2. CXCL1 in Breast Cancer
Female breast cancer is the most commonly diagnosed cancer. In 2020, 2.26 million new cases of this cancer were diagnosed, accounting for 11.7% of all cancer diagnoses [
2]. It also caused 685,000 deaths, which accounted for 6.9% of deaths caused by all cancers. Men can also have breast cancer, although it is more than 100 times less common than in women [
20]. For this reason, this subsection will only discuss the role of CXCL1 in female breast cancer. Breast cancer is divided into subtypes based on the presence or loss of the expression of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) [
21,
22,
23,
24,
25]. The following main subtypes of breast cancer exist:
-
Luminal A (ER+PR+HER2−);
-
Luminal B (ER+HER2− + PR− or Ki-67high);
-
Luminal HER2-positive (HER2+ + ER+ or PR+);
-
Non-luminal HER2-positive (ER−PR−HER2+);
-
Triple-negative breast cancer (ER−PR−HER2−).
Triple-negative breast cancer is sometimes referred to as basal-like breast cancer, which is inaccurate because 1/3 of triple-negative breast cancer cases are not basal-like breast cancers [
26]. The median age of breast cancer incidence in women is 61 [
27]. Risk factors include obesity and an unhealthy diet, and factors that reduce the likelihood of the disease are menopause, a large number of children, and prior breastfeeding. Genetic factors are also important risk factors, accounting for 5–10% of breast cancer cases [
24,
27]. These include mutations in the breast cancer type 1 and 2 susceptibility gene (
BRCA1 and
BRCA2); a mutation in one of these genes is associated with a 55–70% probability of developing breast cancer [
27].
The action of sex hormones plays an important role in the development of breast cancer—no less important are cytokines, including CXCL1. A bioinformatics study indicated that
CXCL1 is an important gene in breast cancer processes in young adults [
28]. There are reports that CXCL1 expression in breast tumors is lower than in normal tissue [
29,
30,
31,
32], although some studies show that CXCL1 expression in breast tumors does not differ from healthy tissue [
33]. This indicates that CXCL1 may not play an important role in all cases of breast cancer. Other CXCR2 ligands may play a more important role in breast cancer than CXCL1, for example, CXCL5/epithelial-cell-derived neutrophil activating factor 78 (ENA78) and CXCL8/interleukin-8 (IL-8), whose expression is increased in breast tumors [
33]. That result was obtained in an analysis of all breast tumors without distinguishing their subtypes.
CXCL1 expression is the lowest in luminal breast cancer and at the same time lower than in normal tissue [
31]. In contrast, CXCL1 expression is the highest in basal-like breast cancer and higher than in normal tissue [
34]. CXCL1 expression is also increased relative to normal tissue in mesenchymal-like triple-negative breast cancer and basal-like triple-negative breast cancer [
35]. CXCL1 expression is higher in inflammatory breast cancer than in other breast cancer subtypes [
36]. Inflammatory breast cancer is a rare breast cancer characterized by intense inflammatory responses in the breast. CXCL1 expression in stroma breast cancer is also higher in invasive ductal carcinoma than in ductal carcinoma in situ, which indicates that CXCL1 expression increases with tumor growth [
37].
Another study showed that expression in breast cancer decreases with increasing tumor grade [
31,
32], while another showed that CXCL1 expression does not differ between grade I and III breast cancers [
29]. CXCL1 expression is higher in breast cancer metastasis (a group composed mainly of liver, lung, skin, and other metastases) than in primary tumors and higher than in normal tissue [
29,
36]. Circulating levels of CXCL1 are increased in breast cancer patients compared to healthy subjects [
38].
CXCL1 expression in breast cancer cells is lineage dependent. MCF-7 cells (ER
+PR
+HER2
−) produce very small amounts of CXCL1 and there is no autocrine stimulation of proliferation by this chemokine in these cells [
39,
40]. The highest production of CXCL1 of all breast cancer lines occurs in triple-negative MDA-MB-231 breast cancer cells (ER
−PR
−HER2
−) [
29]. This chemokine is produced in 14 to 330 times higher amounts by triple-negative MDA-MB-231 breast cancer cells than by MCF-7 cells [
29,
40]. On the other hand, high CXCL1 expression does not always occur in triple-negative breast cancer cell lines. An example of this is triple-negative BT-20 cells, which do not secrete CXCL1 in detectable amounts [
29].
The expression of the receptor for CXCL1, i.e., CXCR2, is lower in triple-negative breast cancer compared to other breast cancer subtypes [
41]. CXCR2 expression is also reduced in ER-negative and HER2-negative breast cancers, indicating a decrease in CXCL1 activity in these tumors and in tumor cells that have lost the expression of these receptors.
The frequency of CXCL1 amplification increases with the progression of breast cancer [
42]. It is estimated that 7.5% of primary breast tumors have
CXCL1 gene amplification, while in lymph node and lung metastases, the percentage is around 20% [
42]. Furthermore, analyses conducted using the cBioPortal tool (
http://www.cbioportal.org/ accessed on 30 March 2023) [
43,
44] based on TCGA PanCancer Atlas studies [
45,
46,
47,
48,
49,
50] showed that
CXCL1 gene amplification was observed in 1.96% (21/1072) of breast invasive carcinoma cases. In tumors with
PT53 gene mutation, the percentage of
CXCL1 gene amplification is 2.88% (10/347). Additionally, in breast cancer with
BRCA1 or
BRCA2 gene mutations, the amplification percentage of
CXCL1 gene is 3.85% (2/52). When it comes to subtypes of breast cancer,
CXCL1 gene amplification was observed in 1.20% (6/499) of luminal A breast cancer cases, 3.55% (7/197) of luminal B breast cancer cases, 7.69% (6/78) of HER2-positive breast cancer cases, and 1.17% (2/171) of basal breast cancer cases.
The expression of CXCL1 is dependent on GATA-binding protein 3 (GATA3), as demonstrated by experiments on T47D cells [
51]. Decreased GATA3 expression leads to increased CXCL1 expression, although the exact mechanism of this relationship is not yet fully understood. It is believed to be associated with the formation of a BRCA1-GATA3 complex, which decreases the expression of
CXCL1 by binding to its promoter [
51].
Another factor that increases CXCL1 expression is obesity. BMI is positively correlated with CXCL1 expression levels in breast tumors [
52]. Due to the significant influence of CXCL1 in tumorigenesis, obesity in breast cancer patients is associated with a worse prognosis.
Other factors are also responsible for CXCL1 expression in breast cancer. The high expression of CXCL1 in triple-negative MDA-MB-231 cells is partly due to decreased expression of the nonreceptor protein tyrosine kinase Syk. This kinase inhibits nuclear factor κB (NF-κB) activation and thus downregulates CXCL1 expression in MCF-7 cells and other breast cancer cell lines [
40]. Other proteins also affect CXCL1 expression in breast cancer. One example is the DEK protein, which downregulates CXCL1 expression [
53], and another is SET domain-containing 2 histone lysine methyltransferase (SETD2), a histone methyltransferase that causes histone methylation in the
CXCL1 gene promoter [
54]. In breast tumors, SETD2 expression is downregulated compared to healthy tissue [
55], and thus CXCL1 expression is upregulated [
54]. Another factor that decreases CXCL1 expression in breast cancer cells is either decreased expression or loss of HER2 expression [
56]. Additionally, a mutation in
KRAS leads to the activation of mitogen-activated protein kinase (MAPK) cascades, which then leads to increased CXCL1 expression in breast cancer cells [
57].
CXCL1 expression in breast tumors is also significantly influenced by intercellular signaling and secretory factors. High CXCL1 expression in triple-negative breast cancer may be due to transforming growth factor-α (TGF-α) [
35]. It is an epidermal growth factor receptor (EGFR) ligand. At the same time, TGF-α does not affect the expression of CXCR2 ligands in non-triple-negative breast cancer cells. CXCL1 expression in breast cancer cells is also increased by secretory factors such as prostaglandin E
2 (PGE
2) [
58], interleukin-17 (IL-17) derived from T helper type 17 (Th17) cells [
59,
60], loss of response to transforming growth factor-β (TGF-β) [
61,
62], tumor necrosis factor-α (TNF-α) [
42], leukemia inhibitory factor (LIF) [
63], plasminogen activator inhibitor-1 (PAI-1) [
64], and Notch signaling [
65].
CXCL1 expression in breast cancer cells is downregulated by TGF-β [
61,
66]. In particular, TGF-β prevents the induction of CXCL1 expression by IL-17 [
61]. At the same time, the expression of the receptor for TGF-β, i.e., TβRII, is downregulated in breast tumors [
62], resulting in an increase in CXCL1 expression in the tumor. This is followed by the recruitment of myeloid-derived suppressor cells (MDSC) into the tumor niche—these cells secrete TGF-β and interleukin-6 (IL-6), which results in the differentiation of CD4
+ T cells into Th17 cells that secrete IL-17, a mechanism further increasing CXCL1 expression in breast cancer cells [
59,
60,
61].
The effect of TNF-α on CXCL1 expression is important in resistance to chemotherapy. Anticancer drugs increase TNF-α expression, which results in the activation of the immune system and the destruction of breast cancer cells [
42]. At the same time, TNF-α also increases the expression of CXCL1, which results in the recruitment of MDSC, cells that inhibit the antitumor immune system response—this means that CXCL1 in this situation is responsible for chemoresistance [
42].
CXCL1 expression is also upregulated by LIF secreted by cancer-associated adipocytes [
63]. LIF expression in these cells is increased as a result of signal transducer and activator of transcription 3 (STAT3) activation by CXCL1 from breast cancer cells. Therefore, a positive feedback loop is formed between cancer-associated adipocytes and breast cancer cells; CXCL1 is one of the components of the loop.
CXCL1 expression in breast cancer cells can also be altered by the plasminogen activation regulatory system. A study on mouse 4T1 breast cancer cells showed that urokinase-type plasminogen activator (uPA) decreases, while PAI-1 increases, the expression of CXCR2 ligands [
64]. CXCL1 expression in breast cancer cells is also increased by Notch signaling [
65]. The activation of this pathway is followed by the activation of hairy/enhancer-of-split related with YRPW motif-like (HEYL), a protein that attaches to the
CXCL1 promoter, thus increasing the expression of CXCL1. In this way, the expression of CXCL2/growth-regulated oncogene-β (GRO-β) and CXCL3/growth-regulated oncogene-γ (GRO-γ) also increases in breast cancer cells [
65].
In breast tumors, cancer-associated fibroblast (CAF) may be the main source of CXCL1 [
37,
67,
68,
69]. CXCL1 expression in CAF is increased under the influence of basal-like breast cancer cells [
70], while luminal breast cancer cells do not increase CXCL1 expression in CAF. This may be the reason for the higher CXCL1 expression in basal-like triple-negative breast cancer than in other breast cancer subtypes [
34,
35].
CXCL1 expression in CAF is also associated with a decrease in TGF-β levels in breast tumors. This cytokine reduces CXCL1 expression in CAF [
37,
69]. Decreased levels of TGF-β increase the expression of hepatocyte growth factor (HGF) [
68], a growth factor that, through its receptor c-Met, increases the expression of CXCR2 ligands in CAF, as shown by experiments on mouse cells [
68].
Oncostatin M, which is secreted by tumor-associated neutrophils (TAN) and tumor-associated macrophages (TAM) [
71,
72], is also responsible for the increase in CXCL1 expression in CAF. On the other hand, oncostatin M also increases the expression of other factors in CAF, such as vascular endothelial growth factor (VEGF), CC motif chemokine ligand (CCL)2/monocyte chemoattractant protein 1 (MCP-1), CCL5/regulated on activation, normally T-cell-expressed and -secreted (RANTES), CCL19/EBI1-ligand chemokine (ELC), CCL21/secondary lymphoid tissue chemokine (SLC), and CXCL12/stromal-derived factor-1 (SDF-1) [
72].
TAM are also responsible for CXCL1 production in breast tumors [
34,
73,
74,
75]. The amount of CXCL1 secreted by TAM may be the highest of all cytokines produced by these cells, as shown by studies of TAM isolated from lung metastasis from a mouse model [
34].
Another source of CXCL1 in breast tumors may be adipose-derived mesenchymal stem cells (MSC) [
76], which increase CXCL1 expression under the influence of interleukin-1β (IL-1β) from breast cancer cells [
77]. Importantly, adipose-derived MSC also secrete CXCL8/IL-8. However, these cells are important in breast tumor development only in some models—they support the development of tumors arising from MCF-7 and ZR-75-30 lines but not from MDA-MB-231 lines [
76].
CXCL1 is involved in tumorigenesis in breast cancer. This effect depends on whether a given cell line expresses the CXCL1 receptor, i.e., CXCR2 [
29]. CXCL1 increases the proliferation of breast cancer cells, as has been shown in experiments on triple-negative MDA-MB-231 breast cancer lines, triple-negative HCC-1937 breast cancer cells, HER2-negative MCF-7 cells, and HER2-positive SKBR3 breast cancer cells (ER
−PR
−HER2
+) [
78]. In triple-negative breast cancer cells, there is often an autocrine increase in cell proliferation by CXCL1. This chemokine is produced and secreted by these cells, which then increases their proliferation [
78].
CXCL1 is also important in the function of breast cancer stem cells. These cells express CXCL1 as well as the receptor for this chemokine, i.e., CXCR2 [
79]. CXCR2 expression in breast tumors is mainly found in cancer stem cells [
41], and for this reason, CXCR2 can be considered a marker of breast cancer stem cells. The simultaneous expression of CXCL1 and the receptor for this chemokine, CXCR2, allows breast cancer stem cells to act on themselves in an autocrine manner. This results in the proliferation of these cells, which increases their stemness and causes self-renewal [
41,
79], as shown by experiments on triple-negative MDA-MB-231 breast cancer [
73,
74].
CXCL1 has pro-survival and anti-apoptotic effects on breast cancer cells. CXCL1 increases the expression of anti-apoptotic Bcl-2 family proteins and decreases the expression of pro-apoptotic Bcl-2 family proteins [
80], which is important in chemoresistance and radioresistance.
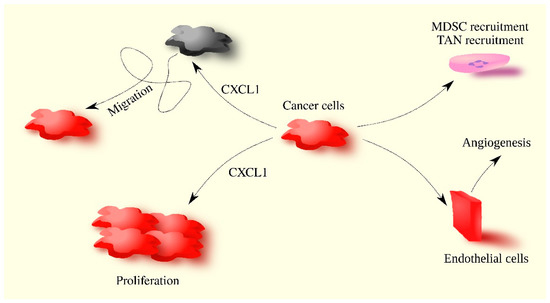
CXCL1 also causes the migration and epithelial-to-mesenchymal transition (EMT) of breast cancer cells, with the effect being lineage-dependent. CXCL1 causes the migration of triple-negative MDA-MB-231 breast cancer cells [
40,
73,
78], MCF-7 cells (ER
+PR
+HER2
−) [
78,
81], Zr-75-1 cells (ER
+PR
+HER2
+) [
81], triple-negative HCC-1937 breast cancer cells, and HER2-positive SKBR3 breast cancer cells (ER
−PR
−HER2
+) [
78]. On the other hand, CXCL1 does not cause the migration of T47D cells (ER
+PR
+HER2
−) [
81], even though T47D cells express CXCR2 [
29]. CXCL1 induces breast cancer cell migration through the activation of extracellular signal-regulated kinase (ERK) MAPK, which increases the expression of matrix metalloproteinase 2 (MMP2) and matrix metalloproteinase 9 (MMP9) [
82]. Also important in the induction of breast cancer cell migration by CXCL1 are Akt/protein kinase B (PKB), NF-κB, and STAT3 [
69].
CXCL1 also induces the EMT of breast cancer cells, as shown by experiments on MDA-MB-231, HCC-1937, SKBR3, and MCF-7 cell lines [
34,
78]. This process is dependent on the CXCL1-induced activation of the NF-κB-sex-determining region Y-related high-mobility group box 4 (SOX4) pathway [
34].
CXCL1-induced migration can be either directly via the activation of the CXCR2 receptor on breast cancer cells or indirectly. Indirectly, CXCL1 causes the recruitment of neutrophils into the tumor niche. TAN cause breast cancer cells to migrate by activating intracellular adhesion molecule-1 (ICAM-1) on these cells [
83]. TAN also show the expression of β
2-integrins, which involves the direct interaction of neutrophils with breast cancer cells and the interaction of β
2-integrins with ICAM-1 on the cancer cell. CXCL1 can also indirectly induce the EMT of breast cancer cells through recruiting MDSC, which then causes the EMT of breast cancer cells via IL-6 [
84].
CXCL1 is also important in breast cancer lymph node metastasis. Breast cancer cells cause an increase in CXCL1 expression in lymphatic endothelial cells (LEC) [
85], which causes breast cancer cells to migrate into lymphatic vessels, resulting in lymph node metastasis.
CXCL1 may also be important in the formation of breast cancer metastasis in other organs. Blood levels of this chemokine are positively correlated with the number of circulating breast cancer cells [
38], and the more circulating cancer cells, the greater the likelihood of metastasis.
CXCL1 has also been found to have a significant effect on the formation of metastasis of breast cancer cells to various organs. CXCL1 may be important in the bone metastasis of the cancer in question. This chemokine acts on osteoclasts precursors through the CXCR2 receptor on these cells [
86,
87,
88,
89]. This leads to osteoclast maturation, bone remodeling, and the formation of bone metastasis. At the same time, CXCL8/IL-8 appears to be more important in the formation of bone metastasis of breast cancer than CXCL1 [
90].
CXCL1 seems to be involved in the formation of brain metastasis of breast cancer. In the formation of brain metastasis, a feedback loop is formed between breast cancer cells and astrocytes [
91]. Breast cancer cells secrete IL-1β, in response to which astrocytes secrete HGF, which in turn increases IL-1β expression in breast cancer cells. IL-1β also increases the expression of CXCL1 and CXCL8/IL-8. These chemokines are responsible for the formation of the perivascular niche and angiogenesis in the initial stages of brain metastasis in breast cancer [
91].
CXCL1 may also play some role in the formation of lung metastasis in breast cancer. The circulating levels of CXCL1 are the highest in patients with lung metastasis in breast cancer and higher than in patients with bone metastasis [
38]. CXCL1 causes an increase in the adhesion of circulating breast cancer cells to human pulmonary microvascular endothelial cells, that is, to the walls of blood vessels in the lungs [
92]. In lung fibroblasts, CXCL1, as well as other CXCR2 ligands, increases the expression of CCL2/MCP-1 and CCL7/MCP-3, both of which increase cholesterol synthesis in breast cancer cells in the lung. This leads to angiogenesis and the generation of lung metastasis in breast cancer [
93]. Notably, this mechanism occurs in triple-negative breast cancer.
CXCL1 acts on tumor-associated cells. CXCL1 in breast tumors causes angiogenesis by acting directly on endothelial cells [
58,
76]. CXCL1 can also indirectly cause angiogenesis by inducing an increase in VEGF expression in breast cancer cells [
78].
CXCL1 is involved in the recruitment of various cells to the tumor niche. It causes the recruitment of neutrophils [
57,
83,
94], granulocytic-myeloid-derived suppressor cells (G-MDSC) [
52,
57], and breast resident adipose tissue-derived MSC [
95]. This process may also be important for the location of the recruited cells in the tumor. Breast cancer cells secrete CXCL1 and CXCL8/IL-8, resulting in the chemotaxis of neutrophils into breast cancer cells [
83]. Direct contact between these cells is then possible, resulting in the stimulation of breast cancer cell migration. This process is dependent on ICAM-1 on breast cancer cells and β
2-integrins on neutrophils [
83].
CXCL1 can recruit c-Kit
+Ly6a/Sca1
+ hematopoietic stem/progenitor cells into the tumor niche, as demonstrated by experiments in mice [
96]. These cells differentiate into MDSC. CXCL1 also induces the recruitment of naive CD4
+ T cells, which are transformed into regulatory T (T
reg) cells under the influence of the tumor microenvironment [
75]. At the same time, CXCL1 can induce the differentiation of naive CD4
+ T cells into T
reg. CXCL1 also acts on cancer-associated adipocytes, causing STAT3 activation in them, which leads to the production of LIF [
63] which in turn increases CXCL1 production in breast cancer cells.
CXCL1 may be involved in chemoresistance and radioresistance. The expression of this chemokine in breast cancer cells is increased by ionizing radiation in microbeam radiotherapy [
97], as well as by chemotherapeutics such as paclitaxel [
98] and doxorubicin [
99]. At least in the case of doxorubicin, this is dependent on a defect in p53 in breast cancer cells and the activation of NF-κB in these cells. The chemotherapeutic-induced increase in CXCL1 expression may be responsible for the increase in TNF-α expression [
42]. CXCL1 also induces the recruitment of MDSC, cells that inhibit an anticancer immune response produced by radio- and chemotherapy (
Figure 2). CXCL1 exerts its anti-apoptotic effect by affecting the expression of Bcl-2 family proteins [
80], which inhibits breast cancer cell apoptosis induced by anticancer therapy.
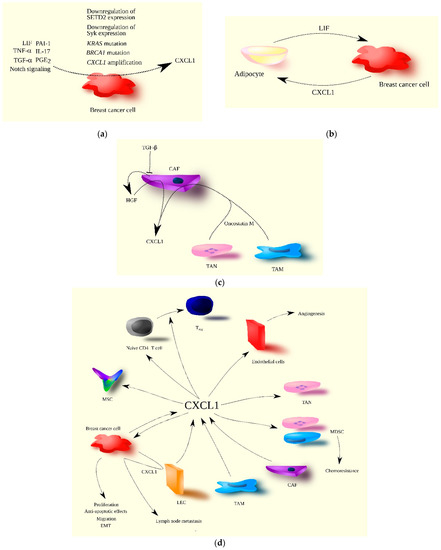
Figure 2. The significance of CXCL1 in breast cancer progression. (a) CXCL1 expression is increased in breast cancer cells by both extracellular and intracellular factors. TNF-α, TGF-α (EGFR ligand), PGE2, IL-17, LIF, PAI-1, and Notch signaling are examples of extracellular factors, while mutations in genes such as KRAS and BRCA1, CXCL1 gene amplification, and the decreased expression of SETD2 and Syk are intracellular factors increasing CXCL1 expression in breast cancer cells. (b) The interaction of breast cancer cells with tumor-associated adipocytes results in increased CXCL1 expression in breast cancer cells due to the secretion of LIF by adipocytes. In turn, CXCL1 increases LIF expression in adipocytes, creating a positive feedback loop between these two types of cells in the breast tumor. (c) In the breast tumor, CXCL1 is secreted by CAF, and its expression in CAF is increased by oncostatin M produced by TAM and TAN. Increased CXCL1 expression in CAF is also due to their loss of responsiveness to TGF-β, resulting in increased HGF production by CAF, which further increases CXCL1 expression. (d) CXCL1 plays multiple roles in breast cancer progression, acting on various types of cells. Source of CXCL1 in breast tumor may be breast cancer cells, CAF, TAM, and LEC. CXCL1 promotes breast cancer cell proliferation, migration, EMT, and anti-apoptotic effects. Breast cancer cells also increase CXCL1 expression in LEC, promoting the migration of breast cancer cells to lymphatic vessels, resulting in lymph node metastasis. CXCL1 also acts on tumor-associated cells. It recruits immune cells such as naïve CD4+ T cells, TAN, MDSC, and MSC to the tumor niche. It can differentiate naïve CD4+ T cells into Treg and facilitate chemoresistance through the recruitment of MDSC. CXCL1 also promotes angiogenesis via its influence on endothelial cells.
High CXCL1 expression in breast tumors is positively correlated with lymph node metastasis [
34,
100] and TNM stage [
34]. On the other hand, there is a study showing that stromal CXCL1 expression is not correlated with lymph node metastasis [
37]. The level of CXCL1 expression is also not related to tumor size [
34,
37,
100] or histologic grade [
34]. The level of CXCL1 in the tumor is not related to the age of patients [
37]. Its highest expression is found in basal-like breast cancer [
34] and triple-negative breast cancer [
35].
Depending on the literature source, the correlation of CXCL1 expression with negative PR and ER status may vary. CXCL1 can be correlated with negative PR [
100] and negative ER status [
82,
94,
100], and in some studies, it is not correlated [
34,
37]. Circulating levels of CXCL1 are not associated with PR and ER status in breast tumors [
38]. Additionally, CXCL1 expression is not associated with HER2-negative status in breast tumors [
34,
37,
100].
Considering all breast cancer cases, the association between the level of CXCL1 expression in the tumor and prognosis varies depending on the literature source (
Table 1). High CXCL1 expression in breast tumors may be associated with either a worse [
34,
37,
100] or better prognosis [
30,
32]. The level of CXCL1 in the blood may be associated with the prognosis of breast cancer patients, with a higher circulating level of CXCL1 indicating a worse prognosis [
38]. Conversely, higher CXCL1 expression is associated with a worse prognosis for patients with basal breast cancer [
52]. For ERα-positive breast cancer, depending on the work cited, high CXCL1 expression in the tumor may be associated with either a worse [
29] or better prognosis [
31].
Table 1. Effect of CXCL1 expression level on survival of breast cancer patients.
|
Type of Cancer
|
Expression Testing Method
|
Impact on Survival at High CXCL1 Expression
|
Number of Patients in the Study
|
Notes
|
Source
|
|
Breast cancer: ERα-positive breast cancer
|
qRT-PCR
|
Worse prognosis
|
48
|
RFS
|
[29]
|
|
Breast cancer: ERα-positive breast cancer
|
Microarray
Kaplan–Meier Plotter database
|
Better prognosis
|
2061
|
RFS
Analysis based on the Kaplan–Meier Plotter database
|
[31]
|
|
Breast cancer
|
IHC
|
Worse prognosis
|
655
|
OS, nuclear CXCL1 expression,
at cytoplasmic CXCL1 expression there was only a trend (p = 0.08)
|
[100]
|
|
Breast cancer
|
Microarray
UALCAN/TCGA database
|
Better prognosis
|
1066
|
OS,
Analysis based on UALCAN
|
[32]
|
|
Breast cancer
|
Microarray
|
Worse prognosis
|
121
|
OS
|
[34]
|
|
Breast cancer
|
Microarray
Kaplan–Meier Plotter database
|
Better prognosis
|
3951/RFS
1402/OS
|
OS, RFS
analysis based on the Kaplan–Meier Plotter database
|
[30]
|
|
Breast cancer
|
Microarray
Kaplan–Meier Plotter database
|
No significant impact on prognosis
|
1402
|
OS,
trend of worse prognosis at high CXCL1 (p = 0.064),
based on the Kaplan–Meier Plotter database
|
[31]
|
|
Breast cancer
|
ELISA
|
Worse prognosis
|
61
|
OS, PFS
circulating level of CXCL1
|
[38]
|
|
Breast cancer: basal breast cancer
|
Microarray
Kaplan–Meier Plotter database
|
Worse prognosis
|
54
|
OS,
based on the Kaplan–Meier Plotter database
|
[52]
|
|
Breast cancer
|
Microarray
Finak microarray database
|
Worse prognosis
|
53
|
RFS
stromal CXCL1 expression
based on Finak microarray database
|
[37]
|
ELISA—enzyme-linked immunosorbent assay; IHC—immunohistochemistry; OS—overall survival; PFS—progression-free survival; RFS—relapse-free survival; qRT-PCR—quantitative real-time polymerase chain reaction. Red color—worse prognosis, Blue color—better prognosis.