Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Environmental Sciences
石油烃是相对顽固的化合物,作为污染物,它们是最严重的环境问题之一。正烷烃是石油烃的重要成分。合成生物学和代谢工程策略的进步使正烷烃生物降解在解决环境污染问题方面更具设计和可操作性。
- biodegradation
- n-alkanes
- alkane hydroxylases
1. 生物降解途径
正构烷烃生物降解主要有1条途径,包括末端氧化途径、亚末端氧化途径、双端氧化途径和Finnerty途径。这里以十六烷为例来解释四种降解途径(图<>)。
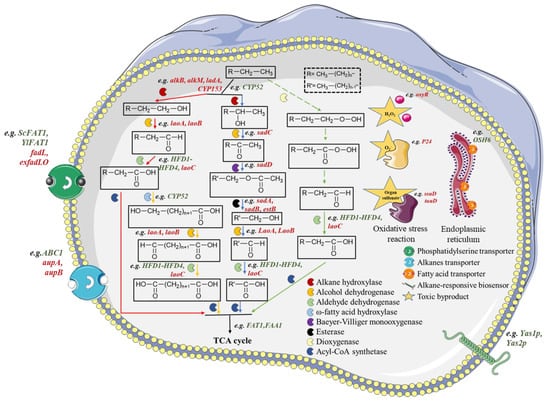
图1.烷烃生物降解的四种途径。末端氧化、亚末端氧化、二端氧化和芬纳蒂方式分别用红色、蓝色、橙色和绿色表示。红色斜体表示原核基因,绿色斜体表示真核基因。虚线表示推定的路径。
末端氧化途径是最常见的降解途径,可见于许多细菌中,如白藜芦SK2(T)[15]、恶臭假单胞菌KT2440[16]和地芽孢杆菌热反硝酸芽孢杆菌NG80-2[17]。以十六烷为例,十六烷被烷烃羟化酶(如AlkB、AlkM、LadA和细胞色素P1家族)氧化成450-十六烷醇,然后1-十六烷醇被醇脱氢酶进一步氧化成1-十六醛[18]。最后,它被醛脱氢酶转化为十六烷酸。作为一种脂肪酸,十六烷酸最终进入β氧化。Forney和Markovitz[19]在铜绿假单胞菌中发现了亚末端氧化。5年后,该途径在戈多尼亚属菌株TY-20中得到认可[2]。与末端氧化途径不同,十六烷被烷烃羟化酶(例如AlkB和细胞色素P450家族)氧化为2-十六烷醇。然后,2-十六烷醇被氧化成2-十六醛。在Baeyer-Villiger单加氧酶(BVMO)的催化下,21-十六醛可以转化为乙酸十四烷基酯。乙酸十四烷基酯通过酯酶进一步氧化成相应的醇,后续步骤与终末途径相同。最后,脂肪酸进入β氧化。二端氧化途径首先在细菌中发现。在几种酵母菌株中也发现了这种途径[16]。该途径的特殊点是ω-羟化酶的存在,它可以将十六烷酸转化为更有价值的16-羟基十六烷酸。1-羟基十六烷酸被醇脱氢酶和醛脱氢酶进一步氧化。产物十六烷二酸进入β氧化。芬纳蒂的方式与上述三种方式不同。该途径由Finnerty假设,并在不动杆菌属菌株HO22-N中发现[1]。第一步,十六烷被双加氧酶催化,转化为<>-氢过氧十六烷和十六烷过氧酸。然后,十六烷过氧酸进一步氧化成十六烷酸,进入β氧化。
2. 相关基因
在微生物中,烷烃降解涉及许多降解途径基因,并且可以受到许多其他过程的影响,例如氧化应激反应和识别以及运输过程。因此,在构建和优化烷烃降解剂时,有必要考虑所涉及的所有基因。随着合成生物学和代谢工程策略的发展,许多与烷烃降解相关的基因已被鉴定和分析。一些重要的基因如下(图1)。
在烷烃降解过程中,编码烷烃羟化酶的基因起着重要作用。一些基因,如alkB [23],alkM [24],ladA [25]和编码CYP153C1蛋白的基因[26]已在原核生物中鉴定出来。同时,rubA和rubB [27]被识别为编码原核生物中的烷烃羟化酶辅酶,而位于alkM旁边的alkR具有相反的方向,编码多肽的转录调节因子。在真核生物的烷烃降解中,CYP52基因家族频繁出现,尤其是在酵母中[28,29]。例如,在蓍草中分离出编码细胞色素P1ALK的12个基因(ALK450至ALK30)[52]。此外,属于CYP21家族的CYP52A23和CYP52A31基因编码烷烃/脂肪酸羟化酶。它不仅可以氧化白色念珠菌中的烷烃,还可以氧化ω-羟基化脂肪酸[32]。在醇的降解中,laoA和laoB分别编码醇脱氢酶及其辅酶来降解伯长链醇;然后,laoC编码醛脱氢酶,将长链醛转化为相应的酸[1]。此外,从脂解酵母中鉴定出的HFD4-HFD33在烷烃降解途径中也可以执行与laoC相同的功能[34]。 在二级醇降解基因簇上,sadC参与二级醇的降解;sadD、sadA和sadB编码BVMO,两种酯酶催化仲醇转化为相应的伯醇[1]。最后,由FAT1和FAA35编码的酰辅酶A合成酶催化脂肪酸进入TCA循环[<>]。
烷烃的识别和转运以及氧化应激反应是影响烷烃降解的两个重要方面。烷烃的识别和运输通常发生在细胞膜上。分别编码外膜蛋白(AupA)和内膜蛋白(AupB)的基因aupA和aupB负责原核生物中烷烃的摄取[36]。编码ABC1转运蛋白的基因ABC1参与真核生物中烷烃的转运[37]。一些编码脂肪酸转运蛋白的基因,如真核生物中的ScFAT1和YlFAT1[38]以及原核生物中的exfadLO [39]和fadL[40],可能通过增加或减少脂肪酸积累间接影响烷烃降解。此外,编码烷烃反应性生物传感器的1个基因Yas2p、Yas3p和Yas41p被人工转移到酿酒酵母[<>]。 本研究对酿酒酵母可降解烷烃的构建具有启发意义。在烷烃降解过程中,一些有害物质如H2O2, O2−,并可能产生有机磺酸盐对细胞造成氧化损伤。同时,已经确定了相关基因。oxyR编码毒力相关的氧化还原感应转录因子,赋予对H的抵抗力2O2 [42]P24基因被认为编码超氧化物歧化酶,可以防止因O而受损2− [ssuD和tauD可以将有机磺酸盐转化为硫源,这是产生一些抗氧化应激感应蛋白和代谢物所必需的,这些蛋白质和代谢物可以抵抗氧化应激[43]。 值得一提的是,由OSH44基因编码的Osh6p是氧化甾醇结合蛋白的同系物。据推测,它通过改变磷脂酰丝氨酸的比例,然后影响内质网膜环境,参与Alk蛋白的形成[6]。
This entry is adapted from the peer-reviewed paper 10.3390/bioengineering10030347
This entry is offline, you can click here to edit this entry!