Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Chickpea (Cicer arietinum L.), one of the most cultivated legumes worldwide, is crucial for the economy of several countries and a valuable source of nutrients. Yields may be severely affected by Ascochyta blight, a disease caused by the fungus Ascochyta rabiei.
- Ascochyta rabiei
- Cicer arietinum
- Ascochyta blight
- chickpea defense
1. Causal Agent
A. rabiei, the causal agent of Ascochyta blight, can attack all parts of the plant and lead to necrosis, tissue collapse, and therefore to the death of organs or even the whole plant. This may translate into total or major yield losses [15]. A. rabiei (Pass.) Labr. is the name of the anamorph of the fungus during its imperfect or asexual stage. Alternative names for the anamorph are Phyllosticta rabiei (Pass.) or Phoma rabiei (Pass.). It also has a sexual reproductive stage or teleomorph, known as Didymella rabiei (Kovachevski) v. Arx (also Mycosphaerella rabiei Kovachevski) (Figure 1). The key taxonomic descriptors for this fungus are phylum Ascomycota, class Dothideomycetes, order Pleosporales, family Didymellaceae [25,29,30].
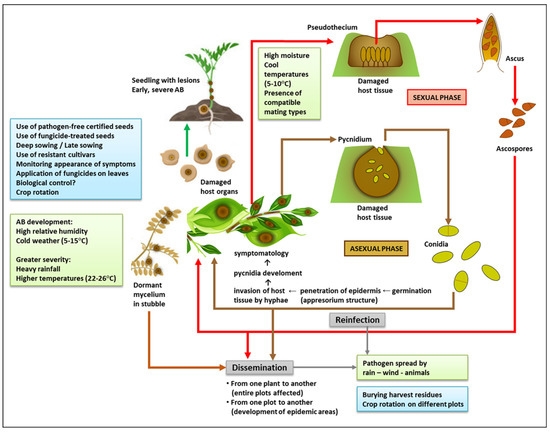
Figure 1. Integrated view of Ascochyta blight (AB) disease: life cycle, predisposing environmental conditions (green boxes), and management practices (light blue boxes). To make this figure, some images were downloaded from https://www.vectorstock.com/royalty-free-vector/green-pod-chickpea-as-annual-legume-plant-vector-31018799 and https://www.vectorstock.com/royalty-free-vector/chickpea-plant-growth-stages-infographic-elements-vector-25494313 (accessed on 5 January 2023).
When the weather is cool and wet, the pseudothecium (the sexual fruiting body) is formed within infected plant tissues. It is a dark brown/black globular structure, akin to a loculated perithecium, which measures approximately 120–270 μm. It contains several pedicellate, curved, cylindrical-clavate asci or sac-like structures (about 48–70 × 9–13.7 μm in size). Within each ascus, there are eight small septate ascospores (12.5–19.0 × 6.7–7.6 μm), each made up of two cells of different size. The bigger cell is prominently formed on the septum itself [31,32]. Since pseudothecia remain inside the tissue after it has died (i.e., inside stubble), ascospores are easily spread to other plots by the wind during the spring and summer [31,32]. The teleomorph is not found in areas where warm conditions prevail after the summer and into the chickpea growing season.
2. Environmental Conditions That Favor the Fungus
The prevalence and spread of the disease are critically influenced by factors such as relative humidity, temperature, and wind [33,34]. Pycnidia, the asexual fruiting bodies of the fungus, can survive more than two years in crop residues if the temperature allows it (10–35 °C with high relative humidity), and the disease typically develops in humid, cold weather (5–15 °C) [35,36]. Temperatures ranging from 22 °C to 26 °C accompanied by heavy rainfall can also favor the appearance of the disease at all stages of the crop cycle (from seedling to pod) [37] (Figure 1). The pathogen is transmitted through stubble and seeds that become infected when it rains in windy weather, through leaves, and by insects and other animals. During the crop cycle, the wind transfers it from infected to uninfected plants, which leads to the formation of spotted areas that may progressively cover entire plots [14]. This progression may be limited in dry weather, but occurs rapidly in humid conditions. Fruiting bodies grow fast at 20 °C [33]. Long periods of cold and humidity are the most propitious for oozing conidia that are spread from pycnidia by the rain [31]. The persistence of such environmental conditions and the presence of compatible mating types [25] favor the sexual cycle, which makes it more likely for the disease to turn into an epidemic. The dissemination distance of the different spores is crucial to determine the spread of the disease. Asexual spores (conidia) disseminate across short distances, and they depend mainly on windy weather and rain splash to spot or infect plots. On the other hand, sexual spores (ascospores) spread through the air, which means they can probably travel greater distances and create epidemic areas of Ascochyta blight disease [38]. These differences are worth considering as part of management and control strategies.
3. Pathogenesis and Symptomatology
A. rabiei is a necrotrophic fungus, and its mycelia can remain dormant in stubble for up to 3–4 years. The chickpea seed is crucial for the continuity of its biological cycle from one generation to the next, since the fungus survives for over five months on the episperm, the cotyledons, and the embryo. Infected grains are therefore the safest and most efficient means for the pathogen to spread and persist [39].
Chickpea is usually affected by A. rabiei during flowering and pod formation [40]. The active fungus can directly target all plant tissues (leaf, petiole, stem, pods, and seeds). Pathogenesis relies on conidia being deposited on the plant surface, their subsequent germination, and the formation of an appressorium-like structure at the tip of the germinal tube. This structure, which penetrates the epidermis, makes it possible for hyphae to invade the adjacent subepidermal tissue and for pycnidia to develop [41,42]. Pycnidia are produced inside the leaves, stem petioles and pods, and even the seeds. They are dark brown globose structures, 140–200 mm in diameter, and have a prominent ostiole [14]. The process may be more or less virulent depending on the aggressiveness of the fungal strain and the tolerance of the infected cultivar [43,44].
The initial symptoms are small necrotic spots. On leaves and pods, necrotic spots are observed as black concentric circles that form round or oval lesions (between 2 and 14 mm in size). On stems, these spots are more oblong (2–30 mm) [26]. Necrosis occurs when the host tissue is degraded by three potent phytotoxins (solanapyrone A, B, and C). Certain enzymes also degrade the plant cell wall (cutinase, pectinase, polygalacturonase, xylanase), while others favor host colonization by acquiring nutrients through the digestion of plant matter (peptidases, lipases), and by inactivating host defenses (dehydrogenases, peroxidases, oxidoreductases) [45]. The production of these virulence factors is related to A. rabiei’s necrotrophic lifestyle, and the levels produced depend directly on the strain’s aggressiveness [45,46]. When infection in the leaves is severe, the entire plant dries up and collapses. High temperatures prevent the disease from progressing to that point, but the tissues remain infected and discrete lesions are still observable [47].
If a pod is infected when it has only just started forming, it becomes unviable for seeds to grow within it [48,49,50]. Once grain formation has begun, the pathogen can easily penetrate the pod wall and settle inside the seed, which significantly enhances its chances of survival and dissemination [51]. Whether superficially or internally infected, seeds may either show no symptoms (if the infection is mild) or shrivel and display dark lesions of different shapes and sizes [52]. As in the case of other plant parts, these lesions are created by brown/black pycnidia growing concentrically (95–220 μm). When hydrated, these pycnidia create a viscous mass that releases conidia [23]. Sowing infected seeds can lead to the establishment of disease at an early stage of plant growth, if the weather conditions are favorable for conidial germination. The emerging seedlings will have dark brown lesions at the base of the stem [53]. Spots formed on the stem at this time are the most serious, since they prevent sap from circulating and thus lead to early death [9] (Figure 1).
4. Plant Defense Mechanisms
Finding chickpea varieties that may be resistant to Ascochyta blight is complicated by a series of factors: the low resistance coded by the primary gene set; the complex genetic basis for resistance conferred by several quantitative trait loci (QTL); the variability of pathogen populations; and the emergence of new pathotypes due to natural recombinations that take place during A. rabiei’s cycle of sexual reproduction [13,38]. Nevertheless, chickpea is known to have an intricate defense response against the fungus. This response varies depending on the crop’s resistance/susceptibility genotype, the fungal strain infecting it, and the environmental factors at the time of infection. Most studies on this topic have focused on the differences in gene expression from one cultivar to another upon exposure to the pathogen, which influences pathogenesis after initial infection [54,55].
Enzymes are among the protective factors that have been identified so far. These include several pathogenesis-related (PR) proteins, such as one acting as a β-1,3-glucanase [56,57], and chitinase, which mediates the degradation of the fungal cell wall [58]. Others, such as polyphenyloxidase, catalase [59], and copper amine oxidase (CuAO), are produced in the epidermis and xylem vessels [60] and are involved in the oxidative burst. This is a process through which the superoxide dismutase copper chaperone precursor (SDCC) and glutathione S-transferase (GST) are downregulated to increase H2O2 levels [53]. Both the oxidative burst and the accumulation of reactive oxygen species (ROS) make up a complex defense mechanism which is related to the hypersensitive response (HR), a way for the plant to prevent microbial propagation through programmed cell death. In resistant cultivars, HR has been detected in association with the synthesis of metabolic enzymes, such as those involved in the production of phytoalexin, an antimicrobial compound [43]. However, certain A. rabiei pathovars have been observed to degrade phytoalexins synthesized by chickpea [61]. Other proteins are overexpressed in plants as part of the defense response against A. rabiei, such as an environmental stress-inducible protein (ESP), a Ca-binding protein, and several others which remain unidentified [53].
Defensive barriers play their part in plant defense as well. Cell walls, for instance, tend to become more stable when exposed to stressful conditions. Infection with A. rabiei induces the expression of the gene that encodes snakin-2 (SN2), a cysteine-rich peptide with a broad antimicrobial spectrum [53], and the production of proline-rich proteins (PRPs) that strengthen the structure of the primary cell wall [62]. This structure is reinforced further by ROS synthesis and the creation of disulphide bridges [63].
Host plants also defend themselves against A. rabiei through the complex upregulation of genes linked to transcription factors. These genes encode products such as (i) a disease-resistance response protein, DRG49-C [53]; (ii) a leucine-zipper protein (LZP), probably involved in the synthesis of PR proteins and the production of salicylic acid [53,64]; (iii) the polymorphic antigen membrane protein PAMP, associated with the transcription of defense genes related to the metabolism of polyamines and nicotianamines [54]; (iv) the ethylene receptor gene ERG, named CaETR1 in C. arietinum L. and located in QTLAR1 [65]; (v) a pathogenesis-related transcription factor, TF1082, linked to the response of ethylene during infection [66]; (vi) the CARNAC transcription factor associated with plant development and defense [67]; and (vii) a resistance gene analog, RGA4, which belongs to chickpea RGA families and has nucleotide binding sites and leucine-rich repeat domains (NBS-LRR) [13,68].
Overall, and in spite of the severity of Ascochyta blight, chickpea can deploy several defense strategies against it, probably in a coordinated manner. The complexity of the response depends on multiple biotic and abiotic factors, and a better understanding of this complexity relies on acquiring further knowledge about the genes involved. The ultimate aim, and perhaps the most difficult to achieve, is to harness these mechanisms for the design of chickpea varieties that will be resistant to blight pathovars under different environmental conditions.
This entry is adapted from the peer-reviewed paper 10.3390/jof9020203
This entry is offline, you can click here to edit this entry!