1. Impact of the Microbiome on Autophagy in Relation to Anticancer Immunity and Self-Tolerance
Microbiome imbalances can lead not only to inflammatory diseases, but also to the growth of tumors (e.g.,
Helicobacter pylori infection and gastric cancer,
Candida infection and oral tumors) [
99,
100,
101]. In addition to the composition of the microbiome, its activity also influences carcinogenesis. Microbial compounds (e.g., N-nitroso compounds, short-chain fatty acids) can have both direct pro- and anti-tumor effects [
102,
103]. Microbiome metabolites also influence immune function and, thus, indirectly affect antitumor immunity. Oral
Candida infection may exert tumor-inhibiting effects by promoting the proliferation of MDSCs [
101]. These findings suggest that the microbiome may influence the efficacy of antitumor therapies. This has also been observed, for example, in probiotics containing
Akkermansia muciniphila or
Bifidobacterium species during immune checkpoint inhibitor (anti-PD-1) treatments and was found to have beneficial effects [
104,
105,
106].
In some immune-mediated diseases (e.g., CD), the interaction of microbial flora, environmental factors, and the immune systems in genetically pre-disposed individuals underlies the development of the disease [
107,
108]. In CD, the composition of the gut microbiome is low in diversity, with some strains (e.g.,
Firmicutes) being present in low numbers [
109]. TLR4 and IL10 receptor mutations have also been linked to
Mycobacterium avium subspecies
paratuberculosis [
110].
The NOD2/CARD15 gene and its variants are crucial to the development and progression of CD [
111,
112]. The NOD2 protein encoded by this gene, which has a role in native immunity against bacterial cell walls, is expressed by monocytes, DCs, intestinal epithelial cells, and Paneth cells [
112]. Some variants (e.g., L1007fs, G908R, R702W) and polymorphisms (e.g., P268S, IVS8 + 158) of this gene increase the risk of developing CD [
110]. In T1D, the structural similarity (i.e., molecular mimicry) among self and microbial antigens (e.g.,
Coxsackievirus,
Rotavirus) has also been associated with the loss of self-tolerance to pancreatic β-cells [
113,
114,
115].
The complex molecular link among insulin resistance, diabetes, obesity, genomic instability and cancer has been described [
116,
117]. Punicalgin, a hydrolyzable tannin isolated from pomegranate juice, has been found to ameliorate high-fat diet-induced insulin resistance, hepatic glucose and fatty acid metabolism disturbances, and damage to the liver by inhibiting the IKKβ/NF-κB inflammation route, controlling the intestinal microbiota equilibrium, and enhancing hepatic autophagy activity [
118]. These results suggest that manipulation of microbial composition may facilitate the restoration of antitumor immunity or self-tolerance, with important therapeutic implications.
2. Involvement of Autophagy in Escape from Central Tolerance and Evading Anticancer Immunity
Researchers have long acknowledged the importance of genetic variables in illness onset and distinct subtypes [
119]. Diverse alleles of HLA class I and II molecules have been linked to the development of autoimmune disorders (e.g., T1D, celiac disease, MS, RA, etc.) [
120,
121]. However, the mechanism by which HLA polymorphisms lead to autoimmunity is currently unclear. It is thought that HLA molecules bind autoantigens with varying affinity [
122].
Patients suffering from non-muscle-invasive, high-risk bladder cancer have a poor prognosis following routine intravenous BCG therapy. Intracellular BCG infection of cancer cells resulted in a post-transcriptional downregulation of HLA-I membrane expression by inhibiting autophagy flux [
123]. After BCG therapy, those with HLA-I-deficient cancer cells exhibited a myeloid immunosuppressive TME with EMT characteristics and poor outcomes. In contrast, following BCG treatment, individuals with HLA-I-competent cancer cells exhibited CD8+ T cell tumor infiltration, an increase in pro-inflammatory mediators, and immune checkpoint-inhibiting compounds. The latter patients had a much better prognosis. It is hypothesized that HLA-I expression in recurrent urinary bladder malignancies following BCG is not the consequence of immunoediting; instead, it originates from an immune subversion mechanism generated directly by BCG on tumor cells and is predictive of a poor prognosis [
123].
Other gene polymorphisms (e.g., cytokines, PTPN22, chemokine receptors, inhibitory checkpoint genes, or costimulatory molecules) have also been linked to antigen recognition and immune cell activation [
124].
Autoimmunity is characterized by the presence of autoreactive T and B cells that are not eliminated by central tolerance mechanisms [
125]. Mutations in the transcriptional AIRE gene are one of the most investigated negative selection defects for T cells. AIRE is predominantly expressed on thymic medullary epithelial cells and serves as the catalyst for tissue-restricted antigen expression. Self-reactive T cells that respond to these antigens are, therefore, killed by negative selection. In the case of AIRE gene defects, autoreactive T cells are discharged into the bloodstream, leading to the development of several autoimmune diseases [
126,
127]. In humans, mutations in AIRE lead to APS1 [
128]. Sex hormones influence the expression of the AIRE gene, causing gender differences in autoimmune disorders [
129,
130]. Additionally, the AIRE gene has been heavily investigated in relation to reproductive malignancies. In prostate cancer, for instance, AIRE expression contributes to anticancer therapy resistance and enhanced tumor invasion. In AIRE+ prostate tumor cells, IL6 and PGE2 production increases, resulting in the formation of M2-polarized (protumor) TAMs. In addition, the AIRE gene also contributes to prostate tumor progression in a murine graft model [
130].
AIRE is also implicated in the regulation of autophagy in THP-1, a human monocytic leukemia cell line. The overexpression of AIRE in THP-1 cells enhanced the quantity of endogenous LC3-II and LC3-expressing vesicles. In addition, the inhibition of autophagy or AIRE knockdown by small interfering RNA mitigated these actions. Contrarily, the expression of p62/SQSTM1 was unaffected by the respective treatment in THP-1 cells [
131]. These data suggest that alterations in autophagy caused by the AIRE gene are more likely to contribute to the co-occurrence of autoimmunity and carcinogenesis than is currently known.
LAMPs regulate cellular processes such as phagocytosis, autophagy, lipid transport, and aging, among others. It is hypothesized that LAMP2 is a functional link between autophagy and self-peptide MHC-II processing for selection in cTECs, which in turn influences the development of CD4 T cells and the variety of their TCR arsenal. LAMP2 is a well-known autolysosome maturation mediator. cTECs were discovered to express LAMP2 at a high level. In particular, genetic suppression of Lamp2 in thymic stromal cell exclusively inhibited the generation of CD4 T cells undergoing positive selection [
132] without inappropriately directing MHC II-restricted cells to the CD8 lineage. The involvement of LAMP2 in several human cancers (such as CRC, HCC, and adenoid cystic carcinoma, as well as breast, prostate, and lung cancers) has also been described and extensively studied [
133,
134]. The results presented indicate that the effects of LAMP2-associated alterations of autophagy on CD4 T cells influence antitumor immunity and tumorigenesis [
135].
The formation of autoreactive B cells in the periphery is caused by a deficiency in the central tolerance mechanism of B cells. The increased number of autoaggressive B cells is also caused by mutations in PTPN22, BTK, and ADA, as well as impaired rearrangements of the BCR light chain and defects in TLRs [
4,
136]. Inhibition of PTPN22 influences T and B cell antigen-specific responses, dectin-1 activation in DCs, the formation and function of Tregs, macrophage responses controlled by NOD2, TLRs, and NLRP3, and neutrophil adherence and mast cell stimulation in an IgE-dependent way [
137]; these functions make it a target for antitumor therapies. A mis-sense mutation at position 1858 (C3T) is the most investigated single nucleotide polymorphism in the PTPN22 locus, which results in the replacement of an Arg (R) to Trp (W) at position 620. The Lyp620W protein variant (rs2476601) interferes with the negative selection of autoreactive B and T cells in the bone marrow and thymus, respectively. [
138]. This protein has been identified in a number of autoimmune conditions, such as T1D, RA, SLE, Graves’ disease, and MG [
139,
140,
141,
142]. However, the identical variant of PTPN22 improves the anticancer defense and decreases cancer incidence [
137,
143]. Holders of the PTPN22(C1858T) mutation possess a reduced risk of skin cancer, whereas homozygotes treated with the anti-PD-L1 antibody atezolizumab have a higher survival rate. In CRC, the majority of important genes were found to be suppressed during tumor formation; however, PTPN22 expression increased at all stages, suggesting its involvement in cancer-related immunity [
144]. These data support the notion that immune tolerance is necessary to prevent autoimmune disorders; however, reducing the value for T cell activation may enhance tumor inhibition and the effectiveness of anticancer therapy.
3. Endocrine Influence on Autophagy, Autoimmunity and Carcinogenesis
Sex hormones may also influence the occurrence of cancer immune evasion and autoimmunity. Immune cells, innate as well as adaptive, produce estrogen α and β receptors (B cells express these receptors at a higher level compared to T cells, NK cells, and monocytes), which are responsible for the activation of tolerogenic actions [
145]. Estrogens control T cell differentiation into Th2 and Treg cells; boost the secretion of IL4, IL10, and TGFβ; enhance FoxP3, GATA-3, CTLA-4; and PD-1 expression on T cells, and suppress the Tfh response. [
146,
147]. During the maturation phase, estrogens modulate B cell tolerance by activating the estrogen receptor. Estradiol is responsible for reducing the generation of B cells while increasing the marginal zone B cell population in the spleen via an increase in BAFF content [
148]. Estrogens modulate the TME-infiltrating immune cells. Some of the estrogen receptor mutations enhance the number of tumor-infiltrating Tregs and Th cells [
149]. Estrogens have also been shown to affect TAMs by inducing their M2-polarization and, thus, enhancing their suppressive function [
150,
151].
It has been known for a while that ERRα has a profound effect on host biology, including autophagy [
152]. E2 regulates the expression of essential autophagy proteins via various transcription factors, microRNAs, and histone modifications that are downstream of the receptors. The E2-regulated autophagic proteins are implicated in the entire mechanism of autophagy [
152,
153]. Autophagy is the primary source of cholesterol, which is the precursor for the manufacture of estrogen. E2 in the blood induces negative feedback that reduces hormone levels in circulation. E2 induces NOS3 activation and NO production via membrane ERRs. NO stimulates autophagy by inhibiting the expression of mTOR [
153].
4. Immune Checkpoints, Apoptosis and Autophagy
Immune checkpoints are sensors that transmit inhibiting signs to immune cells, thereby promoting tolerance and avoiding autoimmunity [
154]. In mouse models, the absence of CTLA-4, PD-1, BTLA, TIGIT, and VISTA results in massive lymphoproliferation, the development of autoimmune diseases, or fatal multi-organ tissue death [
155,
156,
157,
158,
159,
160]. Polymorphisms in immune checkpoint genes have been linked to the susceptibility of humans to autoimmune disorders [
161,
162,
163,
164,
165].
CTLA-4 is a fundamental T cell response regulator that is expressed by both Tregs and activated normal T cells. It inhibits antigen presentation and subsequent activation of naïve T lymphocytes by binding competitively to CD80 and CD86 co-stimulatory receptors on APCs [
166]. CTLA-4 is responsible for ligand binding and trans-endocytosis-mediated ligand clearance from APCs. Therefore, APCs are transiently deficient in CD80 and CD86 and, as a result, co-stimulation via CD28 is diminished [
167]. CTLA-4 is essential for avoiding autoreactivity and its absence in humans leads to immunological dysregulation and PID-associated autoimmunity [
168]. In turn, the absence of CTLA-4 causes an increase in CD28 co-stimulation, which results in the development of autoaggressive T cells and Treg dysfunction [
168,
169]. Expression of CTLA-4 on tumor cells represents a dismal prognosis in melanoma and in pancreatic, nasopharyngeal, and breast cancers [
170,
171,
172,
173]. Anti-CTLA-4 antibodies have been found to enhance the anticancer immune response and deplete tumor-infiltrating Tregs by means of antibody-dependent cell-mediated cytotoxicity [
174,
175]. It has been shown that autophagy suppression contributes to CTLA-4 inhibition resistance in melanoma [
176]. CTLA-4 activation suppresses autophagy by limiting LC3B transcription and autophagosome production [
177]. Activation of autophagy can restore the expression and suppressive function of CTLA-4; therefore, it could be exploited for a potential curative combination with CTLA-4 inhibitors [
176,
177].
PD-1 is an additional essential immunological checkpoint for self-tolerance and immune response termination. It promotes apoptosis in activated T lymphocytes when connected to its ligand PD-L1. In experimental models of autoimmunity, blocking PD-1 or PD-L1 has been linked to illness onset and progression [
178,
179]. PD-L1 is expressed in islet cells of the pancreas, endothelial cells, and placenta, where it protects tissues from autoimmune responses [
180]. In T1D, beta cells that produce insulin overexpress PD-L1 in response to an autoimmune onslaught, which corresponds to the degree of CD8+ T cell infiltration [
181,
182]. The interplay of PD-1 and PD-L1 contributes to the development of inducible Tregs. In vitro and in vivo, PD-L1-negative APCs have a diminished capacity to produce Tregs [
183]. Immune checkpoints are critical for the tolerance and, in particular, maintenance of Tregs. These findings are supported by recent reports of autoimmune side effects in cancer patients treated with PD-1/PD-L1 axis antagonists [
184,
185,
186]. In malignancy, effector T cells are continuously exposed to antigen activation in the TME, exhibiting elevated levels of PD-1 for an extended period of time, leading to T cell depletion. This leaves T cells incapable of eliminating tumor cells, resulting in the progression of cancer [
187,
188]. As a result, cancer cells evade the immune system by using PD-L1, subverting immune surveillance processes that act via PD-L1 expression [
189]. The presence of PD-L1 in melanoma cell-derived exosomes further supports a systemic immunosuppressive effect [
190], whereby CD4+ [
191] and CD8+ [
192] T lymphocytes are reduced and cancer cell clearance is impeded.
By boosting mTORC1 signaling and inhibiting mTORC2 signaling, cancer cell-intrinsic PD-L1 can inhibit autophagy. [
177]. Due to food depletion, PD-L1/PD-1 interaction can promote autophagy in adjacent T cells [
193]. In co-cultures of T cells and tumor cells, a Sigma1 inhibitor induces the degradation of PD-L1 via autophagy and suppresses the functional interaction between PD-1 and PD-L1. Thus, Sigma1 modulators may be able to enhance the tumor immune micro-environment by inhibiting PD-L1/PD-1 [
194,
195]. It was discovered that inhibiting the PD-L1/PD-1 axis with anti-PD1 or anti-PD-L1 antibodies induces autophagy in cancer cells; therefore, it is an appealing treatment for tumors when combined with autophagy inhibitors [
196,
197].
Fas is a crucial component of central and peripheral tolerance. FasL binding suppresses excessive immune cell activation on TCR-stimulated lymphocytes (so-called activation-induced cell death) [
198]. B cell apoptosis triggered by Fas/FasL is crucial for germinal center responses [
198]. FasL inhibits the entrance of activated immune cells to immuno-privileged organs (such as the eye, brain, and placenta) when expressed in these tissues [
199]. In humans, mutations in genes associated with the Fas/FasL axis lead to the development of ALPS [
198,
200]. In ALPS, double-negative, terminally differentiated T cells aggregate and display markers of immune exhaustion [
201]. T1D [
202], autoimmune thyroid disease [
203], and multiple sclerosis [
204] have all been linked to an increase in FasL expression. Fas/FasL signaling is characterized by the fact that soluble molecules do not promote apoptosis in contrast to membrane-bound variants [
205]. Increased soluble Fas/FasL levels have been demonstrated in SLE and Sjögren’s syndrome [
206,
207]. In TME, enhanced Fas signaling is primarily caused by MDSC-produced elevated FasL concentrations. This action results in the apoptosis of TILs, which is an important cause of immunotherapy failure [
208]. FasL can trigger the death of immune cells in the TME, which is obviously linked to an unfavorable prognosis [
208]. Nonetheless, the Fas/FasL system’s significance in cancer growth remains contentious. The ultimate outcome may depend on the level of FasL expression in tumor cells and the concomitant neutrophil-mediated inflammation, according to in vitro studies [
209,
210]. The Bax/Bcl-2 ratio correlates with the susceptibility of tumors to Fas-mediated apoptosis [
211,
212,
213].
Besides the Fas/FasL system, growing evidence reveals connections between other critical proteins of autophagy and apoptosis, which constitute the fundamental mechanisms underpinning their interplay. Through regulating apoptosis, Bcl-2 family proteins play a crucial role in the etiology of cancer [
214]. Bcl-2 transgenic mutations under a B cell-specific immunoglobulin promoter boosted the survival of B and T cells in mice, resulting in splenomegaly, lymphadenopathy, hypergammaglobulinemia, auto-antibody production, and glomerulonephritis [
215,
216,
217]. The relationship between anti-apoptotic protein Bcl-2 and autophagy protein Beclin-1 is essential for modulating the transition between autophagy and apoptosis. Bcl-2 joins to Beclin-1 and dissociates it from class III PI3K, hence inhibiting the autophagic response [
218].
Many investigations have reported that ATGs can play a significant role in carcinogenesis by influencing a number of oncogenic and tumor suppressive pathways [
219]. In addition to Atg genes, which are involved in the development of autoimmunity in multiple ways, Atg12 has a double role in autophagy and apoptosis, connecting both machineries. Through binding to Bcl-2 via a unique BH3-like motif, unconjugated Atg12 favorably promotes mitochondrial apoptosis [
218].
Caspases, a class of cysteine proteases, initiate and regulate apoptotic cascades. A number of malignancies have altered caspase function, with apoptotic evasion designated as the “hallmark of malignancy” [
220]. The pyroptosis-related caspases (i.e., Caspase-1, 3, 4, 5, and 11) are intimately implicated in the pathomechanism of several autoimmune illnesses, including SLE, Sjögren’s syndrome, RA, polymyositis, and IBD [
221]. Caspase-3 can cleave Beclin-1 to prevent autophagy. Caspase-6 can cleave the autophagy regulators Beclin-1, Atg3, and Atg5. Caspase-9 may increase Atg7-dependent autophagosomal LC3-II production and autophagic function [
218].
In inflammation, DNA damage is caused by mitochondrial or nuclear redox imbalance, which initiates the DNA damage response. The signaling protein p53 is implicated in DNA damage response-mediated senescence induction. Increasingly, data suggests that certain organ dysfunctions in autoimmune illnesses are the outcome of inflammation generated by DNA damage-driven senescence [
222]. The important tumor suppressor gene TP53 is most commonly altered in human cancers. Mutant p53 frequently acquires novel oncogenic capabilities in addition to losing its tumor suppressive function, a phenomenon known as “gain-of-function.” Increasingly, evidence reveals that p53 mutations are strongly related to advanced cancers and a poor prognosis [
223]. In the nucleus, p53 increases the production of pro-apoptotic proteins (e.g., Bax, Bid, PUMA, and Noxa), which initiate the intrinsic apoptotic pathway. In the cytoplasm, p53 increases the expression of the TRAIL and Fas receptors, which initiate the extrinsic apoptotic pathway. p53 is also involved in the control of autophagy. DRAM transcriptional activation promotes autophagy induction and autolysosome formation. DRAM appears to be an integral component of the system that modulates apoptosis and autophagy mediated by p53. Moreover, p53 in the cytoplasm inhibits autophagy by stimulating mTOR signaling. Under conditions of nutritional restriction, p53 inhibits the expression of LC3, which regulates autophagy and safeguards cells against “autophagic burst” [
218].
FLIP is an anti-apoptotic protein that suppresses death receptor-mediated apoptosis. Several diseases, including cancer and autoimmune disorders, have been connected to the dysregulation of FLIP expression [
224]. FLIP inhibits autophagy by competing with LC3 for Atg3 binding and inhibiting LC3 lipidation [
218].
BTLA, TIM-3 and TIGIT are also immunological checkpoints [
225,
226]. BTLA impacts B cells in addition to decreasing the response in activated T cells [
227]. In SLE and MS, B and T cell BTLA expression is diminished [
228,
229,
230]. Decreased BTLA expression on naïve B cells in SLE results in elevated levels of IFNγ and autoantibodies, indicating altered B-cell activation during the disease [
228]. In macrophages, BTLA-mediated autophagy and
Mycobacterium tuberculosis clearance require AKT/mTOR signaling [
231]. The expression of BTLA is also present in a wide spectrum of tumor cells. In BTLA-knock-out cells, the overexpression of BTLA reverses the effects of BTLA depletion or HVEM reduction on cell proliferation and colony formation. Upregulation of BTLA or HVEM, in contrast, suppresses cancer cell growth and colony formation [
232]. TLR4 activation of monocytes is required for the production of CXCR5
-PD-1
-BTLA
-CD69
high tissue-resident IL21+ Tfh cells in the TME of HCC. These Tfh cells stimulate plasma cells, leading to optimal conditions for M2 TAM production and the progress of malignancy [
233].
In individuals with autoimmune hepatitis, the number of TIM-3+ T cells declines as IL17 levels rise, which may suggest that TIM-3 deficiency has a hepatotoxic effect. This has been established in mice experiments [
234]. The inhibition of intracellular TIM-3 in melanoma cells was observed to enhance MAPK-dependent carcinogenesis [
235]. In addition, the MAPK and autophagy pathways collaborate to support the survival of RAS mutant cancer cells [
236].
TIGIT is regarded as a hallmark of thymic Tregs with potent suppressive action and stable lineage [
237]. TIGIT and CD226 compete with one another to bind to CD155 and its inhibitory CD112 in DCs [
237]. The pathogenesis of EAE is influenced by TIGIT–CD226 signaling pathway in T cells. EAE mice lacking CD226 have a beneficial Th17/Treg ratio, increased expression of TIGIT and CTLA-4, enhanced pro-inflammatory cytokine levels, and decreased IL10 production [
160,
238]. In cancer cells, a novel TIGIT ligand was recently found. Through interactions with CD155 and CD226/TIGIT, artesunate-induced ATG5-related autophagy was observed to improve NK92 cytotoxicity against endometrial tumor cells [
239]. It has been demonstrated that Nectin4 binds solely to TIGIT [
240]. The TIGIT–Nectin4 interplay reduces NK function, an important component of the anticancer immune response. In addition, antibodies that inhibit Nectin4 have been reported to enhance in vitro and in vivo tumor death [
240]. Nectin4 is also capable of sustaining autophagy induction [
241].
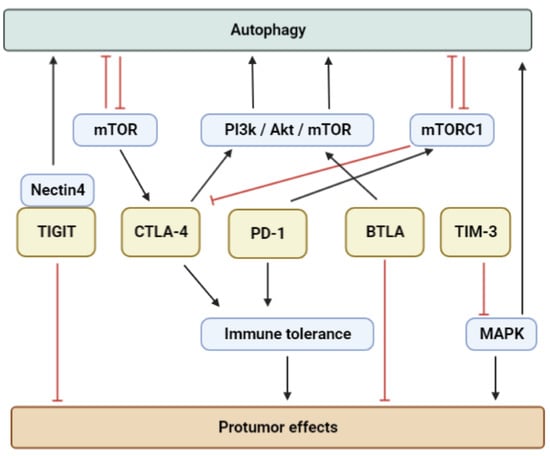
At multiple stages of cancer development, immunological checkpoint molecules and apoptosis are connected to autoimmunity, while autophagy plays a multidimensional role in this intricate interplay (Figure 2).
Figure 2. The intricated molecular correlations between immune checkpoints and autophagy in tumor-promoting immune tolerance are depicted in this diagram. The red arrows represent an inhibitory effect, while the black arrows represent a facilitative impact. The figure was created in part by using
BioRender.com, accessed on the 3 April 2023.
5. Regulatory T and B Cells, Autophagy, Autoimmunity and Carcinogenesis
Thymic Tregs are self-reactive, intermediate-affinity T cell clones that are essential for central tolerance [
242,
243]. In the peripheral blood, Tregs can be potentially generated from naïve or effector T cells. In addition, certain sub-populations of Tregs can be identified based on the cytokines they release (i.e., type 1 Tregs, type 3 Th cells, and IL35-secreting Tregs) [
243]. Functionally, the FoxP3+ population contains Tfr cells that are capable of controlling germinal responses and antibody production [
244,
245,
246].
The primary immunosuppressive mechanisms of Tregs are: (a) strong expression of immune checkpoint inhibitors; (b) tolerance against infections (activation of Tregs by autoantigens results in suppressive activity); (c) secretion of anti-inflammatory cytokines; (d) IL2 deprivation; (e) adenosine accumulation by CD39 and CD73 activity; and (f) immune regulation by extracellular vesicles [
247,
248,
249,
250,
251].
Tregs display a key function in maintaining self-tolerance as their absence results in IPEX syndrome which, without any treatment, leads to multi-organ autoimmune damage [
252,
253]. Many autoimmune disorders (e.g., JIA, RA, systemic sclerosis, and SLE) have been linked to quantitative alterations in Tregs [
254,
255,
256,
257]. It is challenging to explore the involvement of Tregs in human organ-specific disorders since systemic and local immune responses might vary greatly (e.g., autoimmune thyroid disease, relapsing-remitting MS, T1D, and arthritis) [
258,
259,
260,
261,
262,
263,
264]. The transcriptional programs of Th17 and Treg cells are tightly coupled and both are TGFβ-dependent. In response to IL6, Tregs are transformed into Th17 cells, known as IL17+ ex-Tregs. This flexibility decreases suppressive capacity and boosts pro-inflammatory IL-17 and IFNγ production [
265,
266,
267].
Active autophagy in Treg cells promotes their lineage stability and survivability [
268]. Foxp3+ Tregs effectively reduce autoimmunity in vivo via CTLA4-dependent regulation of the autophagic processes in DCs [
269].
Due to the immunosuppressive cytokine environment and chemotactic factors, Tregs are present in a significant proportion of TMEs. The CCR4 or CCR5 chemokine receptor expression on Tregs permits their migration into the TME [
270,
271]. The presence of TGFβ in the TME promotes the development of conventional CD4+ T cells into peripheral Tregs [
272]. Very active and immunosuppressive Treg cells are present in the TME, in part because they overexpress FoxP3 [
273]. Thus, they inhibit antitumor CD8+ T, NK, and NKT cells, as well as M1-polarized macrophages, while promoting DC maturation via IL10, TGFβ, and IDO [
274,
275,
276]. By competitively binding IL2 to conventional T cells and releasing a soluble IL2R component, Tregs remove IL2 and change the actions of cytotoxic T cells. Tregs in the TME augment the expression of PD-1, TIM-3, or LAG-3 via IL35, resulting in the exhaustion of TILs [
277,
278,
279].
The presence of tumor-infiltrating Tregs is associated to a poor prognosis for cancer patients. Substantial intratumoral Treg infiltration suggests that a continuous antitumor T cell response might be finally inhibited [
280,
281,
282]. As autophagy preserves the normal functioning of Tregs by connecting external stimuli and metabolism, the Foxp3CreAtg7
fl/fl mouse model was developed to investigate the intrinsic role of autophagy in Tregs and the impact of Treg-restricted autophagy deficiencies on tumor regulation [
283,
284]. MC38 colon adenocarcinoma cells injected into Foxp3CreAtg7
fl/fl mice significantly suppressed tumor growth. At the tumor site, mice consistently displayed high percentages of tumor-infiltrating CD8+ cells, elevated levels of IFNγ expression in effector CD4+ and CD8+ T cells, and a substantial loss of Tregs [
283]. These results demonstrate the significance of autophagy in the suppression of immune responses against tumors by Tregs.
The B cell maturation process requires checkpoints (i.e., clonal deletion, receptor editing, and anergy) for tolerance to develop [
285,
286]. Breg is a subset of B cells that expresses PD-1, TIM-3, and BTLA; it is essential for the maintenance of peripheral tolerance [
287,
288]. However, no consensus has emerged on the precise definition and phenotype of Bregs [
289,
290,
291,
292,
293,
294,
295,
296,
297,
298,
299,
300,
301]. In experimental murine models of autoimmune conditions, modulation of the Breg compartment by adoptive transfer of separated or ex vivo-induced cells has been investigated (i.e., IL10+ Bregs have been shown to reduce inflammation, especially when used in the early stages of the disease) [
300,
302,
303,
304,
305]. IL10+ B cells and their progenitors are also more frequently found in human autoimmune diseases (e.g., SLE, Sjögren’s syndrome, RA, MS, and autoimmune vesiculobullous skin disease) [
292]; however, their role in pathogenesis is still unknown.
CD25
highCD27
highCD86
highCD1
highIL10
highTGFβ
high human Bregs reduce in a dose-dependent manner the expansion of autologous conventional CD4+ T cells. Moreover, Bregs upregulate the expression of FoxP3 and CTLA-4 in Tregs in a cell-dependent way. When Bregs received prior therapy with TLR9 agonist and CD40L, the effect became even more pronounced [
300]. Other research groups have reported that Bregs inhibit DC as well as macrophage production of cytokines and antigen presentation [
292,
306].
Bregs have a well-defined role in tolerogenesis, producing immunoregulatory cytokines (e.g., IL10 and TGFβ and promoting the contact-dependent repression of autoreactive lymphocytes [
307]. However, the mechanisms of the autophagy/Breg interplay in autoimmunity are not well understood.
B cells, including Bregs, can have pro- and anti-tumor actions [
308,
309,
310,
311]. It is plausible that tumors and TME transform B-cell infiltrators into tumor-induced Bregs [
312,
313]. Bregs that infiltrate tumors destroy the CD3 ζ-chain of CD4+ T cells, hence inhibiting T cell proliferation [
314]. Bregs produced by tumors promote MDSC development [
315] and, in part, contribute to Treg proliferation via IL10 [
294,
300,
316]. TGFβ- or PD-L1-dependent suppression of Th-1 cytokine release and NK cell growth by tumor-induced Bregs in animal models occurs [
313]. Moreover, IL35+ Bregs promote tumor growth by transforming T and B cells into cells with regulatory phenotype [
317].
Tumor cell-released autophagosomes from mouse cancer cell lines were found to stimulate the development of splenic B cells into CD1d + CD5 + IL10+ Bregs, which could potently suppress CD8+ and CD4+ T cell actions in vitro and in vivo [
318].
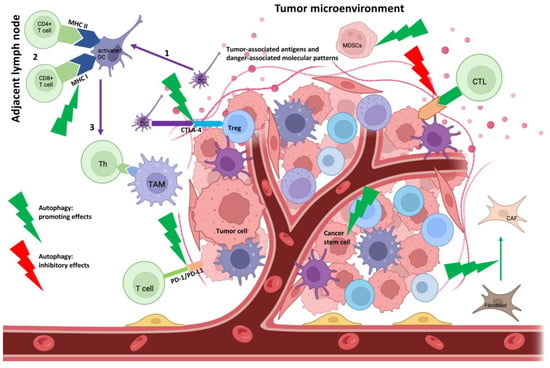
As previously described, the role of regulatory T and B cells in autoimmunity and carcinogenesis is multiple and complex; autophagy is an integral part of these interactions. This may have important therapeutic implications for the future (Figure 3).
Figure 3. Immunobiological impact of autophagy in TME. Tumor-associated antigens formed in tumor cells and DAMPs from dying tumor cells are taken up by DCs (1). During migration and activation of DCs, antigen presentation to naïve CD4+ and CD8+ T cells occurs through MHC and costimulatory molecules (2). Autophagy can enhance the expression of MHC I molecules. Following antigen recognition, T cells differentiate into CTLs and effector Th cells. The differentiated T cells migrate into TME (3), where autophagy may promote increased PD-L1 and CTLA-4 expression and decreased MHC I expression on malignant cells, thereby inhibiting the effectiveness of cytotoxic T cells. In addition, autophagy promotes the formation and survival of MDSCs and CSCs and may enhance the conversion of fibroblasts into CAFs. Together, these effects may be beneficial for tumor development. The figure was in part created by using
BioRender.com, accessed on 4 March 2023.
6. Involvement of γδT Cells in Autophagy, Autoimmunity, and Carcinogenesis
The γδT cells are a CD3+ subset of T lymphocytes that are innate-like and exhibit a TCR composed of γ and δ chains [
319]. Besides the peripheral circulation, they reside in the epithelial layers, constituting the majority of IELs [
320]. They are involved in sustaining homeostasis and modulating the microbiome, as well as exerting potent anti-inflammatory and antitumor effects [
321]. They serve as a link between innate and adaptive immunity, induce rapid immune responses with neutrophils and macrophages against a variety of pathogens, and assist adaptive immune cells in executing their effector functions. In addition, the expression of TCR and NK receptors (e.g., CD94 and NKG2D) enables them to destroy target cells and stimulate additional immune cells [
319,
320].
It is widely recognized that IL17A is essential for the onset and progression of autoimmune conditions [
322,
323,
324]. Innate immune cells, particularly those related to the γδT cell portion, participate in IL17A generation at the initial stage of autoimmune disorders [
325]. The γδT cells can be triggered despite the lack of a corresponding TCR ligand, allowing them to function as potent early inflammatory inducers [
325]. Several molecules (e.g., IL1, IL6, IL23, TGFβ, or TCR agonists) play a role in the differentiation of the Th17 cytokine signature in vitro [
326]. In patients suffering from autoimmune liver conditions (e.g., PBC, PSC, or AIH), there is a significant increase in Vd1+, Vd2+, and Vd3+ γδT cells in the peripheral blood and liver, suggesting the involvement of this particular portion in autoimmunity [
327].
In mice, fine particulate matter (with a diameter of <2.5 µm) has been shown to promote lung inflammation and fibrosis by inducing the production of IL17A in γδT and Th17 cells, hence blocking the PI3K/Akt/mTOR-mediated autophagy machinery in the bronchial epithelium [
328]. MAPK family inactivation was found to block keratinocyte autophagy, which is associated with the worsening of psoriasis in human and mouse models. Krt14Cre/+-hmgb1f/f mice exhibited lessened psoriatic inflammation as a result of the crucial interaction between γδT cells and keratinocyte-specific HMGB1-associated autosecretion [
329]. Further investigation of the role of γδT cells in autophagy and autoimmunity is warranted as they could provide an ideal basis for newly developed cell-based therapies with immunomodulatory effects.
The γδT cells are a component of the TME and are known to influence the antitumor immune response to a wide variety of malignancies [
330]. Targeting immune checkpoints has been shown to restore the malfunctioning condition of γδT cells in the TME. Moreover, immune checkpoint inhibition induces antitumor effects by enhancing the proliferation, activation, and cytotoxic effect of γδT cells [
331,
332]. It has been shown that isolated human γδT cells induce autophagy in myeloma cells by suppressing the expression of autophagy-related phospho-AKT, PI3K, and phospho-mTOR, while increasing the expression of BECN1 and AMPK [
333]. On the other hand, a previous study highlighted the role of IL17A-producing γδTILs in promoting tumor growth in CRC patients; however, these findings were not corroborated by subsequent research [
334,
335].
As can be seen, the rapid activation and cytotoxic nature of γδT cells make them promising candidates for use in cell-based immunotherapies; however, under certain conditions they may induce pro-tumor functions. Investigation of the tumor-influencing role of γδT cells via autophagy is certainly warranted and could form the basis for novel, individualized treatments.
7. Myeloid-Derived Suppressor Cells and Macrophages in Relation to Autophagy, Autoimmunity and Carcinogenesis
MDSCs are found in malignancies as active, immature myeloid cells with immunosuppressive functions. They are classified into two distinct populations: M-MDSCs and PMN-MDSCs [
336]. Many cytokines (e.g., GM-CSF, VEGF, SCF, prostaglandins, TNFα, IFNγ, and IL18) have a role in their differentiation and, hence, the production of immunosuppressive TME and maintenance of CSCs [
337]. MDSCs suppresses the activity of TILs, EMT, and angiogenesis, as well as the development of the pre-metastatic niche [
338,
339]. Additionally, their iNOS activity increases NO generation, which results in T cell death and suppressed proliferation, as well as inhibited antigen presentation in DCs [
340,
341]. They inhibit T cell growth, survival, and TCR signaling via their elevated ROS levels [
342,
343,
344]. By arginase I overexpression, they deplete the TME of critical amino acids, which impairs T cell activation and proliferation [
345,
346]. Moreover, they inhibit T cells by downregulating the CD3 ζ-chain of the TCR complex [
347]. By activating IDO, they inhibit the proliferative capacity and survival of T cells and stimulate Treg activation [
348,
349,
350]. By generating peroxynitrite, these cells nitrate the TCR complex and render cytotoxic T lymphocytes unresponsive to certain antigens presented by MDSCs [
351,
352,
353]. MDSCs are the major source of the immunosuppressive adenosine [
354,
355], hence suppressing T cell activation and effects, predominantly via A2A and A3 adenosine receptors [
356]. MDSC-produced IL10 and TGFβ enhance the T cell-to-Treg transition and inhibit the activation of T and NK cells, as well as DC activity [
357,
358]. Via TGFβ synthesis, they stimulate EMT in cancer cells [
339,
359] and create pro-tumorigenic M2 macrophages and N2 neutrophils [
339,
359,
360,
361].
The persistence of TME cells, such as Tregs and MDSCs, is strictly correlated with HMGB1-induced autophagy and tumor resistance to immune surveillance [
362]. Autophagy specifically modifies the metabolism, longevity, and development of MDSCs, being essential for MDSC regulation. Autophagy displays a significant effect on MDSC modulation by inhibiting antitumor immunity, whereas inhibiting autophagy restricts tumor growth and increases immunity against malignant cells [
363].
The immunosuppressive capacity of MDSCs has also raised attention regarding their function in autoimmune disorders. MDSCs have also been split into two subgroups in immunological studies: M-MDSCs and PMN-MDSCs [
364,
365]. In an EAE animal model, arginase-I-producing MDSCs were identified in the spinal cord, which showed tropism to demyelinated areas of the CNS; their numbers correlated with the number of apoptotic T cells, disease progression, and clinical status, particularly at the end of the active phase [
365,
366]. In humans with relapsing-remitting multiple sclerosis, the number of PMN-MDSCs in the circulation during relapse were considerably higher than during remission or in the control group. Based on in vitro tests, it has been demonstrated that PMN-MDSCs suppress the growth of autologous T cells, suggesting a role in the induction of remission [
367].
Numerous forms of autoimmune disorders are associated with elevated levels of MDSCs, such as T1D, RA, SLE, IBD, and autoimmune hepatitis [
368]. The increasing frequency of MDSCs in individuals with RA and SLE is related to disease progression [
369,
370]. The ratio of Th17 cells to MDSCs was discovered to be inversely proportional in RA [
371]. Th17 cells were initially identified as a cause of some inflammatory disorders. Furthermore, MDSCs have been shown to play a pro-inflammatory function and can trigger arthritis with the help of Th17 cells [
369]. The concept of harnessing the suppressive activity of MDSCs for therapeutic purposes inspired experiments involving MDSC adoptive transfer into diabetes-prone mice; these transfers effectively avoided the development of autoimmune diabetes and induced tolerance to the mice’s own antigens through the induction of Tregs [
372].
Within infection-related inflammatory circumstances, it was shown that inhibiting autophagy contributes to the aggregation and inhibitory effect of PMN-MDSCs by encouraging the activation of STAT3 signals, indicating that autophagy could have a crucial role in controlling the aggregation and function of MDSCs [
373]. Moreover, in tuberculosis, MDSCs were found to express TLR4, mTOR, and IL6 as compared to healthy controls. HMBG1 serum concentration was in parallel to mTOR during treatment [
374]. These findings suggest that MDSCs have a close relationship with the complex process of autophagy in certain infectious inflammations. Since infections can trigger autoimmune reactions, the possibility of MDSC/autophagy interaction in autoimmune diseases cannot be excluded; however, no specific proof has been presented.
Macrophages, a class of highly diverse immune cells, are capable of polarizing to various phenotypes in response to the microenvironment. M1-polarization refers to conventionally activated, pro-inflammatory macrophages, whereas M2-polarization refers to alternatively activated, anti-inflammatory macrophages [
375].
The recruitment of TAMs to the TME is driven by chemokines (e.g., CCL2) in lung and breast cancers, as well as glioblastoma [
376,
377,
378,
379]. In addition, via CCL2 production, TAMs recruit even more macrophages and enhance their M2-polarization [
380,
381]. TAMs are capable of producing IL8, which has been linked to a dismal prognosis regardless of the amount of CD8+ T cells in the tumor [
382]. TAMs can also generate IL6, IL10, and TGFβ. IL6 in conjunction with IL6R can activate anti-apoptotic pathways and extend tumor cell survival [
375]. Additionally, TAMs release inflammatory mediators, such as PGE2 and MMP-7, thereby inhibiting DC and macrophage activation caused by TLR or IFNγ. Moreover, they indirectly impede T cell identification of tumor antigens by directly inducing genes that inhibit APC function [
383].
Despite the fact that macrophage-mediated phagocytosis plays a crucial role in immune surveillance in malignancies, it has been shown that phagocytosis is significantly compromised, a phenomenon that is primarily due to the autophagy machinery in tumor cells [
384,
385]. Moreover, autophagy enhances MHC-II expression in macrophages and MHC-I expression in tumor cells [
385].
A T1D animal model has proven the permanent existence and function of macrophages as APCs in peripheral tissues [
386]. In numerous autoimmune diseases (e.g., MS, RA, SLE, SSc), M1/M2 macrophage subgroup imbalances have been identified [
387,
388]. Recent investigations utilizing multiparametric analysis on the pancreas of T1D patients have shown the existence of macrophages with mixed M1/M2 features, indicating the remarkable adaptability of these cells [
389,
390]. According to studies, the polarization of macrophages in EAE follows the normal trend of the disease [
391]. In a murine model of SLE, adoptive transfer of M2-polarized macrophages ameliorated the disease [
392] and, in NOD mice, delayed the onset of diabetes by targeting the site of insulitis [
393].
SQSTM1/p62-mediated clearance of damaged mitochondria regulates the activation of NLRP3-inflammasome in macrophages; deletion of SQSTM1/p62 generates enhanced inflammasome activation and overproduction of IL-1β [
394]. Notwithstanding the fact that this process inhibits excessive IL1β-dependent inflammation, other reports have demonstrated that autophagy increases NF-kB action in certain tissue macrophages [
395].
MDSCs and macrophages display a crucial role in both autoimmunity and carcinogenesis; the complex process of autophagy is involved at several points in their development, function, and regulation of other immune system elements. This will not only aid the understanding of the pathogenesis of these disorders, but also identify new therapeutic targets in the future (Figure 3).
8. The Cross-Talk of Cell-to-Cell Interactions with Autophagy, Autoimmunity, and Carcinogenesis
Emperipolesis, entosis, and cell cannibalism have been described as cell engulfment phenomena that can even involve tumor cells [
396,
397]. Emperipolesis is the phagocytosis of the host’s intact hematopoietic cells (mainly neutrophils, lymphocytes, and plasma cells) by other, sometimes tumorous cells. Entosis represents a mechanism of homogenous live-cell invasion, such that the invading cell seems to take the initiative in being internalized. Cannibalism is the active internalization and destruction of dead or living tumor cells by other engulfing cells [
396]. In recent years, a growing body of evidence suggests that these cell-to-cell phenomena are linked to autophagy by multiple strands [
398].
Emperipolesis is a characteristic feature of AIH, which is linked to severe necrotizing inflammation and advanced fibrosis [
399]. Emperipolesis is frequently described in conjunction with interface hepatitis, hepatocyte rosettes, and plasma cell infiltration. The vast majority of lymphocytes engulfed by phagocytosis in liver cells are CD8+ T cells and are mainly observed in the area of caspase-3-induced apoptosis [
399]. NK cells are involved in liver fibrosis [
400]. HSCs impair the killing effect of NK cells by TGFβ secretion [
401]. NK cells can initiate mitophagy, a selective form of autophagy, after virus infections, hence maintaining their own survival [
402]. Autophagy is also involved in the transition of effector NK cells to long-live memory cells, partly via an ATG3-dependent mechanism [
402]. Moreover, ATG5 deletion interrupts the development and death of NK cells by causing severe mitochondrial damage and ROS generation [
403]. To accelerate development and maturation, the collaboration between ATG7 and FoxO1 triggers the cytosolic autophagy flux of immature NK cells [
403].
Recent research has shown that overexpression of p62 is linked to lymphocytic emperipolesis and cytoplasmic vacuoles with eosinophilic inclusions in fumarate hydratase-deficient RCC, which is a rare form of renal cancer. The upregulation of p62 is considered to be evidence of defective autophagy [
404] and cannot be excluded as a major contributor to the aggressive nature of this tumor type.
It has been demonstrated that the autophagy process contributes to the demise of engulfed cells. Entotic vacuole membranes surrounding engulfed cells recruit LC3 via autophagy machinery-related proteins (e.g., ATG5, ATG7, and VPS34) [
405,
406]. Internalized cells display some characteristics of autophagic cell death, including an overabundance of autophagosomes and an elevated level of autophagy flux [
405]. It is hypothesized that in liver cells, pro-inflammatory cytokines, such as TNFα, IFNγ, and IL6, may inhibit T cell release following entosis [
407]. This would explain why CD4+ T cells have a tolerogenic mechanism in the liver to prevent further inflammation. Although the majority of pro-inflammatory cytokine treatments did not affect the percentage of T cells released from hepatomas, shorter incubation durations with stimuli resulted in a greater percentage of T cells released from HepG2-CD81s than 24-h incubations. This result indicates that the organization of hepatomas may affect the rate and frequency of entosis and release. It is crucial to acquire a deeper understanding of the causes of entosis and the discharge of CD4+ T cells; this allows T cells to be manipulated therapeutically to prevent the development of inflammatory liver diseases [
407].
It is theorized that entotic cell death serves as a tumor suppressor by promoting the demise of entotic tumor cells [
408]. Nevertheless, entosis can result in polyploidy and aneuploidy, which support the progression of tumors. In addition, multinucleation is frequently observed in incorporating host cells after glucose starvation, mitosis, and matrix deadhesion [
408,
409]. Entosis sometimes does not result in the death of the entotic cell. Consequently, under certain conditions, entotic inner cells retain viability, multiply inside the exterior cell, and eventually leave the outer cell [
408].
The interpretation of entosis as either pro- or anti-tumorigenic is not entirely evident at this time; however, the majority of authors believe that it confers a survival advantage to the engulfing cell [
410]. The majority of published clinico-histopathological studies of entosis link its presence to a more aggressive phenotype and a poorer prognosis. High numbers of entosis were found to be positively associated with improved survival for anal cancer and certain subtypes of breast cancer, whereas low numbers had a favorable prognostic value for rectal and bronchial malignancies, HNSCC, and PDAC [
410]. Investigating the relationship between entosis and autophagy could lead to the development of new anticancer therapeutic targets.
Immature T cells internalized by TNCs in an MHC-driven manner differentiate into αβTCR
highCD4 + CD8 + CD69+ cells and subsequently exit TNCs [
411,
412]. TNCs have a negative selection role in addition to a positive selection function that promotes TCR transformation. Indeed, they select anergic thymocytes for removal at an early stage of intracellular death processes. Recently, it has been shown that concomitant expression of K8- and K5-cytokeratins, P63, TRA, and AIRE in TNCs contribute to negative selection through the expression of autoantigens [
413].
In the case of nutrient depletion, mTORC1 activity decreases, which induces autophagy and macropinocytosis and supports the metabolism of tumor cells [
414]. The mTORC1-regulated ULK kinase complex that promotes autophagy is implicated in this process [
415]. TM9SF4 positively modulates autophagy and cannibalism and contributes to inhibiting mTORC1 activity in response to nutrient deprivation [
416]. Elucidating the association between cannibalism and autophagy may help to improve understanding of the pathomechanism of immune-mediated diseases and cancer, as well as the development of new types of targeted therapies.
This entry is adapted from the peer-reviewed paper 10.3390/biomedicines11041130