Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemistry & Molecular Biology
Melanin is a biological pigment formed by indoles and phenolic compounds. It is widely found in living organisms and has a variety of unique properties. Due to its diverse characteristics and good biocompatibility, melanin has become the focus in the fields of biomedicine, agriculture, the food industry, etc.
- melanin
- structure
- synthesis
- decomposition
- bioactivity
- application
1. Introduction
Melanin originates from the Greek word “melanos”, which means black or very dark, reflecting the characteristics of melanin appearance; this term was first applied by the Swedish chemist Berzelius in 1840 to call a dark pigment extracted from eye membranes [1]. Melanin has a very long history as an ancient pigment that has been produced naturally since the beginning of life on Earth. Melanin can be found in well-preserved dinosaur fossils, prehistoric bird feathers, plants, marine cephalopods, bacteria, and fungi [2]. As early as 1840, “melanin” was used to refer to the black pigment in animals. Currently, melanin is commonly used to refer to a range of bio-pigments associated with most organisms, broadly defined as “heterogeneous polymers” formed by the polymerization of intermediate phenols and quinones from the oxidation of phenols or indole compounds [3].
Melanin can be found widely in nature, such as in the skin and hair of mammals, the ink of cephalopods, plants, and different types of bacteria and fungi [4]. It has a wide variety of functions in the biosystem. In animals, melanin is produced by melanocytes present in the epidermis and hair follicles and plays a role in sun protection and camouflage [5]. In plants, melanin acts as a reinforcer of cell walls and cuticles, increasing the resistance of plants to microbial and viral infections [6]. In addition, melanin protects microorganisms from environmental stresses, such as high sunlight exposure, low temperature, low water content, starvation, elevated reactive oxygen species, and increased radioactivity [7]. There are mainly three types of melanin, which are named eumelanin, pheomelanin, and allomelanin, respectively. Eumelanin is found in animals, microorganisms, and some fungi. It is derived from tyrosine and is black or brown in color [8]. Pheomelanin is endemic to higher animals, mammals, or birds. It is also a tyrosine derivative and is red or yellow in color. Pheomelanin consists of sulfur-containing monomer units, mainly benzothiazine, and benzothiazole, rather than the indole units in eumelanin [9]. Nitrogen-deficient plant and fungal melanin is often called allomelanin. Their precursors are different. Fungal melanin can be formed from γ-glutaminyl-3,4-dihydroxybezene, catechol, and 1, 8-dihydroxy naphthalene, while catechol, caffeic acid, chlorogenic acid, protocatechuic acid, and gallic acid have been considered as possible precursors in plants [3,10].
Melanin is a polymer formed by 5,6-dihydroxyindole (DHI) and its 5,6-dihydroxyindole 2-carboxylic acid (DHICA). However, its macromolecular structure is not very clear. Recent studies have demonstrated that the macromolecular structure of melanin has at least four layers and three particle types, which belong to supramolecular structures [11]. Moreover, the proportion of DHI and DHICA in the oligomer plays a very important role in the subsequent polymerization process. When the DHI ratio is large enough, the oligomers will have a π-stacking effect. Conversely, random bundling is performed due to its nonplanar structure [12]. Depending on the ratio and coupling site of DHI and DHICA, it can form large heteropolymers or laminated depressors. However, both structural forms of melanin have the same physiochemical properties [13]. Melanin is amorphous and predominantly dark brown to black, with red and yellow sometimes being observed [14]. Melanin is known to be resistant to concentrated acids, light, and reducing agents but is soluble in bases and phenols [15].
2. Classification and Origin of Melanin
Melanin is a heterogeneous polymer of phenolic or indoles, which is amorphous and has a wide range of functions. The colors of melanin are mainly from dark brown to black; sometimes, reddish and yellowish are also observed [3,42]. The widely accepted classification of melanin into eumelanin, pheomelanin, and allomelanin is based on the chemical composition of the pigment monomeric subunit structure. Recently, it has also been reported that melanin is classified into five major categories, namely eumelanin, pheomelanin, allomelanin, neuromelanin, and pyomelanin, respectively (Table 1) [43].
Table 1. The five major melanins and their precursors. Adapted from [43] with minor modifications.
|
Pigment |
Monomer Precursors |
|---|---|
|
Eumelanin |
tyrosine,5,6-dihydroxyindoles |
|
Pheomelanin |
cysteinyldopas, benzothiazines |
|
Neuromelanin |
dopamine, catecholamines |
|
Allomelanin |
1,8-dihydroxynaphthalene, phenolic precursors |
|
Pyomelanin |
homogentisic acid |
3. Synthesis and Decomposition of Melanin in Nature
3.1. Synthesis of Melanin in Humans and Animals
In humans and animals, melanin is produced in specialized cells called melanocytes, mainly at the epidermal-dermal junction, and then to the surrounding keratinocytes, which are the most abundant cells in the epidermis. Melanocytes look like dendritic cells and contain specialized lysosomal lineage organelles called melanosomes, which synthesize and store melanin. Melanosomes are transferred from melanocytes to neighboring keratinocytes by elongated dendrites [83]. The synthesis of melanin in humans follows the Raper-Mason pathway, in which L-tyrosine is synthesized in an environment with or without cysteine through a series of enzymatic reactions catalyzed by tyrosinase, resulting in the production of true melanin or brown melanin, a pathway first proposed by Raper in the 1920s and later demonstrated by Mason using spectrophotometry [84,85]. The biochemical pathway of melanin synthesis begins with the amino acid L-tyrosine, and the key enzyme is tyrosinase. Tyrosinase catalyzes two sequential reactions; one is the hydroxylation of L-tyrosine to L-dopa. L-tyrosine, which is delivered by the gastrointestinal tract (GI) or produced by L-phenylalanine hydroxylase (PH) -mediated hydroxylation of L-phenylalanine, can be hydroxylated to L-dopa by tyrosine hydroxylase (TPH) or tyrosinase (Tyr), depending on the cell type [86]. Tyrosine hydroxylation catalyzed by tyrosinase was first proposed by Aaron Bunsen Lerner [87].
The other is the oxidation of tyrosine by tyrosinase to produce dopaquinone. In the absence of cysteine, dopamine quinone undergoes an intramolecular addition of an amino group to produces cyclic dopa (white dopachrome). Redox exchange between white dopachrome and dopaquinone produces dopachrome and dopa. Dopa is the noncyclic reduction product of dopaquinone, while dopachrome is the cyclic oxidation product of cyclodopa. Dopa is recruited into the pathway by tyrosinase, and dopachrome continues to form eu-/pheo-melanin [88]. Dopachrome slowly and spontaneously decarboxylates to 5, 6-dihydroxyindole-2-carboxylic acid (DHI), and dopa pigment tautomerase (Tyrp2/DCT) can catalyze the formation of 5, 6-dihydroxyindole-2-carboxylic acid (DHICA) from dopachrome (Figure 1). DHI and DHICA are indole-o-diphenols that are readily oxidized to their corresponding o-quinones by tyrosinase or tyrosinase-associated protein 1 (Tyrp1). These species eventually aggregate to form eumelanin [89]. DHI and DHICA can be oxidized to their respective o-quinones by oxygen and ROS produced in the previous reaction without any enzymes. The post-dopa oxidase steps of melanogenesis were first described by John Pawelek [90].
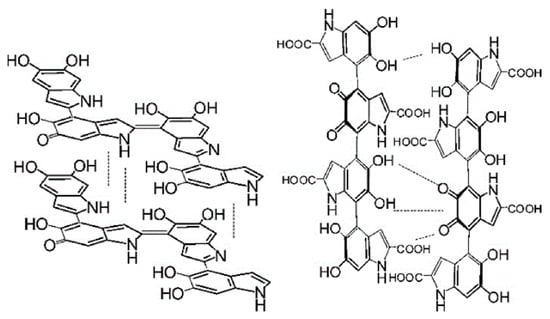
Figure 1. Representative structures of DHI and DHICA melanin. Left: DHI, right: DHICA. Adapted from [75] with minor modifications.
In any case, eumelanin is a polymer with a mixture of decarboxylated (DHI) and carboxylated (DHICA) 5, 6-oxidized indole units (5, 6-dihydroxyl, semiquinone, and 5, 6-quinone units) of varying degrees of oxidation [91]. On the other hand, dopa quinones can bind to thiol-containing compounds such as free L-cysteine or glutathione, thereby branching into the production pathway of sulfur-containing that produce pheomelanin. As a result, human skin contains two types of melanin, eumelanin, and pheomelanin. Eumelanin is dark and varies from black to brown depending on the DHI/DHICA ratio, whereas pheomelanin is a red or yellow pigment and is found mainly in superficial, blonde, or red hair [92,93]. On the other hand, in the presence of cysteine, dopaquinone reacts rapidly with cysteine to produce 5-S-cysteine dopa and, to a lesser extent, 2-S-cysteine dopa. Cysteinyl dopa is then oxidized to generate the benzothiazine intermediate and, finally, pheomelanin [94].
L-dihydroxyphenylalanine (L-dopa) is the product of enzymatic hydroxylation of L-tyrosine, which is decarboxylated to dopamine and further processed to norepinephrine or epinephrine in neurons or chromaffin cells [95]. The non-essential aromatic amino acid, L-tyrosine, in addition to protein synthesis, also acts as a precursor for melanin, catecholamines, tyramine/octopus, and thyroid hormones [96]. L-dopa and L-tyrosine have been recognized not only as continuous substrates and intermediates of melanogenesis but also as positive regulators essential for melanogenesis. In Bomirski hamster melanoma cells, L-tyrosine supplementation not only increased melanin formation but also enhanced tyrosine hydroxylase activity and tyrosinase oxidation activity to dopa. L-dopa exerts a dose-dependent stimulatory effect on tyrosinase activity and melanin deposition by expanding the melanocortin (MSH) receptor system [97,98,99]. L-dopa itself does not affect the melanogenic pathway in melanoma cells, but at micromolar or lower concentrations, it stimulates cell proliferation [100].
L-tyrosine and L-dopa can synergize with UVR to increase MSH receptor expression activity and promote melanin synthesis. L-tyrosine, as well as phosphorylated isoforms of L-dopa, can stimulate cell surface MSH receptor expression, increase the level of MSH stimulation of tyrosinase, and reduces positive cooperativity among cell surface MSH receptors [101,102,103]. Similarly, UVB upregulates a-MSH receptor (MC1R) expression and activity [104], POMC expression and production of POMC peptides, including a-MSH, b-endorphin, and ACTH [105,106], potentially regulating mammalian skin pigmentation, protecting skin from UV-induced damage and modulating skin immune responses [107,108,109].
During melanogenesis, some metal cations may have a significant effect on the rate of formation and the final structure of the synthesized melanin [110]. The degree of influence varies from one metal ion to another, and these differences depend on the nature of the ions, the pathway steps followed, and the presence of dopachrome tautomerases in the reaction mixture. Regarding the first step of the pathway, tyrosine hydroxylation, Co (II), and Ni (II) enhanced this activity, but Zn (II) inhibited it. In the second part of the pathway, the presence of metal ions also increases the rate of conversion of dopachrome to indole, inhibits decarboxylation, and enhances the binding of carboxylated precursors to the final pigment, with a less pronounced effect of Zn (II). Ni (II) and Co (II) stimulated melanin formation from L-tyrosine and L-dopa catalyzed by tyrosinase, but the inhibition of tyrosine hydroxylation by Zn (II) inhibits melanin formation. The activation induced by Ni (II) and Co (II) may originate from the direct reduction of copper at the tyrosinase active site. The differential inhibition of this reaction by Zn (II) does not exclude the inactivating effect of this ion on melanoma tyrosinase [111]. In the second phase, the combination of cations (except Zn (II)) with dopamine tautomerases inhibits decarboxylation, and the combined effect of cations and enzymes always inhibits decarboxylation more than each individually [110,112].
In addition to metal ions, hormones also play an important role in the process of melanogenesis. Locally produced melanocortin (MSH) and adrenocorticotropic hormone (ACTH) can regulate melanogenesis by paracrine, autogenous, or endocrine mechanisms [113]. In humans, MSH or ACTH stimulates hyperpigmentation of sun-exposed skin [114]. Chronic synthetic ACTH leads to skin atrophy, hyperpigmentation, and hirsutism (see references [113,115], and high serum MSH concentrations are also associated with skin hyperpigmentation. Structurally, the MSH peptide and ACTH share the same amino acid sequence -Tyr-X-Met-X-His-Phe-Arg-Trp- containing the tetrapeptide -His-Phe-Arg-Trp- essential for melanic activity. Polymorphisms in MC1 receptors are associated with skin and hair hyperpigmentation. Mutations that reduce MC1 receptor activity leads to lighter skin pigmentation and redness; the external environment, such as ultraviolet radiation or the high level of MSH or ACTH in the plasma, can combine with the over-expressed MC1 receptor in melanocytes, leading to skin or hair hyperpigmentation [108,116,117].
It should be noted that melanogenesis is a metabolic pathway specific to normal and malignant melanocytes that can influence the behavior of their surrounding cells or environment. Melanosomes can alter cellular energy production metabolism by converting oxidative catabolism to anaerobic glycolysis, altering intracellular NAD/NADH and NADP/NADPH ratios, and stimulating the pentose phosphate pathway [118,119,120]. Melanogenesis, however, produces an oxidative environment in which some of its intermediates, quinones, and semiquinones, are directly toxic but also mutagenic. Therefore, the mutational environment associated with melanogenesis may contribute to genetic instability. In addition, since melanogenic intermediates can suppress the activity of immune cells, an immunosuppressive environment may surround the tumor. Finally, the final product, melanin, can scavenger free radicals and reactive oxygen species (ROS), resulting in a relatively hypoxic environment due to increased oxygen consumption [121,122].
3.2. Synthesis of Melanin in Plants
Melanin has been shown to be widespread in the plant kingdom, and the biochemistry and molecular genetics of melanin formation in plants is less well-studied compared to animals and microorganisms. In addition to the complex polymeric nature of pigments, one reason is that plant melanin accumulates in the hard seed envelope, where other compounds with similar colors may be present, such as proanthocyanidins [10]. Some plants have been shown to accumulate melanin in their seeds by physicochemical methods, such as Castanea mollissima, Avena sativa, Helianthus annuus, Citrullus lanatus, Hordeum vulgare, Fagopyrum esculentum, Vitis vinifera, and Ipomoea purpurea [123]. Melanin synthesis in plants and polyphenol oxidase (PPO) in the damaged tissues of enzymatic browning reaction, polyphenol oxidase (PPO) can be used in the presence of low oxygen to phenolic copper redox enzyme family due to aging, damage, and the interaction of pests and pathogens and in the process of post-harvest processing and storage, processing. Loss of compartment integrity in cells results in the release of PPO from the plastids in which they reside into the cytoplasm. PPO contacts vacuolar phenolic substrates and forms highly reactive o-quinones. O-quinone subsequently polymerizes nonenzymatically or interacts with other compounds, such as thiols, amino acids, and peptides, then it forms colored products. O-quinone can also interact slowly with water to form triphenols or reduce to primitive phenols [124,125,126].
3.3. Synthesis of Melanin in Microorganisms
Several bacteria have been reported to produce different melanin genomes by specialized pathways or by exploiting enzymatic imbalances in modified metabolic channels [78]. The regulation of the melanin synthesis mechanism in bacteria includes transcriptional regulation and metabolic regulation. Melanin biosynthesis is present in Gram-positive, and Gram-negative bacteria, such as Streptomyces grays, Bacillus licheniformis, and Rastonia solanacearum [127,128,129]. Fungi have been reported to have all types of melanin. Melanin in fungi is considered to be a secondary metabolite that can be synthesized from endogenous substrates via 1,8-dihydroxynaphthalene (DHN) intermediates or L-3, 4-dihydroxyphenylalanine (L-dopa) [130]. Although melanin is not essential for the growth and development of fungi, it can perform a wide range of biological work. For example, it is essential for host invasion of plant pathogens. Melanin in the cell walls provides mechanical strength to the appressorium and aids in tissue penetration [131].
Typically, most microbial melanin is formed by the conversion of tyrosine (DOPA pathway) or malonyl-CoA (DHN pathway), facilitated by different enzyme genomes. The dopa pathway is very similar to mammalian melanin synthesis. The melanin precursor tyrosine is converted to levodopa, which is then converted to dopaquinone by tyrosinase and laccase. Dopaquinone has high activity and can spontaneously oxidize and self-polymerize to form melanin. Synthesis of melanin through the dopa pathway is called dopa melanin or eumelanin [27]. The precursor of the DHN pathway is propylene glycol—coenzyme a. Polyketo compound synthase catalyzes the continuous decarboxylation of five malonyl-CoA molecules to produce 1,3,6, 8-tetrahydroxy naphthalene (THN), which through a series of reduction and dehydration reactions generates 1, 8-dihydroxy naphthalene (DHN), and finally polymerizes into DHN melanin. Notably, both pathways can be found in bacteria and fungi. Most bacteria and basidiomycete fungi synthesize melanin through the dopa pathway. However, ascomycetes and some bacteria, including non-microscopic fungi, such as truffles, use the DHN pathway to produce melanin [17,79,130].
3.4. Decomposition of Melanin
The metabolic process of melanin mainly includes the synthesis, transport, and degradation of pigment. Currently, most of the research is limited to the synthesis and transport of melanin, especially in the synthesis; however, studies of melanin degradation are rarely involved, and the mechanism of its occurrence is still unclear [132,133].
Melanocytes are cells found in the skin tissue that produce melanin. Melanosomes are large organelles responsible for the synthesis, storage, and transport of melanin. Mature melanosomes are transferred from melanocytes to the nucleus of keratin-forming cells to form hyperpigmentation [134]. Melanocytes use long dendrites to contact many keratinocytes (one epidermal melanocyte provides pigment to more than 40 keratinocytes) and transfer melanosomes from the site of formation in the center of the cell to the main site of transfer at the tip of the dendrite. Melanocytes accomplish this task by combining the long-range, bidirectional microtubule-dependent transport of melanosomes along the length of the dendrite with the actin-based motor protein myosin capture and local movement of organelles in the actin-rich dendritic tips. Finally, melanosomes accumulated at the tip of dendrites by this cooperative trapping mechanism are transferred out of melanocytes and into keratinocytes. This helps distribute pigment in the hair and skin [135,136]. In addition to being associated with melanogenesis, melanocytes also act as sensing and regulatory cells in the human epidermis. For example, melanocytes can act as intra-epithelial stress sensors, alter KC functions by the transfer of melanosomes, regulate immunity, serve as neuroendocrine cells similar to APUD cells for amine-precursor-uptake and decarboxylation, amplify and convert signals collected from adjacent cells into chemical messages to maintain cellular homeostasis [137].
Lysosomal enzymes are involved in the degradation of melanin in keratogens. Lysosomes are cytoplasmic vesicles surrounded by a membrane monolayer, widely distributed in all mammalian cells except erythrocytes, and play an important role in cell proteolysis. Currently, lysosomes have been shown to contain more than 60 kinds of hydrolases, which can degrade and digest proteins, nucleic acids, and polysaccharides [138].
After treating radiolabeled melanosomes with lysosomal hydrolases, Japanese scientists noted that lysosomes isolated from hepatocytes did not appear to degrade melanin by themselves but that lysosomal enzymes hydrolyze the core protein fraction in melanosomes. The melanosomes are mainly composed of an outer membrane and an inner membrane matrix, and the main components are probably proteins on which the melanogenesis of polymerized indole 5, 6-quinones, and other tyrosine intermediates take place [147].
The basic molecular unit of eumelanin is thought to be a small planar oligomer composed of several 5, 6-dihydroxyindole-2-and 5, 6-dihydroxyindole-2-carboxylic acid units, which are further assembled into a stable high-order parallel layer structure by P-stacking and lateral interactions [148]. Therefore, considering its structure, a redox mechanism similar to polycyclic hydrocarbon conversion may be involved in the degradation of melanin [149]. This idea is further supported by several reports. Melanin can be degraded by oxidizing agents such as potassium, permanganate [150], and hydrogen peroxide [151], and the degradation process is accompanied by mild bleaching [21] and a strong fluorescence phenomenon [152,153,154,155]. In addition, potassium permanganate and hydrogen peroxide oxidation are also the basic methods used to quantify melanin [156].
4. Macromolecular Structure and Physicochemical Properties of Melanin
4.1. Macromolecular Structure of Melanin
The structure of melanin is closely related to its properties and functions. Therefore, it is very important to elucidate the macromolecular structure of melanin to get a better understanding of its properties. It is generally believed that melanin is a heteropolymer formed by DHI and DHICA in a certain ratio (Figure 1). The ratio of these two components depends on the type and synthesis pathway of eumelanin, which ultimately leads to changes in the properties, such as appearance, oxidation resistance, metal chelation performance, etc. [157]. The content of DHICA of eumelanin prepared by the enzymatic method is only 10%, while natural eumelanin could reach up to 50% [158].
Eumelanin synthesis involves the enzymatic oxidation of tyrosine or dopa to give dopachrome, which undergoes isomerization to 5,6-dihydroxyindole-2-carboxylic acid (DHICA) with tyrosinase-related protein (Tyrp2). In the absence of Tyrp2, dopachrome spontaneously decarboxylated into 5,6-dihydroxyindole (DHI) [159]. DHICA melanin exhibits more potent hydroxyl radical-scavenging properties than DHI. Moreover, DHI melanin is composed mainly of planar oligomeric scaffolds, while DHICA is composed of distorted linear oligomeric structures characterized by resistive isomerism caused by slow rotation around interunit bonds [160].
Polydopamine (PDA) is a supramolecular aggregate of monomers containing three main types of structural units, namely, uncyclized amine-containing units, cyclized eumelanin-type indole, and pyrrole carboxylic acid units, respectively [161]. Therefore, PDA consisted of a mixture of oligomers in which indole units and open-chain dopamine units with different levels of unsaturation produced charge-transfer interactions between o-quinone and catechol units [162].
Basic eumelanin particles are approximately 15 A in size, consisting of four to five layered stacks with a stack spacing of approximately 3.45 A, and are composed of four to eight molecules composed of 5, 6-dihydroxyindole monomers in a plane [163]. However, graphite-like nanostructures, about 15 nm in size and consisting of dozens of stacked carbon layers, are found to exist in carbonized PDA nanoparticles by using high-resolution TEM and Raman spectroscopy [164].
4.2. Physicochemical Properties of Melanin
The solubility of melanin varies according to the source, purity, and polymerization state. Most melanin is virtually insoluble in distilled water and most organic and inorganic solvents and will precipitate under acidic conditions [165]. However, it is soluble in dimethyl sulfoxide (DMSO) and alkaline water (pH 10.0) [2]. The solubility of melanin highly depends on the pH of the solution. The decrease in the pH value of melanin solution will promote the formation of aggregation and precipitation [43], while the increase in the pH value will lead to the decomposition of particles into small particles with a poor degree of polymerization, which is called oligomers. The presence of intramolecular ionizable units and hydrophobic interactions is responsible for this behavior [166]. The solubility of melanin is also related to the ionization of carboxyl, phenol, and amine groups in the molecule, polyelectrolyte characteristics, and amino acid content [167].
Melanin has unique reaction properties that can be used for the initial detection and characterization of melanin. The bleaching nature of melanin is one of its characteristics. Pigment decolorization occurs in the presence of potassium permanganate, potassium dichromate, sodium hypochlorite, hydrogen peroxide, or other oxidizing agents, which is related to the degradation of the pigment [43]. The reaction mechanism with H2O2 is nucleophilic attack by OOH−, which induces a ring-opening reaction leading to the formation of quinone epoxides that bleach the melanin. The reactivity of melanin also includes the reaction with the AgNO3 solution. Gray precipitation will appear on the tube wall, which is caused by the reduction of AgNO3 [42].
5. Biological Activity of Melanin
Melanin contains phenolic hydroxyl, carboxyl, and amino active groups, which can absorb UV light, and is a natural endogenous functional substance. The biological activities of melanin are closely related to its composition and structure. Among the properties of melanin, the representative ones are free radical scavenging, antibacterial, chelating metal, anti-tumor, and radiation protection activities, which have gradually become the hot spot and trend of melanin activity research.
6. The Application of Melanin
6.1. Applications in Agriculture and Industry
In industry, melanin can be used as an adsorbent for heavy metal pollution control. Heavy metals in various chemical forms and states are highly migratory, enriched, latent, and biotoxic and accumulate when they enter the environment and human body, threatening ecological environment safety and human health [194]. The prevention and control of heavy metal pollution are important to solve environmental problems. Compared with other adsorbents, nanoparticles have a higher reaction rate and adsorption capacity due to their small size, large specific surface area, high surface energy, and chemical activity. As a typical natural, non-toxic nanoparticle [195], melanin has a large specific surface area, and the structure surface has many functional groups and active sites that can bind to heavy metal ions. Secondly, melanin itself can be uniformly dispersed but insoluble in water, which facilitates subsequent operations such as solid-liquid separation. This excellent property of melanin can be effectively utilized and applied in various industries.
It has been proved that melanin has an ideal effect as an adsorbent, which can effectively remove Hg, Cr, Pb, Cu, and other heavy metals in a short period of time (3 h). In addition, melanin also shows good adsorption capacity when the concentration of heavy metals in the solution is lower than 10 mg/L. The adsorption capacity even surpasses some commercial adsorbents currently available on the market [185,196]. Melanin can also adsorb some organic drug molecules because of its diverse and heterogeneous structure. Melanin can bind widely used drugs in high concentrations, such as chloroquine, amlodipine, atorvastatin, and telmisartan. The binding amount is positively correlated with the drug concentration [197,198]. In agriculture, melanin can also be used as a pesticide photoprotective agent, prolonging the action of drugs because of the good light absorption effect. Melanin can reduce the influence of light and climate conditions on the biopesticides of Bacillus thuringiensis and is an ideal photo-protectant of biopesticides [199].
6.2. The Application in the Food Industry
Food packaging is an important part of the food industry. Its main role is to protect food from various external factors, such as temperature, ultraviolet light, humidity, oxygen, pressure, microorganisms, etc., in order to maintain the quality of food and extend its shelf life. Melanin has effective antioxidant, anti-radiation, and antibacterial activities, which can be used in food packaging to extend the shelf life of food products. The addition of melanin to the PLA films can improve the antioxidant and antibacterial activities and enhance mechanical strength and air-tightness [200]. The addition of melanin to PVA films can enhance the UV shielding and antioxidant properties [201]. When melanin isolated from squid ink is added to a nanocomposite film based on carrageenan, the thermal stability, UV radiation resistance, and the ability to resist foodborne pathogens are enhanced [202]. Low-density polyethylene films mixed with melanin can significantly enhance thermal stability and UV shielding, as well as water and oil resistance [203]. As discussed, melanin has a promising future in food packaging.
6.3. Biomedical Applications
Melanin can significantly increase the levels of specific and non-specific (including carbon scavenging) immunity, humoral immunity, and cellular immunity [29]. Calf RPE melanin can be involved in the regulation of retinal immune response, as evidenced by a significant increase in IL-6 secretion and expression in retinal epithelial cells [204]. Melanin extracted from the surface of Aspergillus fumigatus conidia combined with surfactant protein D exerts a PAMP effect, stimulates the secretion of proinflammatory cytokines, activates the host immune response, and promotes the spores to be more effectively phagocytic by macrophages [205]. This unique function could have promising applications for the modulatory treatment of immunocompromised patients and the development of new immune-enhancing nutraceuticals or drugs.
Melanin has the anti-tumor activity of promoting apoptosis and inhibiting angiogenesis. It can effectively improve the body weight, liver, spleen, and thymus index of mice with cancer and effectively regulate inflammation-induced high levels of various proteases, creatinine (CRE), blood urea nitrogen (BUN), and uric acid (UA) without causing toxic and side effects on other normal cells or tissues of the body [16]. Due to its good light absorption effect, melanin can absorb light from the ultraviolet region to the near-infrared region and has good photothermal conversion efficiency, which can be used as a photothermal agent for multimodal imaging-guided photothermal therapy (PTT) to selectively destroy tumor cells or tissues [40]. It has been reported that a transdermal microneedle patch containing melanin generates heat and promotes the uptake of tumor antigens by dendritic cells under near-infrared light. It also enhances the activity of anti-tumor vaccines and promotes anti-tumor immune responses [41].
Melanin or melanoidin themselves have good metal ion chelation properties and can bind various paramagnetic metal ions, such as gadolinium, iron, manganese, etc., without causing toxicity to humans. They have ideal biocompatibility and biosafety and are potential materials for T1 contrast agents in the MRI [36]. The longitudinal relaxation rate of MNP-Mn particles is significantly higher than that of Omniscan, which is about three times higher than that of gadolinium diamide. The intensity of the MRI signal reaches a maximum of 3 h after injection. Moreover, analysis with the cck-8 kit showed that MNP-Mn particles are virtually non-cytotoxic [39]. In addition to binding metal ions, melanin can also enhance the signals of MRI by binding liposomes. After intravenous injection of LIP-Mel in living mice for a certain time, the MRI images of the tumor site become bright, and the MRI signals are gradually enhanced [206].
6.4. Application in the Cosmetics Industry
The development and application of natural active ingredients are favored in the cosmetic industry. Melanin is a natural nanocomponent with a variety of active ingredients. Firstly, melanin, as a non-toxic natural pigment, can be used as a natural colorant in cosmetics. Secondly, the antioxidant and anti-radiation activity of melanin can be added to cosmetics as a photoprotective agent to prolong the aging time of cosmetics. In addition, it has a certain opsonizing effect on the skin, acting as an anti-aging agent, similar to vitamin C and vitamin E [159,207].
This entry is adapted from the peer-reviewed paper 10.3390/ijms24054360
This entry is offline, you can click here to edit this entry!