guillaume, N. Impact of MICA and NKG2D Polymorphisms in HLA-Fully Matched Related and Unrelated Hematopoietic Stem Cell Transplantation. Bone Marrow Transplant. 2018, 53, 918–922.
- Stern-Ginossar, N.; Elefant, N.; Zimmermann, A.; Wolf, D.G.; Saleh, N.; Biton, M.; Horwitz, E.; Prokocimer, Z.; Prichard, M.; Hahn, G.; et al. Host Immune System Gene Targeting by a Viral MiRNA. Science 2007, 317, 376–381.
- Cosman, D.; Müllberg, J.; Sutherland, C.L.; Chin, W.; Armitage, R.; Fanslow, W.; Kubin, M.; Chalupny, N.J. ULBPs, Novel MHC Class I–Related Molecules, Bind to CMV Glycoprotein UL16 and Stimulate NK Cytotoxicity through the NKG2D Receptor. Immunity 2001, 14, 123–133.
- Eagle, R.A.; Traherne, J.A.; Hair, J.R.; Jafferji, I.; Trowsdale, J. ULBP6/RAET1L Is an Additional Human NKG2D Ligand. Eur. J. Immunol. 2009, 39, 3207–3216.
- Fielding, C.A.; Aicheler, R.; Stanton, R.J.; Wang, E.C.Y.; Han, S.; Seirafian, S.; Davies, J.; McSharry, B.P.; Weekes, M.P.; Antrobus, P.R.; et al. Two Novel Human Cytomegalovirus NK Cell Evasion Functions Target MICA for Lysosomal Degradation. PLoS Pathog. 2014, 10, e1004058.
- Sutherland, C.L.; Chalupny, N.J.; Cosman, D. The UL16-Binding Proteins, a Novel Family of MHC Class I-Related Ligands for NKG2D, Activate Natural Killer Cell Functions: ULBPs Activate Human NK Cells. Immunol. Rev. 2001, 181, 185–192.
- Rölle, A.; Mousavi-Jazi, M.; Eriksson, M.; Odeberg, J.; Söderberg-Nauclér, C.; Cosman, D.; Kärre, K.; Cerboni, C. Effects of Human Cytomegalovirus Infection on Ligands for the Activating NKG2D Receptor of NK Cells: Up-Regulation of UL16-Binding Protein (ULBP)1 and ULBP2 Is Counteracted by the Viral UL16 Protein. J. Immunol. 2003, 171, 902–908.
- Duan, S.; Guo, W.; Xu, Z.; He, Y.; Liang, C.; Mo, Y.; Wang, Y.; Xiong, F.; Guo, C.; Li, Y.; et al. Natural Killer Group 2D Receptor and Its Ligands in Cancer Immune Escape. Mol. Cancer 2019, 18, 29.
- Zdrenghea, M.T.; Telcian, A.G.; Laza-Stanca, V.; Bellettato, C.M.; Edwards, M.R.; Nikonova, A.; Khaitov, M.R.; Azimi, N.; Groh, V.; Mallia, P.; et al. RSV Infection Modulates IL-15 Production and MICA Levels in Respiratory Epithelial Cells. Eur. Respir. J. 2012, 39, 712–720.
- Velapasamy, S.; Dawson, C.W.; Young, L.S.; Paterson, I.C.; Yap, L.F. The Dynamic Roles of TGF-β Signalling in EBV-Associated Cancers. Cancers 2018, 10, 247.
- Ma, J.; Guo, X.; Wu, X.; Li, J.; Zhu, X.; Li, Z.; Li, J.; Pan, L.; Li, T.; Li, H.; et al. Association of NKG2D Genetic Polymorphism with Susceptibility to Chronic Hepatitis B in a Han Chinese Population. J. Med. Virol. 2010, 82, 1501–1507.
- Liu, W.-C.; Wu, I.-C.; Chiu, Y.-C.; Tseng, K.-C.; Chen, C.-Y.; Chiu, H.-C.; Cheng, P.-N.; Chang, T.-T. Genotyping of Immune-Related Loci Associated with Delayed HBeAg Seroconversion in Immune-Active Chronic Hepatitis B Patients. Antiviral Res. 2020, 176, 104719.
- Roszak, A.; Lianeri, M.; Jagodziński, P.P. Prevalence of the NKG2D Thr72Ala Polymorphism in Patients with Cervical Carcinoma. Genet. Test. Mol. Biomark. 2012, 16, 841–845.
- Moretta, L.; Biassoni, R.; Bottino, C.; Cantoni, C.; Pende, D.; Mingari, M.C.; Moretta, A. Human NK Cells and Their Receptors. Microbes Infect. 2002, 4, 1539–1544.
- Zhu, Y.; Jiang, M.; Gao, L.; Huang, X. Single Cell Analysis of ACE2 Expression Reveals the Potential Targets for 2019-nCoV. Preprints 2020, 2020020221.
- Van Eeden, C.; Khan, L.; Osman, M.S.; Cohen Tervaert, J.W. Natural Killer Cell Dysfunction and Its Role in COVID-19. Int. J. Mol. Sci. 2020, 21, 6351.
- Gupta, M.; Noel, G.J.; Schaefer, M.; Friedman, D.; Bussel, J.; Johann-Liang, R. Cytokine modulation with immune gamma-globulin in peripheral blood of normal children and its implications in Kawasaki disease treatment. J. Clin. Immunol. 2001, 21, 193–199.
- Piotrowski, P.; Lianeri, M.; Olesińska, M.; Jagodziński, P.P. Prevalence of the NKG2D Thr72Ala Polymorphism in Patients with Systemic Lupus Erythematosus. Mol. Biol. Rep. 2012, 39, 1343–1347.
- Ogasawara, K.; Hamerman, J.A.; Ehrlich, L.R.; Bour-Jordan, H.; Santamaria, P.; Bluestone, J.A.; Lanier, L.L. NKG2D Blockade Prevents Autoimmune Diabetes in NOD Mice. Immunity 2004, 20, 757–767.
- Vadstrup, K.; Bendtsen, F. Anti-NKG2D MAb: A New Treatment for Crohn’s Disease? Int. J. Mol. Sci. 2017, 18, 1997.
- Allez, M.; Petryka, R.; Skolnick, B.E.; Wisniewska-Jarosinska, M.A. Mo1213 Efficacy and Safety of NNC0142-0002, a Novel Human Monoclonal Antibody Targeting NKG2D: A Randomized, Double-Blind, Single-Dose Phase 2 Trial in Patients with Crohn’s Disease. Gastroenterology 2014, 146 (Suppl. S1), S-587.
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK Cells Regulate Key Developmental Processes at the Human Fetal-Maternal Interface. Nat. Med. 2006, 12, 1065–1074.
- Beaman, K.D.; Ntrivalas, E.; Mallers, T.M.; Jaiswal, M.K.; Kwak-Kim, J.; Gilman-Sachs, A. Immune Etiology of Recurrent Pregnancy Loss and Its Diagnosis. Am. J. Reprod. Immunol. 2012, 67, 319–325.
- Toth, B.; Jeschke, U.; Rogenhofer, N.; Scholz, C.; Würfel, W.; Thaler, C.J.; Makrigiannakis, A. Recurrent miscarriage: Current concepts in diagnosis and treatment. J. Reprod. Immunol. 2010, 85, 25–32.
- Mei, B.; Zhang, S.-R.; Chen, Y.-L.; Wang, C.-F. Defects in NKG2D Ligand Expression Result in Failed Tolerance Induction at the Maternal–Fetal Interface: A Possible Cause for Recurrent Miscarriage. Med. Hypotheses 2012, 79, 465–467.
- Wallace, A.E.; Goulwara, S.S.; Whitley, G.S.; Cartwright, J.E. Oxygen modulates human decidual natural killer cell surface receptor expression and interactions with trophoblasts. Biol. Reprod. 2014, 91, 134.
- Hizem, S.; Mtiraoui, N.; Massaoudi, S.; Fortier, C.; Boukouaci, W.; Kahina, A.; Charron, D.; Mahjoub, T.; Tamouza, R. Polymorphisms in Genes Coding for the NK-Cell Receptor NKG2D and Its Ligand MICA in Recurrent Miscarriage. Am. J. Reprod. Immunol. 2014, 72, 577–585.
- Asl, A.A.; Nezamdoust, F.V.; Fesahat, F.; Astani, A.; Barati, M.; Raee, P.; Asadi-Saghandi, A. Association between Rs1049174 NKG2D Gene Polymorphism and Idiopathic Recurrent Spontaneous Abortion in Iranian Women: A Case-Control Study. J. Obstet. Gynaecol. 2020, 1–5.
- Egerup, P.; Lindschou, J.; Gluud, C.; Christiansen, O.B.; ImmuReM IPD Study Group. The Effects of Intravenous Immunoglobulins in Women with Recurrent Miscarriages: A Systematic Review of Randomised Trials with Meta-Analyses and Trial Sequential Analyses Including Individual Patient Data. PLoS ONE 2015, 10, e0141588.
- Ng, Y.Y.; Tay, J.C.K.; Wang, S. CXCR1 Expression to Improve Anti-Cancer Efficacy of Intravenously Injected CAR-NK Cells in Mice with Peritoneal Xenografts. Mol. Ther. Oncolytics 2020, 16, 75–85.
- Wang, J.; Lupo, K.B.; Chambers, A.M.; Matosevic, S. Purinergic Targeting Enhances Immunotherapy of CD73+ Solid Tumors with PiggyBac-Engineered Chimeric Antigen Receptor Natural Killer Cells. J. Immunother. Cancer 2018, 6, 136.
- Li, Y.; Hermanson, D.L.; Moriarity, B.S.; Kaufman, D.S. Human IPSC-Derived Natural Killer Cells Engineered with Chimeric Antigen Receptors Enhance Anti-Tumor Activity. Cell Stem Cell 2018, 23, 181–192.e5.
- Oberschmidt, O.; Morgan, M.; Huppert, V.; Kessler, J.; Gardlowski, T.; Matthies, N.; Aleksandrova, K.; Arseniev, L.; Schambach, A.; Koehl, U.; et al. Development of Automated Separation, Expansion, and Quality Control Protocols for Clinical-Scale Manufacturing of Primary Human NK Cells and Alpharetroviral Chimeric Antigen Receptor Engineering. Hum. Gene Ther. Methods 2019, 30, 102–120.


