Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Metabolic syndrome is characterized by abdominal obesity, high blood pressure, insulin resistance, a proinflammatory and prothrombotic state, atherogenic dyslipidemia , and low high-density lipoprotein cholesterol (HDL-C). Metabolic syndrom is mediated by oxidative stress and inflammation. Although the mechanisms are complicated and not very clear, this entry elucidates the mechanisms and pathways that drive oxidative stress in contributing to metabolic syndrome.
- oxidative stress
- inflammatory cytokines
- metabolic syndrome
- obesity
1. Introduction
Metabolic syndrome is characterized by the presence of several interconnected risk factors for type 2 diabetes (T2DM) and cardiovascular disease (CVD) [1]. The presence of metabolic syndrome increases the risk of developing T2DM by 5 fold, CVD by 2 fold, and the risk of all-cause mortality by 1.5 fold [2][3]. Metabolic syndrome is highly prevalent in the United States, with about 35% prevalence and almost half in those aged 65 years and older [2][4]. The risk factors of metabolic syndrome include increased waist circumference or belly fat, high plasma triglycerides, elevated blood pressure, high blood sugar, and low plasma high density lipoprotein (HDL) [1]. If a patient has three of the five major risk factors, a diagnosis of metabolic syndrome is made [5].
Although many factors contribute to the pathophysiology of metabolic syndrome, several studies show that oxidative stress, in conjunction with chronic inflammatory conditions, is at the core of the development of metabolic diseases [4][6][7]. The imbalance between oxidants and antioxidants, which is often tilted in favor of the oxidants, is what causes oxidative stress, which in turn causes a disruption in redox signaling and regulation as well as molecular and cellular damage [8][9]. Metabolic syndrome is characterized by obesity-related problems, indicating a relationship between obesity and metabolic syndrome [10]. Inflammation and oxidative stress play a significant role in the development of metabolic comorbidities such as hyperlipidemia, high blood pressure, and increased glucose intolerance, all of which lead to metabolic dysfunction [10][11]. Several studies have shown that the risk for metabolic syndrome can be greatly reversed by reducing body weight and focusing interventions on dietary changes such as time-restricted eating, special diets such as the Mediterranean diet, including increasing physical exercise, sleep changes, or even reduce stress [2][3][5][12].
2. Metabolic Syndrome Components
Metabolic syndrome is characterized by abdominal obesity, high blood pressure, insulin resistance (a risk factor for T2DM), a proinflammatory and prothrombotic state, and atherogenic dyslipidemia (high triglycerides, high apolipoprotein B, high low-density lipoprotein particle (LDL-p) number, and low high-density lipoprotein cholesterol (HDL-C) [3][5][13][14]. The components of metabolic syndrome are interrelated, as briefly described below. Obesity is the primary risk factor in the development of T2DM, and it is estimated that roughly 90 percent of people who have T2DM are either overweight or obese [15][16]. Obesity is also linked to an increased risk of CVD, which includes conditions such as high blood pressure, atherosclerosis, acute myocardial infarction, and heart failure [4]. Central obesity is defined as having an abdominal circumference that is greater than 102 cm for males and greater than 88 cm for women [17]. Central obesity is one of the most important factors in the etiology of metabolic syndrome, including insulin resistance [18]. Obesity is associated with low-grade inflammation, which may lead to insulin resistance, insulin deficiency, and metabolic disturbances [4]. Insulin, a peptide hormone released by pancreatic beta cells in response to rising blood glucose, blocks lipolysis, and hepatic gluconeogenesis, and increased glucose absorption in the liver, muscles, and adipose tissues [19]. Insulin resistance is an altered physiologic response to insulin stimulation of the target tissues, such as the liver, muscle, and adipose tissue, and the resistance hinders glucose metabolism, resulting in hypertrophy of beta cells, increased beta-cell insulin production, and hyperinsulinemia [20][21]. Insulin resistance may result in hyperglycemia, hypertension, dyslipidemia, visceral obesity, hyperuricemia, increased inflammatory markers, endothelial dysfunction, and thrombosis, which may lead to metabolic syndrome, nonalcoholic fatty liver disease (NAFLD), and T2DM through several complex mechanisms [21][22][23].
Lipid metabolism is critical to the etiology of insulin resistance and the subsequent development of metabolic syndrome [24]. Lipid changes also contribute to the diagnostic criteria for metabolic syndrome, and the two major lipids include fasting triglyceridemia >150 mg/dL and HDL cholesterol concentration <40 mg/dL. This lipid derangement is characterized by an increase in the synthesis of very low-density lipoproteins (VLDL), a decrease in the plasma’s lipolytic capacity, and an increased cholesterol ester transfer protein activity [18]. Patients with metabolic syndrome also exhibit hemostatic changes that can elevate the risk of both atherothrombotic and thromboembolic cardiovascular disease [18][25]. The atherothrombotic and thromboembolic changes result from endothelial dysfunction, which may be caused by chronic inflammation, dyslipidemia, and hypertension [18][25].
3. Mechanisms of Reactive Oxygen Species and Their Role in the Development and Progression of Metabolic Syndrome
ROS production is tightly regulated by redox signaling and sensing mechanisms [26] (Figure 1).
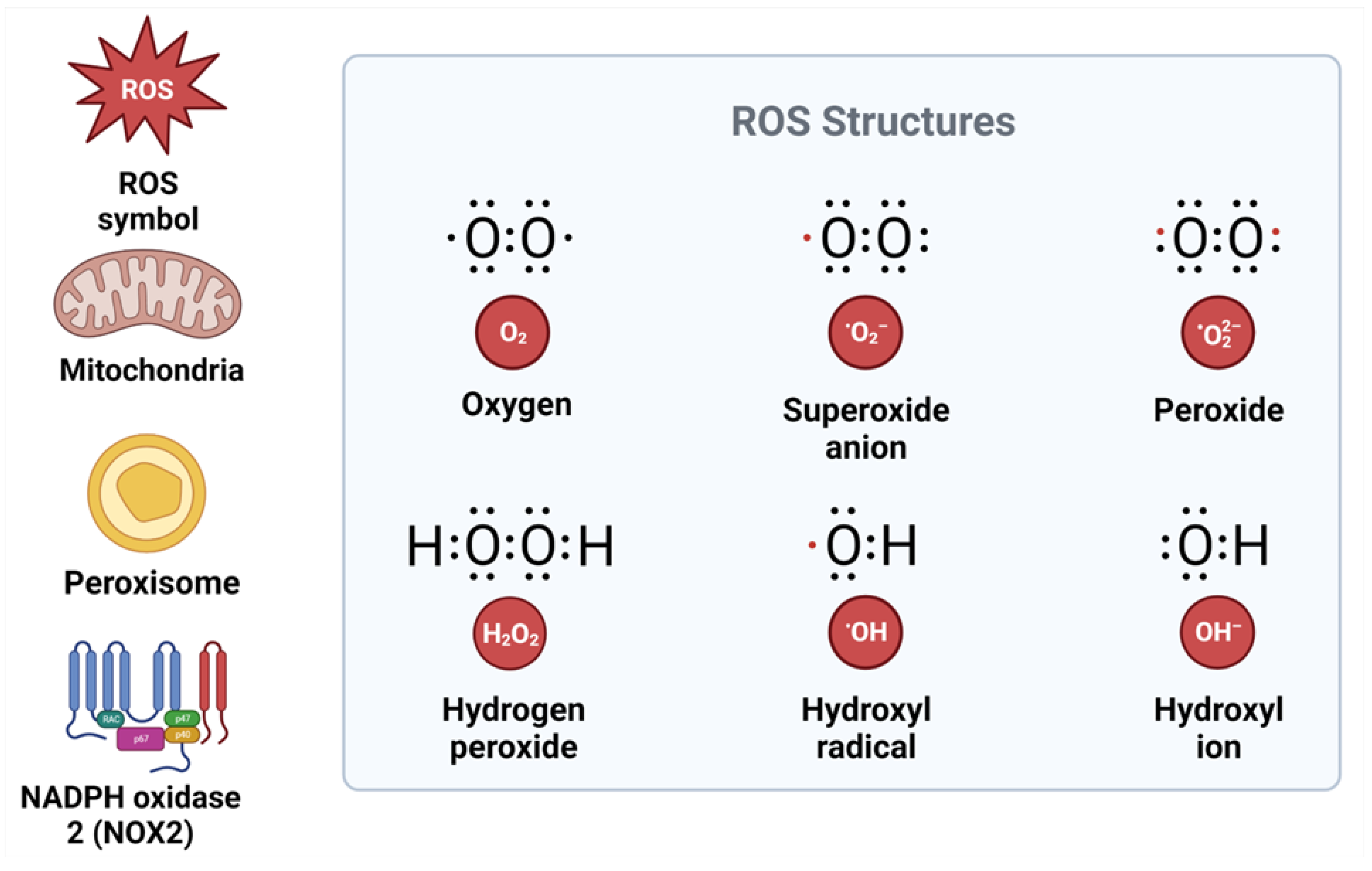
Figure 1. Structure of reactive oxygen species and their sources. ROS, reactive oxygen species; NADPH, nicotinamide adenine dinucleotide phosphate; NOX2, NADPH oxidase.
ROS signaling may participate in normal physiological processes or contribute to maladaptive responses that result in metabolic dysfunction and inflammatory signaling, depending on the ROS source, cell type, and tissue environment [9][27][28]. The two major sources of ROS inside the cell are nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) enzymes and the mitochondria [29]. The NOX enzymes are a family of enzymes (NOX1, NOX2, NOX3, NOX4, NOX5, DUOX1, and DUOX2) located in the cell membrane, and NOX2-NOX3 is important in most pathological conditions [29]. In the mitochondria, ROS are formed during oxidative phosphorylation by oxidizing reduced nicotinamide adenine dinucleotide (NADH) to NAD+ [30][31]. The superoxide anion that is produced by the mitochondria and NOX2 is rapidly converted by an enzyme called superoxide dismutase into hydrogen peroxide (H2O2), which serves as a signaling molecule [32][33] (Figure 2). Hydrogen peroxide is a powerful oxidizing agent. For this reason, cells express antioxidant proteins, including peroxiredoxin, catalase, glutathione (GSH), and thioredoxin, that convert H2O2 to water [31][34]. The level of H2O2 must be strictly maintained; hence, its production must be equal to its reduction [9]. High H2O2 in the presence of free ferric iron (Fe2+) produces hydroxyl radicals (•OH) in the Fenton reaction [9][28].
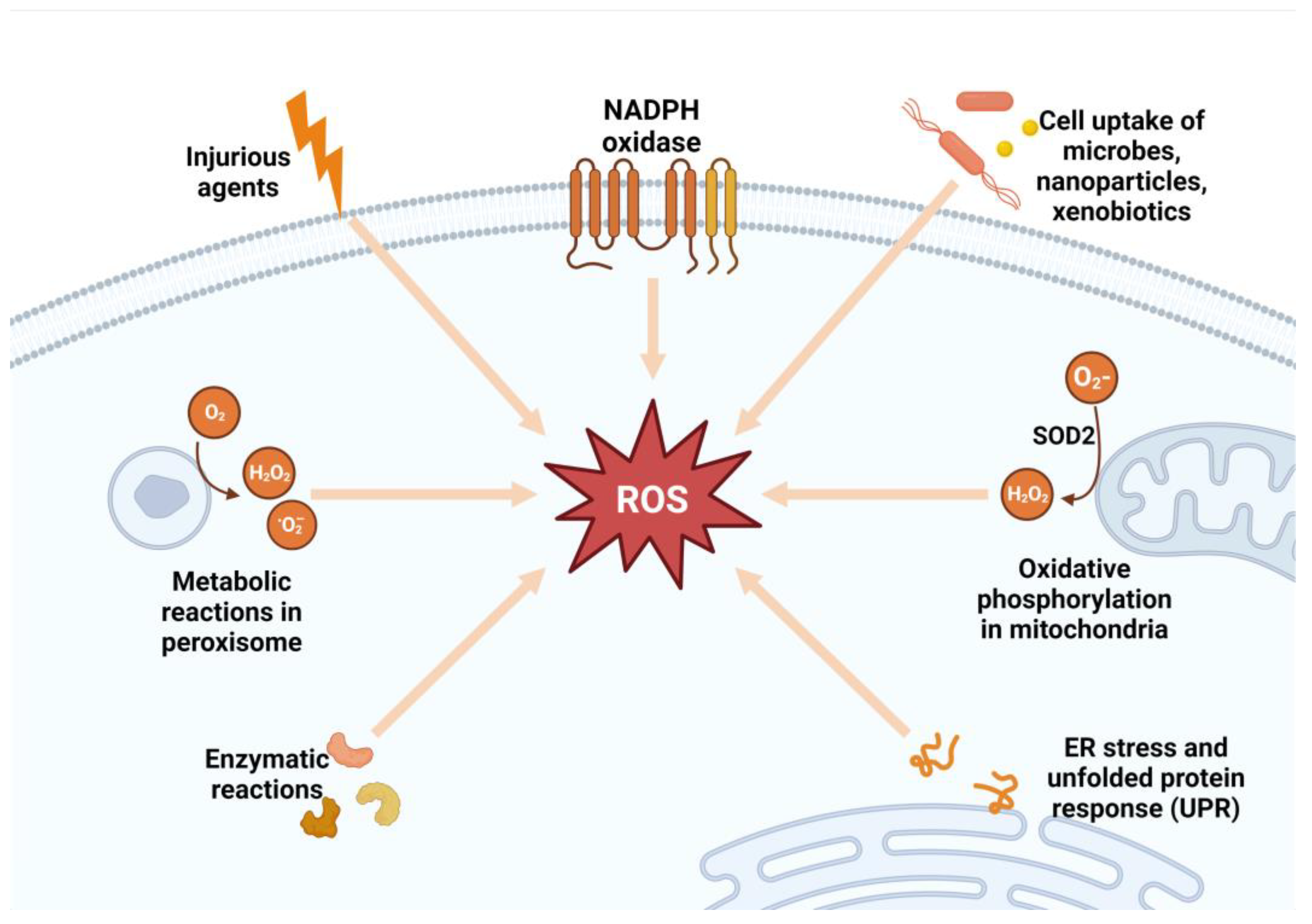
Figure 2. Sources of reactive oxygen species. SOD2, superoxide dismutase 2; ROS, reactive oxygen species; ER, endoplasmic reticulum; NADPH, nicotinamide adenine dinucleotide phosphate; H2O2, hydrogen peroxide.
Through tightly controlled redox regulation, signaling, and sensing, ROS are essential for normal biological functions in physiologic settings [35]. Oxidative posttranslational modification (Ox-PTM), also known as oxidative protein modification, is a crucial molecular process that regulates proteins, which eventually affect the biological responses of cells [36]. Redox-sensitive proteins include ion transporters, receptors, signaling molecules, transcription factors, cytoskeletal structural proteins, and matrix metalloproteases [9]. Proteins are normally targets of reversible Ox-PTM; however, in pathological conditions associated with oxidative stress, such as hypertension, proteins undergo irreversible Ox-PTM, which results in a loss of protein function and, as a consequence, cell damage, tissue injury, and failure of the target organs [37][38]. ROS, such as H2O2, are also essential for the activation of cellular pathways, including those that interact with vasoactive drugs such as angiotensin II (Ang II), endothelin-1 (ET-1), aldosterone, and prostanoids used to mediate cellular effects, and those that regulate intracellular calcium homeostasis [9]. ROS activate transcription factors such as hypoxia-inducible factor (HIF) that regulates angiogenesis, activate the phosphoinositide 3 kinase (PI3K) pathway that regulates cellular growth, the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) pathway, which in normal conditions prevents apoptosis by regulating cell survival, activates the mitogen-activated protein kinase (MAPK) pathway, which regulates cellular proliferation [39]. ROS also stimulate the transcription of pro-inflammatory chemokine and cytokine production and the recruitment and activation of inflammatory and immune cells [40][41].
Overproduction of ROS can occur in pathological disorders such as obesity, insulin resistance, hyperglycemia, chronic inflammation, and dyslipidemia [40][42][43]. Oxidative stress is detrimental because all the excess ROS induces cellular damage, specifically damaging DNA and peroxidize lipids [44]. Lipids present in plasma, mitochondrial, and endoplasmic reticulum membranes are major targets of ROS attack and peroxidation in most macromolecules [30][44]. The end products of lipid peroxidation, known as lipid peroxides, can be toxic to a cell and require removal by glutathione through an elusive mechanism [45]. Many studies have found that metabolic syndrome patients had lower plasma antioxidant enzyme activity and greater biomarkers of oxidative damage than healthy individuals, which may contribute to oxidative stress [46]. In the same manner, proteins and nucleic acids can be subject to peroxidation as well as nitrosylation [31]. Nevertheless, these end products are not usually directly toxic to the cell [31]. However, accumulation of inactive proteins can overload the ability of a cell to metabolize them and hence lead to the damage of DNA as they are capable of activating apoptosis [45]. In addition, the accumulation of modified proteins decreases their function, leading to a severe loss of normal cell activity [4][27][32][45]. The overproduction of ROS results in an oxidative stress environment, which also destabilizes redox signaling and control and leads to deleterious effects on gene expression, increases growth factors and stress response elements, and activates the apoptosis pathway [9][27]. The disrupted redox signaling also promotes pro-inflammatory and pro-fibrotic pathways, which affect insulin metabolic signaling and endothelial dysfunction and promote cardiovascular and renal inflammation and fibrosis, which contribute to target organ damage [9][47]. The mechanisms of ROS and their role in the development of metabolic syndrome are shown in Figure 3.
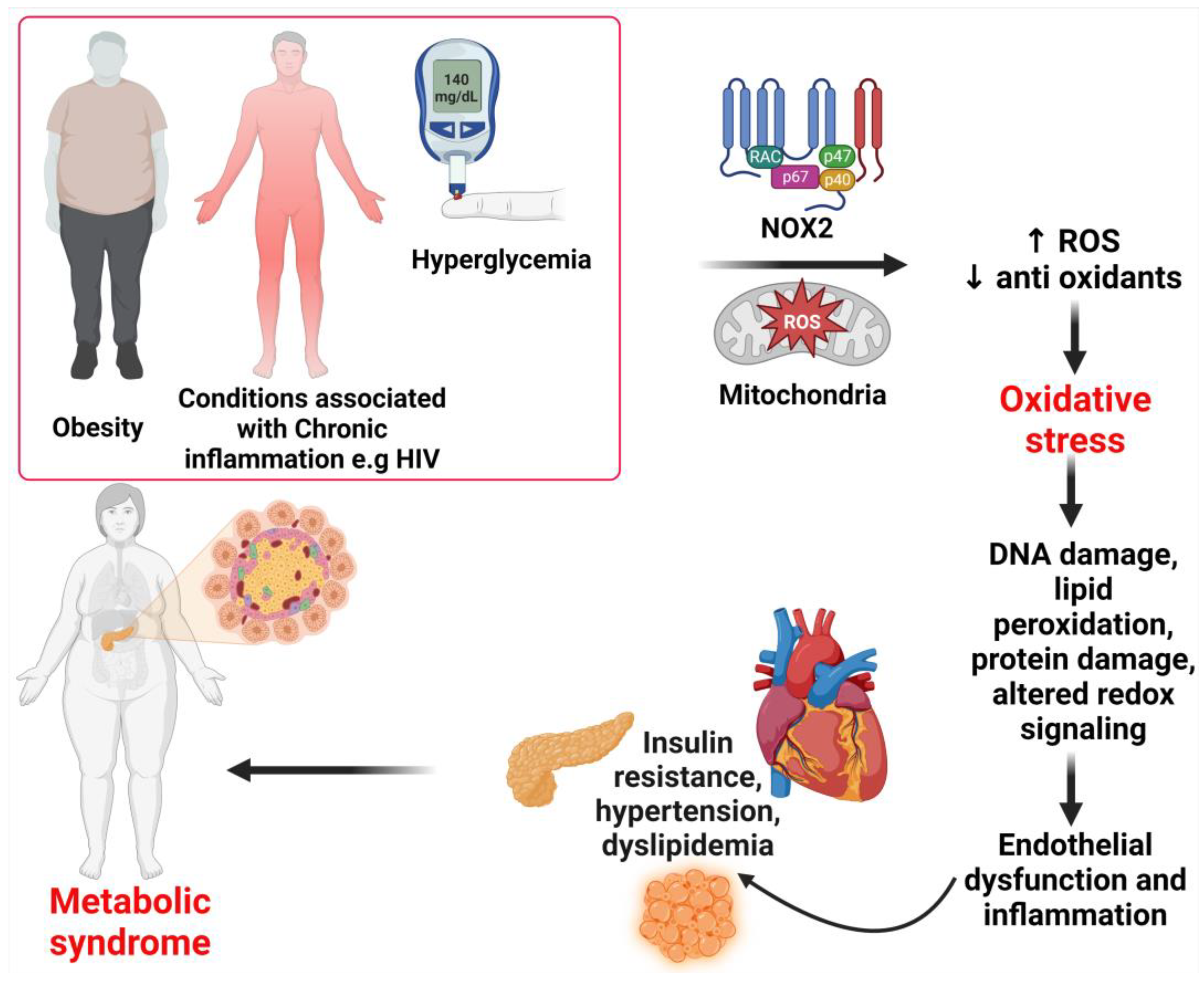
Figure 3. Mechanisms of metabolic syndrome. Under pathological conditions such as obesity, chronic inflammation, and hyperglycemia, excessive ROS generation can occur. ROS production occurs through the activation of enzymes in the cytosol, membrane, and mitochondria. An increase in the production of ROS and the depletion of antioxidants result in oxidative stress. The resulting oxidative stress leads to intracellular cell damage and altered redox, which leads to the irreversible accumulation of oxidation products, promoting endothelial dysfunction, which leads to insulin resistance, hypertension, dyslipidemia, and, subsequently, metabolic syndrome. ROS, reactive oxygen species; NOX2, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) enzymes.
This entry is adapted from the peer-reviewed paper 10.3390/ijms24097898
References
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic Syndrome: Pathophysiology, Management, and Modulation by Natural Compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225.
- Gallardo-Alfaro, L.; Bibiloni, M.d.M.; Mascaró, C.M.; Montemayor, S.; Ruiz-Canela, M.; Salas-Salvadó, J.; Corella, D.; Fitó, M.; Romaguera, D.; Vioque, J.; et al. Leisure-Time Physical Activity, Sedentary Behaviour and Diet Quality Are Associated with Metabolic Syndrome Severity: The PREDIMED-Plus Study. Nutrients 2020, 12, 1013.
- Wilkinson, M.J.; Manoogian, E.N.C.; Zadourian, A.; Lo, H.; Fakhouri, S.; Shoghi, A.; Wang, X.; Fleischer, J.G.; Navlakha, S.; Panda, S.; et al. Ten-Hour Time-Restricted Eating Reduces Weight, Blood Pressure, and Atherogenic Lipids in Patients with Metabolic Syndrome. Cell Metab. 2020, 31, 92–104.e5.
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Capó, X.; Bouzas, C.; Mateos, D.; Pons, A.; Tur, J.A.; Sureda, A. Metabolic Syndrome Is Associated with Oxidative Stress and Proinflammatory State. Antioxidants 2020, 9, 236.
- Dieli-Conwright, C.M.; Courneya, K.S.; Demark-Wahnefried, W.; Sami, N.; Lee, K.; Buchanan, T.A.; Spicer, D.V.; Tripathy, D.; Bernstein, L.; Mortimer, J.E. Effects of Aerobic and Resistance Exercise on Metabolic Syndrome, Sarcopenic Obesity, and Circulating Biomarkers in Overweight or Obese Survivors of Breast Cancer: A Randomized Controlled Trial. J. Clin. Oncol. 2018, 36, 875–883.
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 8267234.
- Franco, C.; Sciatti, E.; Favero, G.; Bonomini, F.; Vizzardi, E.; Rezzani, R. Essential Hypertension and Oxidative Stress: Novel Future Perspectives. Int. J. Mol. Sci. 2022, 23, 14489.
- Sies, H. Oxidative Stress: A Concept in Redox Biology and Medicine. Redox Biol. 2015, 4, 180–183.
- Touyz, R.M.; Rios, F.J.; Alves-Lopes, R.; Neves, K.B.; Camargo, L.L.; Montezano, A.C. Oxidative Stress: A Unifying Paradigm in Hypertension. Can. J. Cardiol. 2020, 36, 659.
- Han, T.S.; Lean, M.E. A Clinical Perspective of Obesity, Metabolic Syndrome and Cardiovascular Disease. JRSM Cardiovasc. Dis. 2016, 5, 2048004016633371.
- Todowede, O.O.; Sartorius, B. Prevalence of Metabolic Syndrome, Discrete or Comorbid Diabetes and Hypertension in Sub-Saharan Africa among People Living with HIV versus HIV-Negative Populations: A Systematic Review and Meta-Analysis Protocol. BMJ Open 2017, 7, e016602.
- Kunduraci, Y.E.; Ozbek, H. Does the Energy Restriction Intermittent Fasting Diet Alleviate Metabolic Syndrome Biomarkers? A Randomized Controlled Trial. Nutrients 2020, 12, 3213.
- Jeong, J.; Suh, Y.J. Association between Serum Uric Acid and Metabolic Syndrome in Koreans. J. Korean Med. Sci. 2019, 34, e307.
- Thomas, M.S.; Huang, L.; Garcia, C.; Sakaki, J.R.; Blesso, C.N.; Chun, O.K.; Fernandez, M.L. The Effects of Eggs in a Plant-Based Diet on Oxidative Stress and Inflammation in Metabolic Syndrome. Nutrients 2022, 14, 2548.
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications. Circ. Res. 2016, 118, 1723–1735.
- Grant, B.; Sandelson, M.; Agyemang-Prempeh, B.; Zalin, A. Managing Obesity in People with Type 2 Diabetes. Clin. Med. 2021, 21, e327–e331.
- Schroder, J.D.; Falqueto, H.; Mânica, A.; Zanini, D.; de Oliveira, T.; de Sá, C.A.; Cardoso, A.M.; Manfredi, L.H. Effects of Time-Restricted Feeding in Weight Loss, Metabolic Syndrome and Cardiovascular Risk in Obese Women. J. Transl. Med. 2021, 19, 3.
- Pedro-Botet, J.; Ascaso, J.F.; Barrios, V.; De la Sierra, A.; Escalada, J.; Millán, J.; Mostaza, J.M.; Pérez-Martínez, P.; Pintó, X.; Salas-Salvadó, J.; et al. COSMIC Project: Consensus on the Objectives of the Metabolic Syndrome in Clinic. Diabetes Metab. Syndr. Obes. Targets Ther. 2018, 11, 683–697.
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786.
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223.
- Wolosowicz, M.; Prokopiuk, S.; Kaminski, T.W. Recent Advances in the Treatment of Insulin Resistance Targeting Molecular and Metabolic Pathways: Fighting a Losing Battle? Medicina 2022, 58, 472.
- Kaur, J. A Comprehensive Review on Metabolic Syndrome. Cardiol. Res. Pract. 2014, 2014, 943162.
- Marušić, M.; Paić, M.; Knobloch, M.; Liberati Pršo, A.-M. NAFLD, Insulin Resistance, and Diabetes Mellitus Type 2. Can. J. Gastroenterol. Hepatol. 2021, 2021, 6613827.
- Denisenko, Y.K.; Kytikova, O.Y.; Novgorodtseva, T.P.; Antonyuk, M.V.; Gvozdenko, T.A.; Kantur, T.A. Lipid-Induced Mechanisms of Metabolic Syndrome. J. Obes. 2020, 2020, 5762395.
- Otamas, A.; Grant, P.J.; Ajjan, R.A. Diabetes and Atherothrombosis: The Circadian Rhythm and Role of Melatonin in Vascular Protection. Diab. Vasc. Dis. Res. 2020, 17, 1479164120920582.
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462.
- Samson, S.L.; Garber, A.J. Metabolic Syndrome. Endocrinol. Metab. Clin. N. Am. 2014, 43, 1–23.
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive Oxygen Species—Sources, Functions, Oxidative Damage. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2020, 48, 124–127.
- DeVallance, E.; Li, Y.; Jurczak, M.J.; Cifuentes-Pagano, E.; Pagano, P.J. The Role of NADPH Oxidases in the Etiology of Obesity and Metabolic Syndrome: Contribution of Individual Isoforms and Cell Biology. Antioxid. Redox Signal. 2019, 31, 687–709.
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive Oxygen Species (ROS) Homeostasis and Redox Regulation in Cellular Signaling. Cell Signal. 2012, 24, 981–990.
- Zhu, J.; Wu, F.; Yue, S.; Chen, C.; Song, S.; Wang, H.; Zhao, M. Functions of Reactive Oxygen Species in Apoptosis and Ganoderic Acid Biosynthesis in Ganoderma Lucidum. FEMS Microbiol. Lett. 2019, 366, fnaa015.
- Veith, A.; Moorthy, B. Role of Cytochrome P450s in the Generation and Metabolism of Reactive Oxygen Species. Curr. Opin. Toxicol. 2018, 7, 44–51.
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial Electron Transport Chain: Oxidative Phosphorylation, Oxidant Production, and Methods of Measurement. Redox Biol. 2020, 37, 101674.
- Beckhauser, T.F.; Francis-Oliveira, J.; De Pasquale, R. Reactive Oxygen Species: Physiological and Physiopathological Effects on Synaptic Plasticity. J. Exp. Neurosci. 2016, 10, 23–48.
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.-A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793.
- Chung, H.S.; Wang, S.-B.; Venkatraman, V.; Murray, C.I.; Van Eyk, J.E. Cysteine Oxidative Post-Translational Modifications: Emerging Regulation in the Cardiovascular System. Circ. Res. 2013, 112, 382–392.
- Baba, S.P.; Bhatnagar, A. Role of thiols in oxidative stress. Curr. Opin. Toxicol. 2018, 7, 133–139.
- Tomin, T.; Schittmayer, M.; Honeder, S.; Heininger, C.; Birner-Gruenberger, R. Irreversible Oxidative Post-Translational Modifications in Heart Disease. Expert Rev. Proteom. 2019, 16, 681–693.
- Zhang, H.; Xu, R.; Wang, Z. Contribution of Oxidative Stress to HIF-1-Mediated Profibrotic Changes during the Kidney Damage. Oxid. Med. Cell Longev. 2021, 2021, 6114132.
- Padgett, L.E.; Broniowska, K.A.; Hansen, P.A.; Corbett, J.A.; Tse, H.M. The Role of Reactive Oxygen Species and Proinflammatory Cytokines in Type 1 Diabetes Pathogenesis. Ann. N. Y. Acad. Sci. 2013, 1281, 16–35.
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167.
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902.
- Čolak, E.; Pap, D. The Role of Oxidative Stress in the Development of Obesity and Obesity-Related Metabolic Disorders. J. Med. Biochem. 2021, 40, 1–9.
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642.
- Gaschler, M.M.; Stockwell, B.R. Lipid Peroxidation in Cell Death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425.
- Bekkouche, L.; Bouchenak, M.; Malaisse, W.J.; Yahia, D.A. The Mediterranean Diet Adoption Improves Metabolic, Oxidative, and Inflammatory Abnormalities in Algerian Metabolic Syndrome Patients. Horm. Metab. Res. Horm. Stoffwechselforschung Horm. Metab. 2014, 46, 274–282.
- Scioli, M.G.; Storti, G.; D’Amico, F.; Rodríguez Guzmán, R.; Centofanti, F.; Doldo, E.; Céspedes Miranda, E.M.; Orlandi, A. Oxidative Stress and New Pathogenetic Mechanisms in Endothelial Dysfunction: Potential Diagnostic Biomarkers and Therapeutic Targets. J. Clin. Med. 2020, 9, 1995.
This entry is offline, you can click here to edit this entry!