Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Laccase belongs to the superfamily of multicopper oxidases and has been widely investigated in recent decades. Due to its mild and efficient oxidation of substrates, laccase has been successfully applied in organic catalytic synthesis, the degradation of harmful substances, and other green catalytic fields. Adding a mediator not only effectively improves the reaction efficiency of laccase but also expands the scope of the substrate.
- laccase
- laccase–mediator system
- green catalysis
1. Introduction
Laccase (EC 1.10.3.2) is a copper-containing polyphenol oxidase that belongs to the blue copper oxidase (MCO) family [1][2]. It was first discovered in the Japanese lacquer tree Rhus Vernicifera [3][4]. Subsequently, laccases were found in different plant species [5], microbes [6][7], and animals [8]. There have been more investigations on microbial laccases than on animal and plant laccases, for which there have been relatively few. Microbial laccases are divided into fungal laccases and bacterial laccases. Bacterial laccase mainly plays a role in melanin production, spore wall defense, morphological change, and copper ion detoxification [9][10]. Fungal laccase is mainly related to pigment generation, plant disease, and lignin degradation [11][12]. Plant laccase is closely related to lignin biosynthesis [13]. At the same time, the primary function of animal laccase protein is to control the ossification of the epidermis [14].
Laccase is a glycoprotein with a molecular mass ranging from 50 to 140 kDa. Their amino acid sequence can span from 220 to 800 amino acids and may contain three cupredoxin-like domains. These domains bind copper centers involved in intermolecular electron transfer reactions and constitute the catalytic core of laccases (Figure 1) [15]. The active copper center of laccase generally contains four copper ions: a type I copper ion (T1-Cu), a type II copper ion (T2-Cu), and two type III copper ions (T3-Cu). T1-Cu is a mononuclear center that can gain electrons from the substrate and then transfer them to the trinuclear cluster (TNC), and the oxidation of the substrate occurs there. T2-Cu is a single-electron acceptor, whereas T3-Cu forms coupled ion pairs and is a double-electron acceptor. T2-Cu and T3-Cu together form a trinuclear cluster (TNC). Oxygen accepts four electrons and four protons to form water, which joins the bulk solvent [16][17]. Laccases can perform the single-electron oxidation of the substrate without using hydrogen peroxide while reducing molecular oxygen to water; therefore, they have a surprisingly broad substrate spectrum and can oxidize simple diphenols, polyphenols, diamines, and aromatic amines. The optimum temperature and pH of laccases depend on the enzyme source and substrate properties; those ranges are, respectively, from 20 °C to 75 °C and 3 to 8 (Table 1).
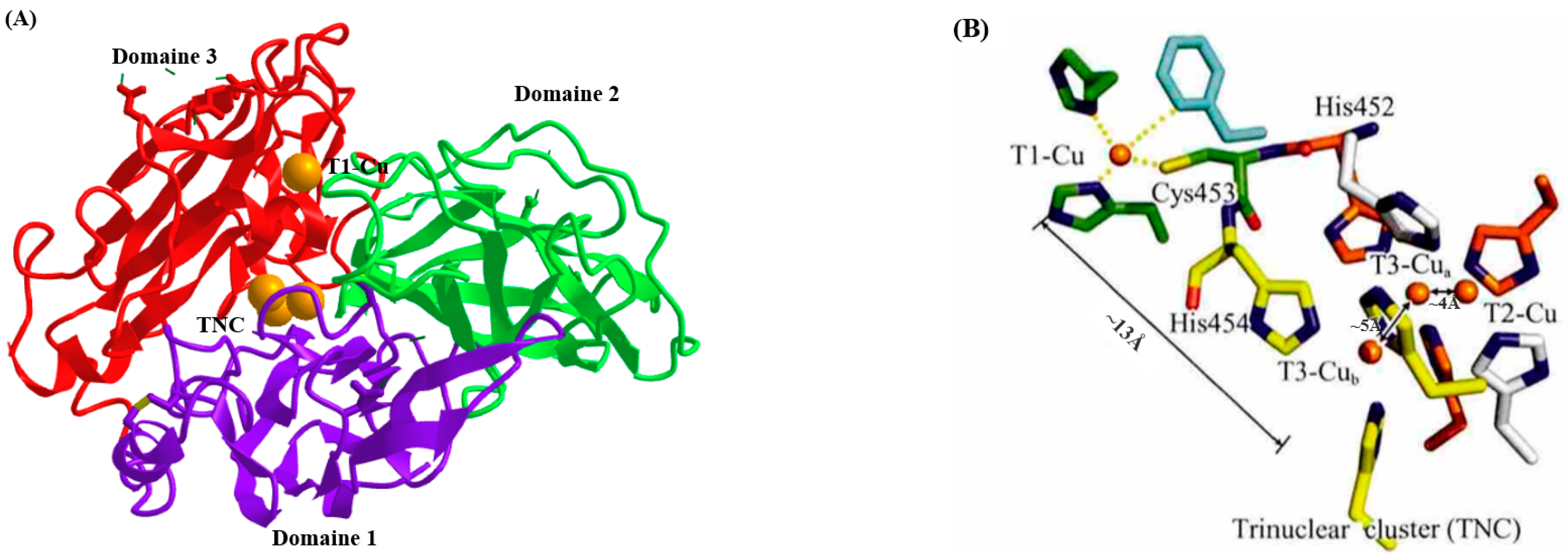
Figure 1. Crystal structure of the laccase (LccI) (PDB code: 1GYC) in Trametes versicolor. (A) The overall structure of LccI; (B) structure of the schematic representation of the four copper sites in LccI.
Table 1. Enzymatic properties of laccase from different sources.
As an oxidase, the ability of laccase to oxidize the substrate is directly related to its redox potential (E0) [33][34]. E0 is an important characteristic of the catalytic oxidation capacity of laccase, which is the energy required for laccase to capture an electron from a reducing substrate. The E0 is critical to the reactivity of laccase and the overall reaction characteristics. Laccase can directly oxidize substrates with low E0, whereas some mediators are needed to assist laccase in oxidizing substrates with high E0. Adding a mediator not only effectively improves the reaction efficiency of laccase but also expands the scope of the substrate. For instance, with the mediator’s help, laccase can oxidize nonphenolic structures with high E0 and is used in pulp bleaching [35]. Recently, laccase and the laccase–mediator system (LMS) have received extensive attention in green catalysis, such as synthesizing complex organic compounds, the selective modification of natural products, and the degradation of harmful substances [36][37][38][39]. For example, the C–N bond breakage of amines catalyzed by laccase is essential for synthesizing amino acids and nucleosides [40][41]. Existing research indicates that using Pleurotus ostreatus laccase and its natural mediator (syringaldehyde) to catalyze C–C bond breakage results in the removal of up to 100% and 85% of BPA at concentrations of 0.44 and 0.88 mmol/L in wastewater within 1 h [42].
2. The Effect of the Mediator System on Laccase Catalysis
Currently, some problems still need to be solved urgently to directly apply laccase to industrial production. For instance: (1) Numerous substrates cannot directly bind to laccase specifically. (2) With laccase, it is difficult to oxidize nonphenolic compounds with high E0 (E0 > 1.3 V) due to its low E0 (E0 < 0.8 V). Thus, the development of laccase in industries such as lignin degradation and bio-bleaching is limited [43][44]. In order to reduce the oxidation potential of substrates and improve the oxidation efficiency, some mediators can be used as an intermediate substrate for laccase to form new intermediate states to transport electrons [41][45]. These mediators are compounds with low molecular mass and low E0, such as 2,2′-Azino-bis-(3-ethylbenzothiazoline-sulphonate) (ABTS) and 2,2,6,6-Tetramethyl-1-piperidinylox (TEMPO), which can easily gain and lose electrons. They can form highly active and stable intermediates under the action of laccase and act on the substrate to is oxidized. In the LMS, laccase first oxidizes the mediator into a free radical. The oxidized mediator rapidly applies to compounds above the E0 of laccase and to those polymers that cannot directly access the laccase active center [46][47][48] (Figure 2).
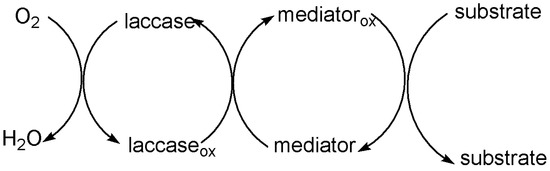
Figure 2. Oxidation of substrates by LMS.
Laccase mediators are usually divided into artificial and natural mediators (Figure 3) [49]. Due to their high efficiency and inexpensive availability, artificial mediators are widely used in lignin degradation, polycyclic aromatic hydrocarbon (PAH) oxidation, and dye decolorization. Common artificial mediators include ABTS, TEMPO, and 1-hydroxy-benzotriazole (HBT) (Figure 3B) [50][51][52]. Three mechanisms have been proposed for the function of mediators in the LMS: (1) hydrogen atom transfer (HAT), (2) electron transfer (ET), and (3) the ionic mechanism (IM). ABTS was the first mediator found to promote the laccase-catalyzed oxidation of nonphenolic lignin. The action mechanism of ABTS belongs to the electron transfer mechanism (ET), which undergoes two stages (Figure 4): forming an ABTS+· cationic radical and slowly oxidizing to ABTS2+. ABTS2+ with higher reduction potential (but not ABTS+·) performs a more critical function in the laccase–ABTS system, which mediates the oxidation of nonphenolic lignin substrates [53][54][55]. The HAT mechanism, which is generally the oxidation mechanism mediated by the N-OH type mediator, uses a form of nitryl (>N-O·) to perform oxidation, such as the HBT system. Meanwhile, the purpose of the ion mechanism (IM) is mainly to form an ammonium oxide ion (>N=O·) through the nitryl group (>N-O·) to carry out the oxidation, such as the TEMPO system [39][56].
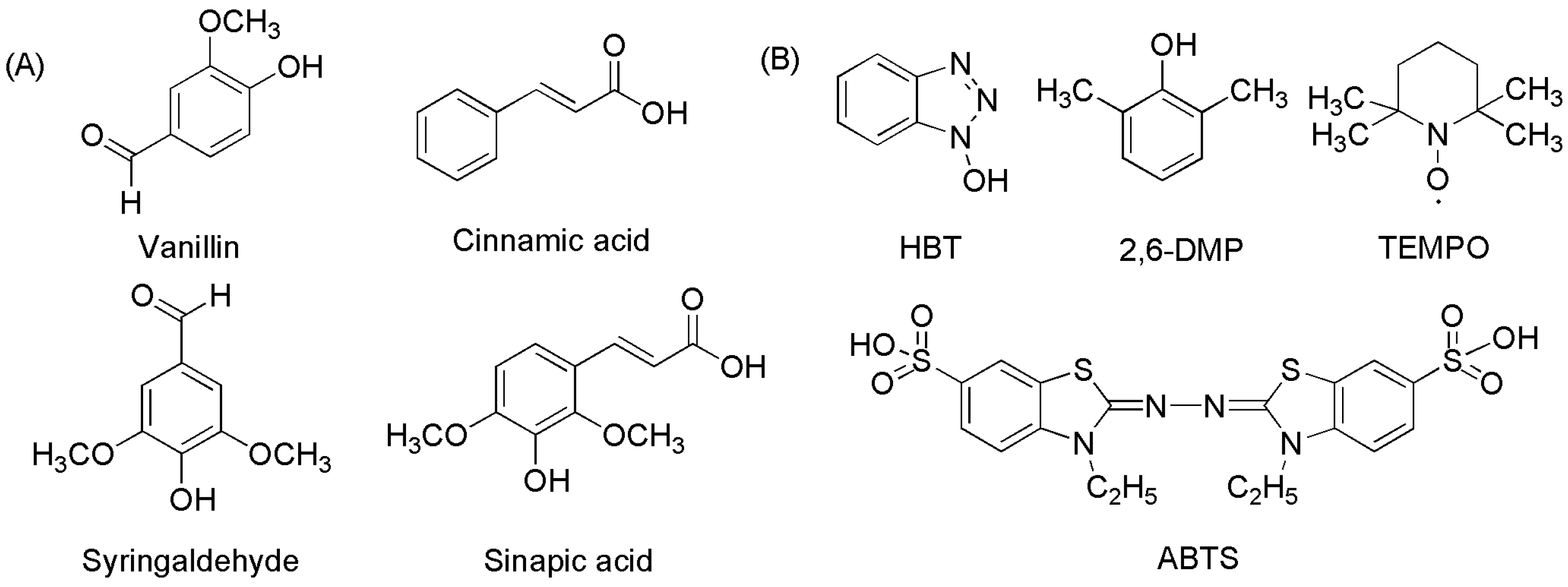
Figure 3. The chemical structures of several artificial and natural redox mediators in laccase-catalyzed oxidation reaction systems. (A) natural redox mediators; (B) artificial redox mediators.
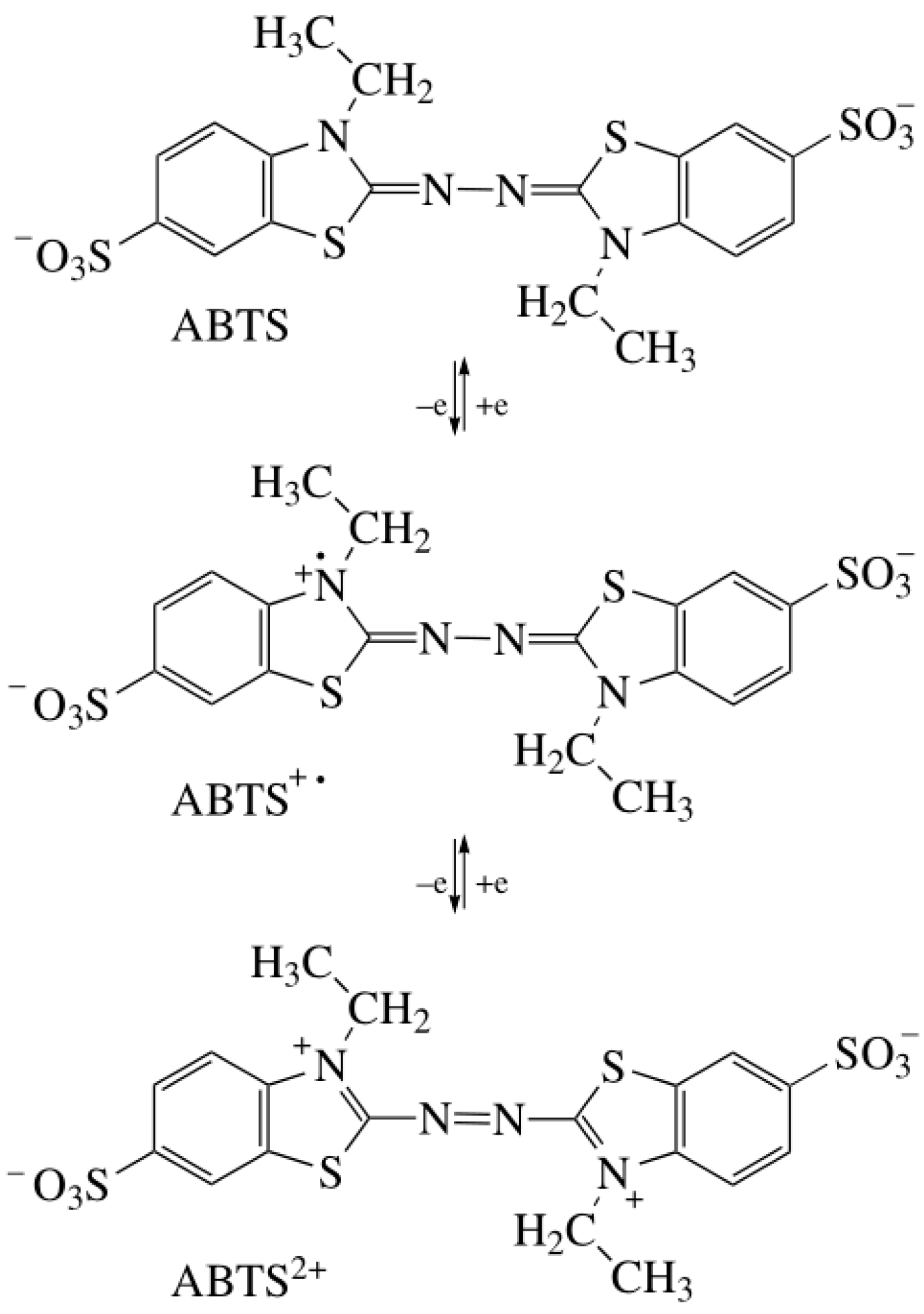
Figure 4. Oxidation of ABTS by Laccase.
Artificial mediators have potential applications in the areas of lignin degradation and polycyclic aromatic hydrocarbon (PAH) oxidation for dye decolorization, but some disadvantages limit their use. For instance: (1) poor stability, (2) potential toxicity [57], and (3) difficulty in regeneration when the molar ratio of mediators to substrates is as high as 40:1 [53][58]. Compared to artificial mediators, natural mediators have more economic value because they are readily obtained, environmentally friendly, and reproducible (Figure 3A) [35][53]. Some fungal metabolites and lignin derivatives could be used as natural mediators of laccase, including but not limited to vanillin, acetyl vanillin, acetosyringone, syringaldehyde, 2,4,6-trimethyl phenol, and p-coumaric acid [59]. Taking the laccase p-coumaric acid system as an example, it can remove 95% anthracene (80% with HBT) and benzoin anthracene within 24 h [57].
Besides indirectly assisting laccase-catalyzing substrates, mediators show synergism with each other, and the degradation efficiency increases with the increase in mediator concentration [60]. For example, the complex mediator system composed of laccase, ABTS, and HBT can oxidize phenanthrene with only one intermediary phase with a degradation rate that can be increased by 30–40% compared with a single-mediator system (such as the ABTS system or HBT system) [61]. Therefore, with intensive research on and development of the LMS, the biocatalytic substrates of laccase can be further expanded.
This entry is adapted from the peer-reviewed paper 10.3390/catal13040750
References
- Ozer, A.; Ay Sal, F.; Belduz, A.O.; Kirci, H.; Canakci, S. Use of feruloyl esterase as laccase-mediator system in paper bleaching. Appl. Biotech. Biochem. 2020, 190, 721–731.
- Nakamura, K.; Go, N. Function and molecular evolution of multicopper blue proteins. Cell. Mol. Life Sci. 2005, 62, 2050–2066.
- Singh, G.; Arya, S.K. Utility of laccase in pulp and paper industry: A progressive step towards the green technology. Int. J. Biol. Macromol. 2019, 134, 1070–1084.
- Claus, H. Laccases: Structure, reactions, distribution. Micron 2004, 35, 93–96.
- Kagalkar, A.N.; Khandare, R.V.; Govindwar, S.P. Textile dye degradation potential of plant laccase significantly enhances upon augmentation with redox mediators. RSC Adv. 2015, 5, 80505–80517.
- Chauhan, P.S.; Goradia, B.; Saxena, A. Bacterial laccase: Recent update on production, properties and industrial applications. 3 Biotech 2017, 7, 323.
- Balaes, T. Ligninolytic enzyme system in ecological adaptation of lignicolous macrofungi. Appl. Ecol. Env. Res. 2017, 15, 207–224.
- Janusz, G.; Pawlik, A.; Swiderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkolazka, A.; Paszczynski, A. Laccase properties, physiological functions, and evolution. Int. J. Mol. Sci. 2020, 21, 966.
- Guan, Z.B.; Luo, Q.; Wang, H.R.; Chen, Y.; Liao, X.R. Bacterial laccases: Promising biological green tools for industrial applications. Cell. Mol. Life Sci. 2018, 75, 3569–3592.
- Singh, D.; Gupta, N. Microbial laccase: A robust enzyme and its industrial applications. Biologia 2020, 75, 1183–1193.
- Piscitelli, A.; Del Vecchio, C.; Faraco, V.; Giardina, P.; Macellaro, G.; Miele, A.; Pezzella, C.; Sannia, G. Fungal laccases: Versatile tools for lignocellulose transformation. Comptes Rendus Biol. 2011, 334, 789–794.
- Sousa, A.C.; Martins, L.O.; Robalo, M.P. Laccases: Versatile biocatalysts for the synthesis of heterocyclic cores. Molecules 2021, 26, 3719.
- Wang, J.; Feng, J.; Jia, W.; Chang, S.; Li, S.; Li, Y. Lignin engineering through laccase modification: A promising field for energy plant improvement. Biotechnol. Biofuels 2015, 8, 145.
- Asano, T.; Taoka, M.; Yamauchi, Y.; Craig Everroad, R.; Seto, Y.; Isobe, T.; Kamo, M.; Chosa, N. Re-examination of a α-chymotrypsin-solubilized laccase in the pupal cuticle of the silkworm, Bombyx mori: Insights into the regulation system for laccase activation during the ecdysis process. Insect. Biochem. Mol. Biol. 2014, 55, 61–69.
- Agrawal, K.; Chaturvedi, V.; Verma, P. Fungal laccase discovered but yet undiscovered. Bioresour. Bioprocess. 2018, 5, 4.
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. Cell. Mol. Life Sci. 2015, 72, 869–883.
- Malmstrom, B.G.; Agro, A.F.; Antonini, E. The mechanism of laccase-catalyzed oxidations: Kinetic evidence for the involvement of several electron-accepting sites in the enzyme. Eur. J. Biochem. 1969, 9, 383–391.
- Dong, J.L.; Zhang, Y.Z. Purification and characterization of two laccase isoenzymes from a ligninolytic fungus Trametes gallica. Prep. Biochem. Biotech. 2004, 34, 179–194.
- Chakroun, H.; Mechichi, T.; Martinez, M.J.; Dhouib, A.; Sayadi, S. Purification and characterization of a novel laccase from the ascomycete Trichoderma atroviride: Application on bioremediation of phenolic compounds. Process Biochem. 2010, 45, 507–513.
- Giovanelli, G.; Ravasini, G. Apple juice stabilization by combined enzyme-membrane filtration process. LWT Food Sci. Technol. 1993, 26, 1–7.
- Marques de Souza, C.G.; Peralta, R.M. Purification and characterization of the main laccase produced by the white-rot fungus Pleurotus pulmonarius on wheat bran solid state medium. J. Basic Microbiol. 2003, 43, 278–286.
- Martins, L.O.; Soares, C.M.; Pereira, M.M.; Teixeira, M.; Costa, T.; Jones, G.H.; Henriques, A.O. Molecular and biochemical characterization of a highly stable bacterial laccase that occurs as a structural component of the Bacillus subtilis endospore coat. J. Biol. Chem. 2002, 277, 18849–18859.
- Endo, K.; Hayashi, Y.; Hibi, T.; Hosono, K.; Beppu, T.; Ueda, K. Enzymological characterization of EpoA, a laccase-like phenol oxidase produced by Streptomyces griseus. J. Biochem. 2003, 133, 671–677.
- Molina-Guijarro, J.M.; Pérez Torres, J.; Muñoz Dorado, J.; Guillén Carretero, F.; Moya Lobo, R.; Hernández Cutuli, M.; Arias Fernández, M.E. Detoxification of azo dyes by a novel pH-versatile, salt-resistant laccase from Streptomyces ipomoea. Int. Microbiol. 2009, 12, 13–21.
- Niladevi, K.N.; Jacob, N.; Prema, P. Evidence for a halotolerant-alkaline laccase in Streptomyces psammoticus: Purification and characterization. Process Biochem. 2008, 43, 654–660.
- Grass, G.; Rensing, C. CueO is a multi-copper oxidase that confers copper tolerance in Escherichia coli. Biochem. Biophys. Res. Commun. 2001, 286, 902–908.
- Ranocha, P.; McDougall, G.; Hawkins, S.; Sterjiades, R.; Borderies, G.; Stewart, D.; Cabanes-Macheteau, M.; Boudet, A.M.; Goffner, D. Biochemical characterization, molecular cloning and expression of laccases–a divergent gene family–in poplar. Eur. J. Biochem. 1999, 259, 485–495.
- Sato, Y.; Bao, W.; Sederoff, R.; Whetten, R. Molecular cloning and expression of eight laccase cDNAs in loblolly pine (Pinus taeda)*. J. Plant. Res. 2001, 114, 147–155.
- Hüttermann, A.; Mai, C.; Kharazipour, A. Modification of lignin for the production of new compounded materials. Appl. Microbiol. Biotechnol. 2001, 55, 387–394.
- Dittmer, N.T.; Suderman, R.J.; Jiang, H.; Zhu, Y.C.; Gorman, M.J.; Kramer, K.J.; Kanost, M.R. Characterization of cDNAs encoding putative laccase-like multicopper oxidases and developmental expression in the tobacco hornworm, Manduca sexta, and the malaria mosquito, Anopheles gambiae. Insect Biochem. Mol. Biol. 2004, 34, 29–41.
- Niu, B.L.; Shen, W.F.; Liu, Y.; Weng, H.B.; He, L.H.; Mu, J.J.; Wu, Z.L.; Jiang, P.; Tao, Y.Z.; Meng, Z.Q. Cloning and RNAi-mediated functional characterization of MaLac2 of the pine sawyer, Monochamus alternatus. Insect Mol. Biol. 2008, 17, 303–312.
- Lang, M.; Kanost, M.R.; Gorman, M.P.O. Multicopper oxidase-3 is a laccase associated with the peritrophic matrix of Anopheles gambiae. PLoS ONE 2012, 7, e33985.
- Fabbrini, M.; Galli, C.; Gentili, P. Radical or electron-transfer mechanism of oxidation with some laccase/mediator systems. J. Mol. Catal. B-enzmy. 2002, 18, 169–171.
- Kunamneni, A.; Plou, F.J.; Ballesteros, A.; Alcalde, M. Laccases and their applications: A patent review. Recent Pat. Biotechnol. 2008, 2, 10–24.
- Sigoillot, C.; Record, E.; Belle, V.; Robert, J.L.; Levasseur, A.; Punt, P.J.; van den Hondel, C.A.M.J.J.; Fournel, A.; Sigoillot, J.C.; Asther, M. Natural and recombinant fungal laccases for paper pulp bleaching. Appl. Microbiol. Biotechnol. 2004, 64, 346–352.
- Mayolo-Deloisa, K.; Gonzalez-Gonzalez, M.; Rito-Palomares, M. Laccases in food industry: Bioprocessing, potential industrial and biotechnological applications. Front. Bioeng. Biotechnol. 2020, 8, 222.
- Lassouane, F.; Ait-Amar, H.; Amrani, S.; Rodriguez-Couto, S. A promising laccase immobilization approach for Bisphenol A removal from aqueous solutions. Bioresour. Technol. 2019, 271, 360–367.
- Parra Guardado, A.L.; Belleville, M.-P.; Rostro Alanis, M.d.J.; Parra Saldivar, R.; Sanchez-Marcano, J. Effect of redox mediators in pharmaceuticals degradation by laccase: A comparative study. Process Biochem. 2019, 78, 123–131.
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Yaropolov, Y.I. Laccase–mediator systems and their applications: A Review. Appl. Biochem. Microbiol. 2007, 43, 523–535.
- Martínez-Montero, L.; Díaz-Rodríguez, A.; Gotor, V.; Gotor-Fernández, V.; Lavandera, I. Broadening the chemical scope of laccases: Selective deprotection of N-benzyl groups. Green Chem. 2015, 17, 2794–2798.
- Verkade, J.M.M.; van Hemert, L.J.C.; Quaedflieg, P.J.L.M.; Schoemaker, H.E.; Schürmann, M.; van Delft, F.L.; Rutjes, F.P.J.T. Laccase-mediated deprotection of para-methoxyphenyl (PMP)-protected amines. Adv. Synth. Catal. 2007, 349, 1332–1336.
- Freitas, E.D.; Bubna, G.A.; Brugnari, T.; Kato, C.G.; Nolli, M.; Rauen, T.G.; Regina, D.; Peralta, R.A.; Bracht, A.; Souza, C.D. Removal of bisphenol A by laccases from Pleurotus ostreatus and Pleurotus pulmonarius and evaluation of ecotoxicity of degradation products. Chem. Eng. J. 2017, 330, 1361–1369.
- Cañas, A.I.; Camarero, S. Laccases and their natural mediators: Biotechnological tools for sustainable eco-friendly processes. Biotechnol. Adv. 2010, 28, 694–705.
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates: An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102.
- Könst, P.; Kara, S.; Kochius, S.; Holtmann, D.; Arends, I.W.; Ludwig, R.; Hollmann, F. Expanding the scope of laccase-mediator systems. ChemCatChem 2013, 5, 3027–3032.
- Bourbonnais, R.; Paice, M.; Freiermuth, B.; Bodie, E.; Borneman, S. Reactivities of various mediators and laccases with kraft pulp and lignin model compounds. Appl. Environ. Microbiol. 1997, 63, 4627–4632.
- Eggert, C.; Temp, U.; Dean, J.F.; Eriksson, K.-E.L. A fungal metabolite mediates degradation of non-phenolic lignin structures and synthetic lignin by laccase. FEBS Lett. 1996, 391, 144–148.
- Kawai, S.; Umezawa, T.; Higuchi, T. < original> Oxidation of methoxylated benzyl alcohols by laccase of Coriolus versicolor in the presence of syringaldehyde. Wood Res. Bull. Wood Res. Inst. Kyoto Univ. 1989, 76, 10–16.
- Park, S.; Jung, D.; Do, H.; Yun, J.; Lee, D.; Hwang, S.; Lee, S.H. Laccase-mediator system using a natural mediator as a whitening agent for the decolorization of melanin. Polymers 2021, 13, 3671.
- Valls, C.; Colom, J.F.; Baffert, C.; Gimbert, I.; Roncero, M.B.; Sigoillot, J.-C. Comparing the efficiency of the laccase–NHA and laccase–HBT systems in eucalyptus pulp bleaching. Biochem. Engin. J. 2010, 49, 401–407.
- Gu, Y.; Yuan, L.; Jia, L.; Xue, P.; Yao, H. Recent developments of a co-immobilized laccase–mediator system: A review. RSC Adv. 2021, 11, 29498–29506.
- Witayakran, S.; Ragauskas, A.J. Synthetic applications of laccase in green chemistry. Adv. Synth. Catal. 2009, 351, 1187–1209.
- Johannes, C.; Majcherczyk, A. Natural mediators in the oxidation of polycyclic aromatic hydrocarbons by laccase mediator systems. Appl. Environ. Microbiol. 2000, 66, 524–528.
- Matsumura, E.; Yamamoto, E.; Numata, A.; Kawano, T.; Shin, T.; Murao, S. Structures of the laccase-catalyzed oxidation products of hydroxybenzoic acids in the presence of ABTS . Agric. Biol. Chem. 1986, 50, 1355–1357.
- Zhao, Q.; Huang, W.; Luo, Z.; Liu, L.; Lu, Y.; Li, Y.; Li, L.; Hu, J.; Ma, H.; Chen, J. High-capacity aqueous zinc batteries using sustainable quinone electrodes. Sci. Adv. 2018, 4, eaao1761.
- Kudanga, T.; Nyanhongo, G.S.; Guebitz, G.M.; Burton, S. Potential applications of laccase-mediated coupling and grafting reactions: A review. Enzym. Microb. Technol. 2011, 48, 195–208.
- Li, K.; Helm, R.F.; Eriksson, K.E.L. Mechanistic studies of the oxidation of a non-phenolic lignin model compound by the laccase/1-hydroxybenzotriazole redox system. Biotechnol. Appl. Biochem. 1998, 27, 239–243.
- d’Acunzo, F.; Galli, C. First evidence of catalytic mediation by phenolic compounds in the laccase-induced oxidation of lignin models. Eur. J. Biochem. 2003, 270, 3634–3640.
- Blanquez, A.; Rodriguez, J.; Brissos, V.; Mendes, S.; Martins, L.O.; Ball, A.S.; Arias, M.E.; Hernandez, M. Decolorization and detoxification of textile dyes using a versatile Streptomyces laccase-natural mediator system. Saudi J. Biol. Sci. 2019, 26, 913–920.
- Vandertol-Vanier, H.; Vazquez-Duhalt, R.; Tinoco, R.; Pickard, M. Enhanced activity by poly (ethylene glycol) modification of Coriolopsis gallica laccase. J. Ind. Microbiol. Biotechnol. 2002, 29, 214–220.
- Han, M.-J.; Park, H.-T.; Song, H.-G. Degradation of phenanthrene by Trametes versicolor and its laccase. J. Microbiol. 2004, 42, 94–98.
This entry is offline, you can click here to edit this entry!