1. Introduction
Atopic dermatitis, allergic contact dermatitis and chronic spontaneous urticaria are common inflammatory conditions involving the skin. This inflammation may result from unique and, yet, shared pathogenic mechanisms, which may account for their coexistence and association in atopic patients.
1.1. Pathogenesis of Atopic Dermatitis
Atopic dermatitis (AD) is the most common inflammatory skin disorder in the developed world, with a lifetime prevalence of 15–20% [
1]. It is often associated with other atopic conditions, such as asthma, allergic rhinitis and chronic rhinosinusitis with nasal polyps [
2]. AD is a chronic, relapsing and inflammatory skin disease characterized by intense itching and excoriations with erythematous, xerotic, fissured and lichenificated skin, as well as an increased risk of skin infections [
3,
4]. The pathogenesis of AD has not been completely elucidated. Genetic heritage and its interactions with the environment contribute to epidermal skin barrier disruption, altered inflammatory response and skin microbiome dysbiosis [
5,
6], and oxidative stress also plays an important pathogenetic role [
7]. Recent studies have pointed to the key role of epigenetic changes in AD development, mainly mediated by DNA methylation, histone acetylation and the action of specific non-coding RNAs, including micro-RNAs (miRNAs), small interfering RNAs, long non-coding RNAs and Pivi-interacting RNAs [
8]. Skin barrier abnormalities include decreased filaggrin, loricrin, involucrin, ceramides, antimicrobial peptides and disorders of tight junctions [
9,
10]. In particular, filaggrin, a structural protein responsible for keratinization, moisturization and antimicrobial peptide functions, is mutated in 15–50% of patients with AD [
11,
12]. Skin barrier disruption stimulates keratinocytes and dendritic cells to produce activation-regulated chemokine (TARC), thymic stromal lymphopoietin (TSLP) and cytokines, including IL-1β, IL-25 and IL-33, which trigger type-2 inflammation, activating both innate and adaptative immunity [
13,
14]. Type-2 innate lymphoid cells and Th2 cells produce IL-4, IL-5 and IL-13 [
13], key mediators of type-2 response. They promote Th-naïve lymphocytes developing Th2 phenotypes, B lymphocyte isotypes switching from IgM to IgE [
15], eosinophil recruitment and activation [
16], itch exacerbation [
17] and skin remodeling toward fibrosis in the chronic disease phase [
1].
1.2. Pathogenesis of Allergic Contact Dermatitis
Allergic contact dermatitis (ACD) is a common inflammatory skin disorder characterized by pruritus, erythema, vesicles and scaling of the skin [
18]. ACD is common, with some studies demonstrating prevalence rates as high as 20% in the general population, and it accounts for the vast majority of occupational skin disorders in the Western world [
18,
19]. ACD involves type IV-mediated hypersensitivity to a specific allergen, resulting in an inflammatory response with exposure. The first phase is sensitization, when a person is first exposed to a hapten, which is defined as a low-molecular-weight antigen that, when bound to a larger carrier, can elicit an immune response.
After hapten has been recognized and phagocytized by dermal dendritic cells and Langerhans cells, the hapten–peptide complexes migrate to regional lymph nodes and induce the proliferation and circulation of hapten-specific T cells (Th1, Th2, Th17 and T regulatory cells) and the creation of effector and memory T cells. Clinical symptoms of ACD are produced in the elicitation phase, when re-exposure to the allergen, recognized by the sensitized hapten-specific T cells, causes an inflammatory cascade of cytokines and cellular infiltrates [
20]. Since the reaction to the allergen is not always immediate, with a possible delay of up to 72 h, it may be difficult to identify the culprit agents. Thorough questioning of the patient’s occupation, hobbies and any changes in personal products or clothing is helpful [
21], and certain distributions, such as on the eyelid, lateral face, central face, neck or hands suggest the consideration of ACD. Patch testing is the only practical, scientific and objective method to confirm the diagnosis of ACD.
1.3. Pathogenesis of Chronic Spontaneous Urticaria
Chronic spontaneous urticaria (CSU) is defined as the spontaneous appearance of itchy red wheals and/or angioedema for more than six weeks without any apparent cause [
22,
23], and it affects about 1.8% of the adult population [
24]. CSU is mainly caused by the activation of cutaneous mast cells (MCs), leading to the release of histamine and other mediators, such as platelet-activating factor (PAF) and cytokines, which results in vasodilatation, plasma extravasation and sensory nerve activation, as well as cell recruitment in the urticarial lesions (mainly T cells, eosinophils and basophils). Wheals are characterized by edema of the upper and mid-dermis, whereas similar changes may occur in the lower dermis, resulting in angioedema [
22]. The pathophysiology of CSU is not well understood, but there are two main pathogenetic mechanisms underlying the disease: dysregulation of intracellular signaling pathways within mast cells and basophils, which leads to defects in the trafficking or function of these cells, and development of autoantibodies to FcεRIα or IgE on both mast cells and basophils [
25]. Basophil numbers appear to be reduced in at least 50% of patients [
26], and they seem to be hyporeactive [
27]. Furthermore, their development is dependent on the interaction of stem cell factor (SCF) with the c-kit, as well as TGF-β produced by Tregs. In CSU, circulating Tregs (CD4 + CD25 + FOXP3+) are reduced and/or defective compared with normal cells [
28], and the reduced frequency of Tregs is consistent with an autoimmune hypothesis for CSU [
29,
30]. These events lead to mast cell degranulation and the predisposition to pathological mast cell activation if inappropriately upregulated. However, autoimmune theory is the most widely accepted hypothesis for explaining the pathogenesis of CSU, as it is thought that up to 45% of cases of CSU have autoimmune etiology [
25]. Specifically, CSU is associated with the development of circulating IgG antibodies against IgE and the high-affinity IgE receptor FcεR1; approximately 40% of these patients have circulating antibodies against one of the two targets [
31], with a higher frequency of positivity in CSU patients with a positive autologous serum skin test (ASST) [
32]. It seems likely that different pathomechanisms are interlinked in CSU rather than independent cascades [
33].
1.4. MicroRNAs
miRNAs represent an abundant class of small non-coding RNAs which regulate gene expression post-transcriptionally [
34]. miRNAs are short (20–22 nucleotides long), endogenous and single-stranded RNAs, whose genes [
35], highly conserved throughout evolution across species [
36], are located within the introns and exons of protein-coding genes [
37].
The first miRNAs were found in Caenorhabditis elegans in early 1990s [
34]; afterward, the first human miRNA was discovered in the 2000s [
38]. Currently, 2588 mature miRNAs have been identified for humans in miRbase [
39], and it is estimated that they modulate more than 60% of all human protein-coding genes by sequence-specific base pairing [
35]. In fact, it is widely recognized that one miRNA may regulate many genes as its targets, while one gene may be targeted by many miRNAs. [
40]
Primary transcripts of miRNAs are generated by RNA polymerase II and sequentially processed by ribonuclease III class enzymes [
37] (Drosha) to produce an miRNA precursor of 70 nucleotides (the pre-miRNA) [
41]. After nuclear processing, the pre-miRNA is transferred to the cytosol, where Dicer, another ribonuclease III class enzyme, cleaves it into mature double-stranded miRNA with 19–24 nucleotides. One strand of this mature miRNA duplex is then incorporated into the RNA-induced silencing complex (RISC) [
42] and serves as a guide for the recognition of target mRNAs [
43]. The other strand is unrolled from the guide strand and degraded, but, in some cases, both strands of the miRNA duplex are functional [
44]. The miRNA–RNA-induced silencing complexes interact with mRNA targets by sequence-specific base pairing, generally within the 3′-untranslated region of the target mRNAs [
45]. Two different mechanisms of RISC-mediated gene regulation exist, and they depend on the level of miRNA–mRNA complementarity: at sites with extensive complementarity, the miRNA can mediate mRNA cleavage; however, sites with a lower degree of complementarity may lead to translational repression or mRNA destabilization [
46].
Circular RNAs (CircRNAs) are single-stranded, covalently closed RNA molecules that are ubiquitous across species ranging from viruses to mammals [
47]. One of the most commonly reported mechanisms of action of circRNAs is the “sponging” of miRNAs by binding to the miRNA response element (MRE), thereby indirectly increasing the transcription of their target mRNAs [
48].
MiRNAs play an important role in biological processes such as cellular proliferation, differentiation and apoptosis. Further, they are implicated in inflammatory diseases, cancer [
49], immune response, neural development, DNA repair and oxidative stress response [
50].
In addition, miRNAs are stable in body fluids, so they are more available for assays and less invasive than biopsies [
51]. Thus, the interest in miRNAs as both biomarkers and possible therapeutic targets is increasing [
52].
2. miRNA in AD, ACD and CSU: Pathogenetic Role and Therapeutic Strategies
Increasing evidence suggests that miRNAs may play important roles in regulating physiological skin processions, such as self-renewal and differentiation, in stem cells. The most highly expressed miRNAs in normal-condition skin are miR-152, miR-143, miR-126, miR -21, miR-27a, miR-214, miR-16, miR-203, miR 125b, miR-34a, miR-205, miR-27b, miR-30b, miR-125a, miR-191, the miR-200 family (-200a, -200b, -200c, -141, -429), the miR-199 family (-199a, -199b) and the miR-19/-20 family (-19b, -20, -17-5p, -93) [
53]. Skin inflammation is a common underlying feature of AD, ACD and CSU.
2.1. Pro-Inflammatory and Anti-Inflammatory miRNA in AD
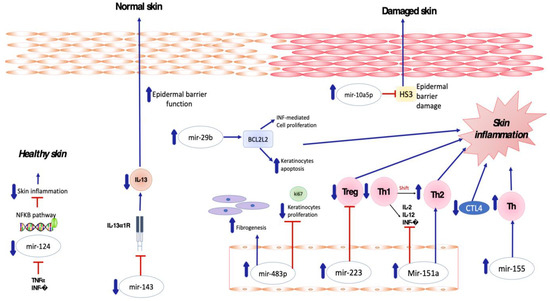
Several miRNAs have been found to be implicated in the crosstalk between inflammatory cells and keratinocytes in patients affected by atopic dermatitis (
Figure 1). In particular, Sonkoly et al. [
54] reported that mir-155 is one of the most significantly upregulated miRNAs in the lesional skin of patients affected by AD, suggesting its major role in the pathogenesis of the disease. MiR-155 is a multifunctional miRNA that is involved in immune cell maturation and the regulation of innate and adaptive immune systems, playing a critical role in both T- and B-cell responses [
55,
56,
57]. Overexpression of miR-155 decreases CTLA-4 levels and increases the proliferation of T-helper cells, promoting chronic skin inflammation. Rebane et al. [
58] demonstrated upregulation of miR-146 in the serum and skin of patients with AD compared to a healthy population, as well as in murine skin specimens obtained from an AD-like mice model in comparison to a control group.
Figure 1. Skewed skin inflammation associated with dysregulated miRNA expression in patients with Atopic Dermatitis.
MiR146a is a known anti-inflammatory and NFκB pathway-dependent miRNA expressed in B cells, T cells, monocytes and dendritic cells [
59,
60,
61]. Its role is to regulate innate and adaptive immunity, regulating antibody production, inflammatory factor secretion and immune cell differentiation [
62].
MiR-151a was also found to be overexpressed in the plasma of 500 atopic dermatitis patients by Chen XF et al. [
63], whereas its expression in eczematous skin was not determined to be relevant. Upregulation of miR-151a favors the shift toward Th2 cell response, being predominant in AD [
64] and suppressing the expression of Th1 cytokines including IL-2, IL-12 and INF-gamma [
65].
MiRNA-143 has been shown to decrease IL-13 activity and inflammation by targeting and downregulating IL-13Rα1 in epidermal keratinocytes, thus playing a potential role in decreasing AD-induced skin inflammation. In detail, miR-143 opposes the negative regulation of filaggrin, loricrin and involucrin induced by IL-13 in human keratinocytes, potentially improving epidermal barrier function [
75].
MiR-124 has been demonstrated to directly target nuclear factor (NF)-κB in B-cell lymphoma [
76]. Yang et al. [
77] investigated the role of miR-124 in atopic dermatitis, demonstrating that miR-124 expression was downregulated in chronic AD skin lesions. Tumor necrosis factor (TNF)-α and IFN-γ were also able to inhibit MiR-124 expression. Modulating the NF-κB pathway, miR-124 reduces chronic skin inflammation and inflammatory responses in keratinocytes in AD [
77].
2.2. Pro-Inflammatory and Anti-Inflammatory miRNA in ACD
Concerning the involvement of miRNAs in the pathogenesis of ACD, Vennegaard et al. were the first to describe aberrant miRNA expression in this disease, analyzing skin biopsies from subjects who had been sensitized with diphenylcyclopropenone (DPCP). They found that miRNA-21, miR-223, miR-142-3p and miR-142-5p were upregulated in the inflammatory lesions. In addition, they were also upregulated in skin biopsies from mouse models which were sensitized with 2,4-dinitrofluorobenzene, showing that mouse models are valuable tools for further study of the involvement of miRNAs in ACD [
78]. Subsequently, in 2015, Gulati et al. [
79] confirmed the altered expression of miR-21, miR-223, miR-142-3p and miR-142-5p in skin lesions obtained 3 and 14 days after a challenge with DPCP in sensitized subjects. They also identified 6 miRNAs that were significantly upregulated 120 days after the challenge, demonstrating the long-lasting allergen-mediated immune reaction that occurs in the skin [
79]. Anderson et al. demonstrated consistent changes in miRNA expression for miR-21, miR-22, miR-27b, miR-31, miR-126, miR-155, miR-210 and miR-301a during the sensitization phase of an allergic response to TDI in a murine model [
80]. In 2021, Werner et al. analyzed miRNA expression data of positive patch test reactions from patients exposed to allergens such as nickel sulphate, epoxy resin and methylochloroisothiazolinone, and to irritants such as sodium lauryl sulfate (SLS) and nonanoic acid. All allergens induced miRNAs expression in human skin, while of the irritants, only SLS did. Eighty-six miRNAs were significantly upregulated or downregulated; miR-142-3p, miR-142-5p, miR-146b and miR-155-5p were differentially expressed across all investigated allergens. Meanwhile, miR-497-5p was significantly expressed only in MCI; miR-22b-3p, miR-99a-5p, miR-193b-3p and miR-199a-3p were significantly expressed in epoxy resin ACD [
81].
2.3. Pro-Inflammatory and Anti-Inflammatory miRNA in CSU
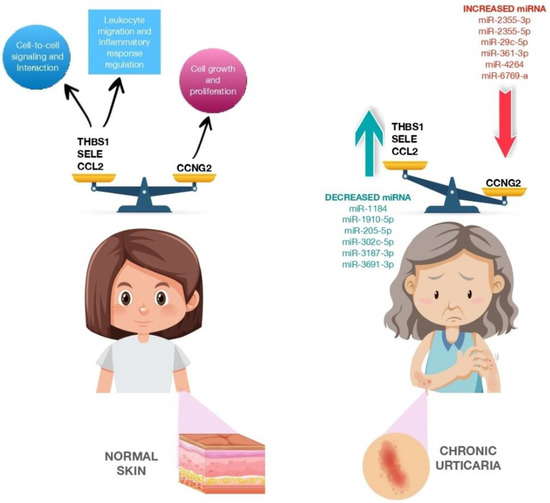
A few studies have examined the role of several miRNAs in the pathophysiology of CSU. In 2017, Lin et al. studied whether miRNAs are involved in CSU regulation as biomarkers. Their study involved 12 patients who were divided according to according to whether they had active hives or no hives and to the presence or absence of CSU. MiRNAs were isolated from patient plasma, and 16 miRNAs were found to be differentially expressed in patients with active hives. Among these, miR-2355-3p, miR-4264, miR-2355-5p, miR-29c-5p and miR-361-3p were upregulated in exacerbated CSU patients; thus, they could be useful biomarkers for patients with chronic autoimmune urticaria. Then, the researchers compared these targets against urticaria-related genes. Twenty-five genes were found to match, eight of which were significantly downregulated, while the other eight were significantly upregulated.
In addition, 12 genes (
Figure 2) did not serve a signaling role, whereas the other 13 were involved in regulatory pathways such as the transforming growth factor beta (TGF-β) signaling pathway (nuclear receptor subfamily 3, group C, member 1, glucocorticoid receptor (NR3C31), kit ligand (KITLG), thrombospondin I (THBS1), chemokine (C-C motif) ligand 2 (CCL2)), glucocorticoid receptor signaling pathway (NR3C1, selectin E (SELE), CCL2), p53 signaling pathway (cyclin G2 (CCNG2), THBS1, CCL2), p21-activated kinase pathway (PAK1 interacting protein 1 (PAK1IP1), KITLG, CCL2), phosphoinositide-3 kinase protein kinase B signaling pathway (KITLG, cholinergic receptors muscarinic (CHRM), THBS1) and neuroactive ligand–receptor interaction (NRC31, histamine receptors H1 (HRH1), CHRM), which could play important roles in CSU [
82]. In 2019, Zhang et al. found miR-125a-5p and CCL17 (C-C motif chemokine ligand 17) to be significantly upregulated in the serum of patients with active CSU and decreased in the serum of patients in remission. Furthermore, upregulated expression of miR-125a-5p was observed in refractory CSU, indicating its potential use as a biomarker [
33].
Figure 2. CCNG2, THBS1, SELE and CCL2 expression in normal skin and CCNG2, THBS1, SELE and CCL2 expression modulated by miRNA dysregulation in patients with chronic urticaria. The figure is designed by Freepik.
This entry is adapted from the peer-reviewed paper 10.3390/biomedicines11051266