In the last thirty years, there has been a significant increase in clinical interest in “natural” medicine, with a particular emphasis on the widespread use of plant products in the microbiological field. The increased resistance of microorganisms to antimicrobial drugs, and the emergence of newer diseases, require the urgent development of new, more effective drugs. Plants, due to the great biological and structural diversity of their components, can constitute a unique and renewable source for the discovery of new antimicrobial compounds. The use of essential oils is appealing and increasingly being investigated among new therapeutic approaches based on medicinal plants and their extracts [
1,
2].
Aromatherapy, a branch of alternative medicine that claims essential oils and other aromatic compounds have curative properties, has revived interest in essential oils in recent decades. Essential oils are concentrated hydrophobic liquids derived from plant secondary metabolism that contain volatile aroma compounds. Although they are widely employed in traditional medicine, the fragrance industry, and food flavoring and preservation, their good antimicrobial significance has only lately been recognized [
3,
4,
5,
6]. This new trend is due to the spread of microorganisms that are resistant to conventional antimicrobial agents, as well due to increased research into the antimicrobial activity of essential oils [
1,
7,
8]. Drug resistance can arise because of bacteria and yeasts’ rapid “mutation”, which alters cellular membrane proteins in a way that prevents drugs from recognizing them [
9]. In fact, most bacteria have the ability to alter membrane-associated macromolecules, the penicillin-binding proteins (PBPs), and enzymes that play key roles in the biosynthesis of the main component of the cell wall, the peptidoglycan. PBPs are the primary targets of β-lactam antibiotics. Many bacteria have the ability to produce a wide range of enzymes, including endo- and eso-β-lactamases or transferases, which can inactivate β-lactam and aminoglycoside antibiotics, respectively [
10]. Efflux pump modifications, or a significant decline (by 50%) in ergosterol levels in fungal cellular membranes, which can no longer be the drugs’ target site, might cause yeasts to become resistant to antifungal treatments, particularly azoles [
11]. In addition to these issues, antibiotics used to treat pathogen microorganisms may cause severe side effects, particularly in patients undergoing prolonged therapeutic treatment, or may alter the microbiome, which is critical for intestinal and overall body eubiosis. Antimicrobial agents, by increasing dysbiosis, create an ideal environment for pathogen microorganism colonization and subsequent infections, with the possibility of recurrent episodes. Therefore, with the wide use of synthetic and semi-synthetic antimicrobial drugs, the spread of drug-resistant clinical isolates has increased, and research is increasingly oriented towards natural products, such as essential oils, as useful antimicrobial resources [
1,
4,
6,
12]. Essential oils, on the other hand, contain hundreds of naturally active ingredients in varying proportions, which eliminates the risk of antibiotic resistance because microbes cannot adapt to their heterogeneous structure. Although the literature is fragmentary and incomplete, there is evidence that essential oils are active against a wide range of microorganisms, including bacteria, fungi, parasites, and viruses, with the ability to eliminate pathogens while preserving “friendly” microbiota [
3,
5,
13,
14].
However, good antimicrobial activity alone is insufficient from a clinical point of view. In fact, clinical experience shows that the therapeutic efficacy of conventional antimicrobial agents is dependent on both their direct effect on a given microorganism and the host immune system’s activity [
15,
16,
17,
18,
19,
20]. As with conventional drugs, in order to eradicate the infectious agent, it is critical to assess the potential influence of essential oils on host defense mechanisms in order to identify compounds that can stimulate them rather than interfere negatively with them [
15,
16,
17,
18,
19,
20].
2. Antimicrobial Potential of Essential Oils
It has shown the real efficacy of essential oils, and in particular, the ability of some of them to inhibit the growth of yeasts and filamentous fungi (dermatophytes,
Fusarium), already at low concentrations [
7]. This is the case of the essential oils of
Thymus vulgaris (thyme red), pine, clove, oregano, TTO, and MPP, which are proven to be the most effective, showing MIC values that are, in some cases, very low, especially towards the strains of
Cryptococcus neoformans, C. albicans,
C. glabrata, and
C. krusei [
33,
34,
35]. These findings are of great importance in light of the evident resistance that many strains of
C. glabrata show against traditional therapies with azoles, and the fact that
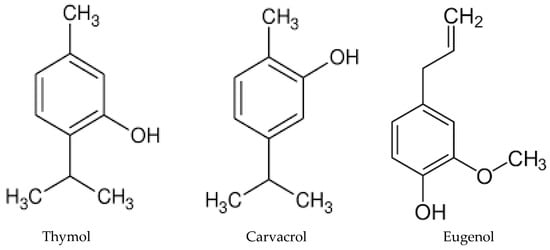
C. krusei is naturally resistant to fluconazole. It is not surprising that thyme, oregano, and clove essential oils exhibit a high inhibition of fungal growth, as their main components are phenols, such as thymol (26.52%
v/
v), carvacrol (62.61%
v/
v), and eugenol (77.5%
v/
v), compounds characterized by high bioavailability. These components may determine yeast membrane deterioration through oxidative stress and alteration of the antioxidant defense system even at low concentrations, leading to altered yeast virulence (
Figure 1) [
8,
35].
Figure 1. Chemical structures of thymol, carvacrol, and eugenol, the main components of thyme, oregano, and clove essential oils, respectively.
Pine essential oil, surprisingly, show, for some strains of non-
albicans Candida (
C. krusei,
C. norvegensis,
C. lusitaniae,
C. valida) and uncommon yeasts (
C. neoformans,
Pichia etchellsii/
carsonii,
Saccharomyces cerevisiae) higher inhibiting activity than thyme, clove, and oregano. Its major components are terpene hydrocarbons, such as α-pinene (55.76%
v/
v) and β-pinene (9.034%
v/
v) [
8] which, as many studies have already shown, possess high antimicrobial abilities, not only against fungi, but also against bacteria and viruses, proving to be one of the most active classes of compounds of plant origin (
Figure 2) [
8,
34,
35,
39,
40,
41].
Figure 2. Chemical structures of α- and β-pinene.
Since 2011, the effects of thyme red oil (thymol chemotype), one of the most active oils from a microbiological point of view, were investigated at subinhibitory and inhibitory concentrations to see how it may influence the intracellular killing activity by human PMNs against
C. albicans (thyme sensitive) [
42,
43] and
C. krusei [
44] naturally resistant to fluconazole, in comparison with conventional antifungal drugs .
For intracellular studies, human PMNs from healthy subjects were infected with yeast cells and treated with the essential oil at 1/2 × MIC and 1 × MIC. Intracellular killing values were expressed as the yeast survival index (SI) after 30, 60, and 90 min of incubation. Essential-oil-free controls were also included. In addition, in order to provide a frame of reference for the activity of this oil, its time–kill curve without PMNs was also evaluated. Extracellular studies in broth were performed using thyme red oil from 0.1 to 2 × MIC. Viable yeasts were enumerated at 0, 30, 60, 90, 120, 180 min, and 24 h [
42]. The influence of the essential oil on intracellular and extracellular activity was compared with that of fluconazole, caspofungin, and anidulafungin, some of the most widely used drugs in the prophylaxis and treatment of candidiasis [
32,
45].
When compared to essential-oil-free controls, thyme red oil significantly stimulates the killing of intracellular
C. albicans and
C. krusei by PMNs, even at subinhibitory concentrations (1/2 MIC), with killing rates overlapping (for
C. albicans) or significantly higher (for
C. krusei) than those observed with fluconazole and anidulafungin. Caspofungin-containing systems further inhibit the survival of
C. albicans, with fungicidal activity higher, both at 1/2 × MIC and 1 × MIC, than that of essential oil and fluconazole [
42,
43,
44].
In the absence of PMNs, the activity of essential oil against
C. albicans and
C. krusei is only fungistatic but greater than it is in the presence of fluconazole and/or anidulafungin [
42,
44]. In fact, even at 1/2 MIC, oil kills
C. krusei more efficiently than anidulafungin does in the same experimental conditions at higher concentrations. The fact that thyme red oil only has fungistatic activity suggests that the increased fungicidal intracellular killing by PMNs is the result of thyme red oil and phagocytes interacting favorably, similar to what is observed with conventional drugs [
29,
32,
33]. The mechanism of intracellular killing enhancement by oil is still unknown, even if a possible synergistic antifungal interaction with human phagocytes could help to explain why this essential oil, which appears to be only fungistatic in time–kill assays, has great efficacy in killing yeast cells once simultaneously incubated with PMNs.
Other essential oils, such as tea tree oil,
Melaleuca alternifolia (TTO) and
Mentha x piperita of Pancalieri-MPP (Torino, Italy), at sub-MIC (1/4 and 1/8) concentrations, on human PMNs’ capacity to kill a clinical strain of
C. krusei resistant to both fluconazole and anidulafungin [
56]. Anidulafungin was used as a comparison to assess the intraPMN killing potential of these essential oils. The viability of intracellular yeasts is significantly reduced by both TTO and MPP at 1/4 × MIC (42–50% vs. 46–61%) and 1/8xMIC (55–62% vs. 49–67%), with killing percentages 2–3 times higher than in controls (22–33%) and systems containing anidulafungin at 1/2 × MIC (27–33%). However, both oils at 1/8 × MIC significantly enhance the intracellular killing of
C. krusei by PMNs, with killing values comparable or higher than those observed at higher concentrations (1/4 × MIC). This is probably due to the fact that TTO and MPP at 1/4 × MIC are toxic, and they could interfere with the PMN functionality and killing capacity. On the contrary, the presence of 1/8 × MIC of both oils stimulates yeast killing by PMNs, indicating that the decreasing concentrations do not cause lower candidacidal activity.