Arsenic contamination in soil and water is a major problem worldwide. Inorganic arsenic is widely present as arsenate and arsenite. Arsenic is transferred to crops through the soil and irrigation water. It is reported to reduce crop production in plants and can cause a wide array of diseases in humans, including different types of cancers, premature delivery, stillbirth, and spontaneous abortion. Arsenic methyltransferase (AS3MT) in the human body converts inorganic arsenic into monomethylarsonic acid and dimethylarsinic acid, which are later excreted from the body. Arsenic transfer from the soil to grains of rice involves different transporters such as Lsi1, Lsi2, and Lsi6. These transporters are also required for the transfer of silicate, which makes them important for the plant.
- arsenate
- arsenite
- rice
- cancer
- arsenic transporters
1. Effect of Arsenic on Plants
| Plant | Change in Morphological and Physiological Parameters | Reference |
|---|---|---|
| Black Gram Vigna mungo | Increased activity of APX, SOD, and peroxidase (POD), decreased activity of Catalase (CAT), reduced photosynthetic pigment, and increased lipid peroxidation | [3] |
| Wheat Triticum aestivum |
Decreased biomass of roots, stems and spikes, decreased number of spikes per plant, stunted root and stems, necrosis, and wilting of leaf margins | [4] |
| Soybean Glycine max |
Decreased number of lateral roots, thickening and darkening of roots, necrotic and slimy root tips, darker and thicker stems, inhibited leaf development, affected biomass and root cell death, thin-walled parenchyma cells, reduced root cortex area, increased total peroxidase and superoxide dismutase (SOD) activity in As(III) treatment compared to As(V), and decreased chlorophyll a | [5] |
| Blackgram Vigna mungo | Root and shoot growth decreased, fresh root weight, fresh shoot weight, and fresh total biomass reduced, and shoot weight more affected than root weight | [6] |
| B. juncea | Decreased shoot length and number of leaves, increased activity of enzymes SOD, CAT, POD, APX, GR, and MDA content in a 30-day old plant, while only SOD and GR increased in 60 days old, protein content enhanced in 30-day old plant | [7] |
| Barley Hordeum vulgare | Decreased root and shoot dry weight, SOD and CAT activity increase | [8] |
| V. radiata | Length, root oxidizing capacity of seedling, and dry biomass decreased, ferric chelate reductase (FCR) activity increased, increased proline, H2O2, malondialdehyde (MDA), SOD, and CAT | [9] |
| C. moschata | Reduced phenolic compound and flavonoids, root length, reduced shoot length, root and shoot fresh weight, root and shoot dry weight. Reduced Chl a and Chl b, increased H2O2 and MDA content while increasing the activity of SOD, CAT, and APX | [10] |
| Rice | Decreased number of tillers, above-ground biomass, and grain yield | [11] |
| Rice cultivar Lalat Oryza sativa ssp. indica | Decreased germination with weight more affected than length, roots turned black, more accumulation in shoots when As(V) used, otherwise more in roots, increased lipid peroxidation, SOD, APX, and glutathione reductase (GR) activity | [12] |
| Spinach | Decreased plant biomass and chlorophyll-a and b content, increased oxidative stress through high H2O2, upregulated CAT, guaiacol peroxidase (GPX), APX, and SOD | [13] |
| Maize Zea mays |
Decreased dry weight of roots and shoot, As(V) decreased chlorophyll content | [14] |
2. Arsenic Accumulation Depends on Plant Genotype
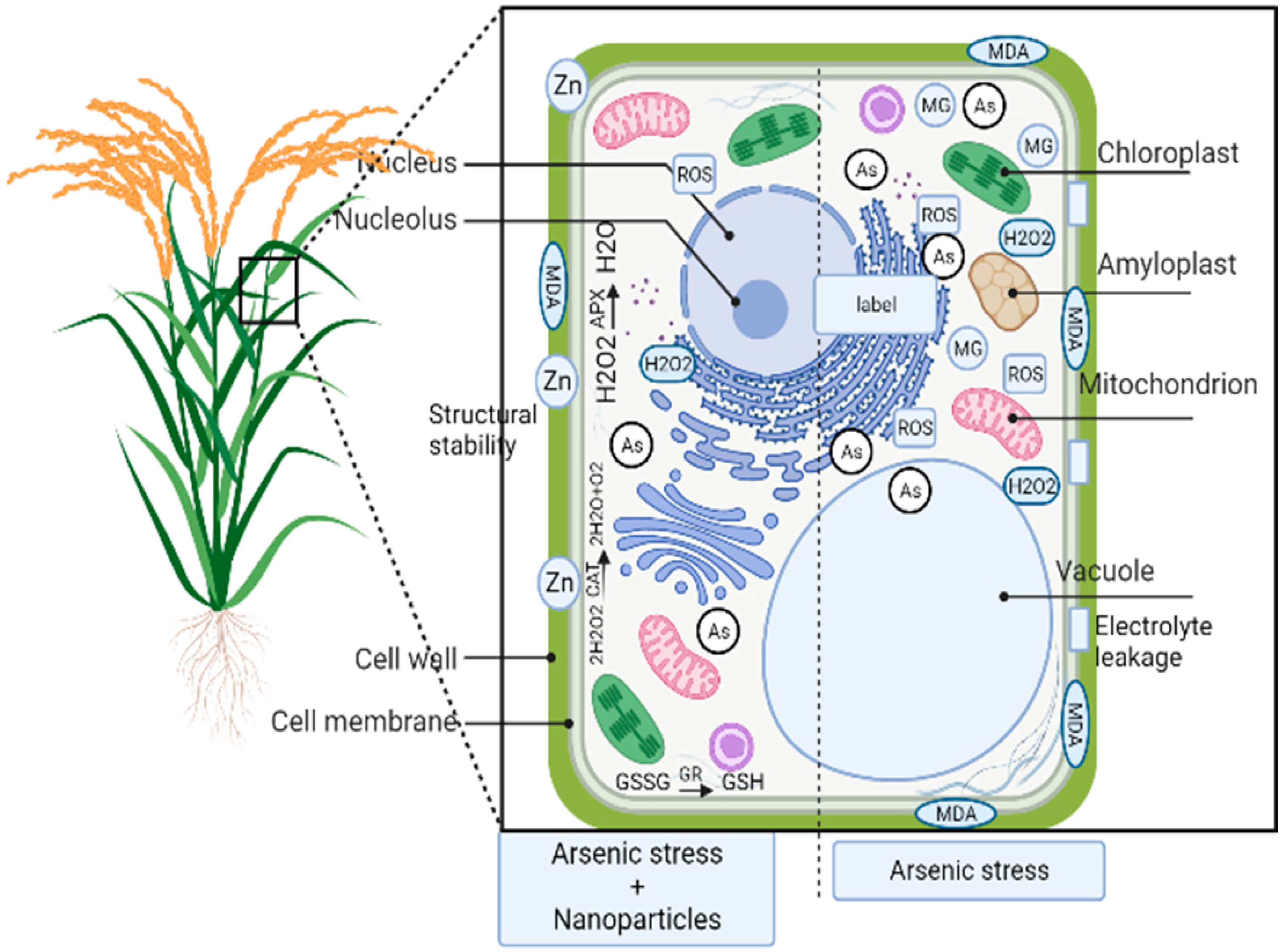
3. Application of Nanotechnology to Counter Negative Effects of Arsenic on Plants
Plant Extracts Mitigate Arsenic Effects
This entry is adapted from the peer-reviewed paper 10.3390/agriculture13020401
References
- Sandhi, A.; Yu, C.; Rahman, M.M.; Amin, M.N. Arsenic in the water and agricultural crop production system: Bangladesh perspectives. Environ. Sci. Pollut. Res. Int. 2022, 29, 51354–51366.
- Sun, S.K.; Chen, Y.; Che, J.; Konishi, N.; Tang, Z.; Miller, A.J.; Ma, J.F.; Zhao, F.J. Decreasing arsenic accumulation in rice by overexpressing OsNIP 1;1 and OsNIP 3;3 through disrupting arsenite radial transport in roots. New Phytol. 2018, 219, 641–653.
- Srivastava, S.; Sinha, P.; Sharma, Y.K. Status of photosynthetic pigments, lipid peroxidation and anti-oxidative enzymes in Vigna mungo in presence of arsenic. J. Plant Nutr. 2017, 40, 298–306.
- Zhang, W.D.; Liu, D.S.; Tian, J.C.; He, F.L. Toxicity and accumulation of arsenic in wheat (Triticum aestivum L.) varieties of China. Phyton 2009, 78, 147–154.
- Armendariz, A.L.; Talano, M.A.; Travaglia, C.; Reinoso, H.; Oller AL, W.; Agostini, E. Arsenic toxicity in soybean seedlings and their attenuation mechanisms. Plant Physiol. Biochem. 2016, 98, 119–127.
- Shamim, M.Z.; Pandey, A. Effects of arsenic toxicity on morphological characters in blackgram (Vigna mungo L.) during early growth stage. Cell. Mol. Biol. 2017, 63, 38–43.
- Kanwar, M.K.; Bhardwaj, R. Arsenic induced modulation of antioxidative defense system and brassinosteroids in Brassica juncea L . Ecotoxicol. Environ. Saf. 2015, 115, 119–125.
- Zvobgo, G.; Lwalaba JL, W.; Sagonda, T.; Mapodzeke, J.M.; Muhammad, N.; Shamsi, I.H.; Zhang, G.P. Alleviation of arsenic toxicity by phosphate is associated with its regulation of detoxification, defense, and transport gene expression in barley. J. Integr. Agric. 2019, 18, 381–394.
- Shabnam, N.; Kim, M.; Kim, H. Iron (III) oxide nanoparticles alleviate arsenic induced stunting in Vigna radiata . Ecotoxicol. Environ. Saf. 2019, 183, 109496.
- Mushtaq, T.; Shah, A.A.; Akram, W.; Yasin, N.A. Synergistic ameliorative effect of iron oxide nanoparticles and Bacillus subtilis S4 against arsenic toxicity in Cucurbita moschata: Polyamines, antioxidants, and physiochemical studies. Int. J. Phytoremediation 2020, 22, 1408–1419.
- Panaullah, G.M.; Alam, T.; Hossain, M.B.; Loeppert, R.H.; Lauren, J.G.; Meisner, C.A.; Ahmed, Z.U.; Duxbury, J.M. Arsenic toxicity to rice (Oryza sativa L.) in Bangladesh. Plant Soil 2009, 317, 31–39.
- Shri, M.; Kumar, S.; Chakrabarty, D.; Trivedi, P.K.; Mallick, S.; Misra, P.; Shukla, D.; Mishra, S.; Srivastava, S.; Tripathi, R.D.; et al. Effect of arsenic on growth, oxidative stress, and antioxidant system in rice seedlings. Ecotoxicol. Environ. Saf. 2009, 72, 1102–1110.
- Shahid, M.; Khalid, S.; Saleem, M. Unrevealing arsenic and lead toxicity and antioxidant response in spinach: A human health perspective. Environ. Geochem. Health 2022, 44, 487–496.
- Várallyay, S.; Bódi, É.; Garousi, F.; Veres, S.; Kovács, B. Effect of arsenic on dry weight and relative chlorophyll content in greeningmaize and sunflower tissues. J. Microbiol. Biotechnol. Food Sci. 2021, 2021, 167–169.
- Zemanová, V.; Popov, M.; Pavlíková, D.; Kotrba, P.; Hnilička, F.; Česká, J.; Pavlík, M. Effect of arsenic stress on 5-methylcytosine, photosynthetic parameters and nutrient content in arsenic hyperaccumulator Pteris cretica (L.) var. Albo-lineata. BMC Plant Biol. 2020, 20, 130.
- Mallick, I.; Bhattacharyya, C.; Mukherji, S.; Dey, D.; Sarkar, S.C.; Mukhopadhyay, U.K.; Ghosh, A. Effective rhizoinoculation and biofilm formation by arsenic immobilizing halophilic plant growth promoting bacteria (PGPB) isolated from mangrove rhizosphere: A step towards arsenic rhizoremediation. Sci. Total Environ. 2018, 610, 1239–1250.
- Mohd, S.; Shukla, J.; Kushwaha, A.S.; Mandrah, K.; Shankar, J.; Arjaria, N.; Saxena, P.N.; Narayan, R.; Roy, S.K.; Kumar, M. Endophytic fungi Piriformospora indica mediated protection of host from arsenic toxicity. Front. Microbiol. 2017, 8, 754.
- Singh, A.P.; Dixit, G.; Kumar, A.; Mishra, S.; Singh, P.K.; Dwivedi, S.; Trivedi, P.K.; Chakrabarty, D.; Mallick, S.; Pandey, V.; et al. Nitric oxide alleviated arsenic toxicity by modulation of antioxidants and thiol metabolism in rice (Oryza sativa L.). Front. Plant Sci. 2016, 6, 1272.
- Singh, N.; Marwa, N.; Mishra, J.; Verma, P.C.; Rathaur, S.; Singh, N. Brevundimonas diminuta mediated alleviation of arsenic toxicity and plant growth promotion in Oryza sativa L. Ecotoxicol. Environ. Saf. 2016, 125, 25–34.
- Andrade, H.M.; Oliveira, J.A.; Farnese, F.S.; Ribeiro, C.; Silva, A.A.; Campos, F.V.; Neto, J.L. Arsenic toxicity: Cell signalling and the attenuating effect of nitric oxide in Eichhornia crassipes . Biol. Plant. 2016, 60, 173–180.
- Dixit, G.; Singh, A.P.; Kumar, A.; Mishra, S.; Dwivedi, S.; Kumar, S.; Trivedi, P.K.; Pandey, V.; Tripathi, R.D. Reduced arsenic accumulation in rice (Oryza sativa L.) shoot involves sulfur mediated improved thiol metabolism, antioxidant system and altered arsenic transporters. Plant Physiol. Biochem. 2016, 99, 86–96.
- Mishra, S.; Alfeld, M.; Sobotka, R.; Andresen, E.; Falkenberg, G.; Küpper, H. Analysis of sublethal arsenic toxicity to Ceratophyllum demersum: Subcellular distribution of arsenic and inhibition of chlorophyll biosynthesis. J. Exp. Bot. 2016, 67, 4639–4646.
- Kaur, S.; Chowhan, N.; Sharma, P.; Rathee, S.; Singh, H.P.; Batish, D.R. β-Pinene alleviates arsenic (As)-induced oxidative stress by modulating enzymatic antioxidant activities in roots of Oryza sativa . Ecotoxicol. Environ. Saf. 2022, 229, 113080.
- Talukdar, P.; Hartley, S.E.; Travis, A.J.; Price, A.H.; Norton, G.J. Genotypic differences in shoot silicon concentration and the impact on grain arsenic concentration in rice. J. Plant Nutr. Soil Sci. 2019, 182, 265–276.
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212.
- Wu, C.; Wang, Q.; Xue, S.; Pan, W.; Lou, L.; Li, D.; Hartley, W. Do aeration conditions affect arsenic and phosphate accumulation and phosphate transporter expression in rice (Oryza sativa L.)? Environ. Sci. Pollut. Res. 2018, 25, 43–51.
- Diba, F.; Khan, M.Z.H.; Uddin, S.Z.; Istiaq, A.; Shuvo, M.S.R.; Ul Alam, A.R.; Hossain, M.A.; Sultana, M. Bioaccumulation and detoxification of trivalent arsenic by Achromobacter xylosoxidans BHW-15 and electrochemical detection of its transformation efficiency. Sci. Rep. 2021, 11, 21312.
- Yan, G.; Chen, X.; Du, S.; Deng, Z.; Wang, L.; Chen, S. Genetic mechanisms of arsenic detoxification and metabolism in bacteria. Curr. Genet. 2019, 65, 329–338.
- Mukherjee, P.; Mitra, A.; Roy, M. Halomonas rhizobacteria of Avicennia marina of Indian Sundarbans promote rice growth under saline and heavy metal stresses through exopolysaccharide production. Front. Microbiol. 2019, 10, 1207.
- Ahmad, P.; Alyemeni, M.N.; Al-Huqail, A.A.; Alqahtani, M.A.; Wijaya, L.; Ashraf, M.; Kaya, C.; Bajguz, A. Zinc oxide nanoparticles application alleviates arsenic (As) toxicity in soybean plants by restricting the uptake of as and modulating key biochemical attributes, antioxidant enzymes, ascorbate-glutathione cycle and glyoxalase system. Plants 2020, 9, 825.
- Wu, F.; Fang, Q.; Yan, S.; Pan, L.; Tang, X.; Ye, W. Effects of zinc oxide nanoparticles on arsenic stress in rice (Oryza sativa L.): Germination, early growth, and arsenic uptake. Environ. Sci. Pollut. Res. 2020, 27, 26974–26981.
- Liu, H.; Li, P.; Yu, H.; Zhang, T.; Qiu, F. Controlled fabrication of functionalized nanoscale zero-valent iron/celluloses composite with silicon as protective layer for arsenic removal. Chem. Eng. Res. Des. 2019, 151, 242–251.
- Ahmed, T.; Noman, M.; Manzoor, N.; Shahid, M.; Hussaini, K.M.; Rizwan, M.; Ali, S.; Maqsood, A.; Li, B. Green magnesium oxide nanoparticles-based modulation of cellular oxidative repair mechanisms to reduce arsenic uptake and translocation in rice (Oryza sativa L.) plants. Environ. Pollut. 2021, 288, 117785.
- Gautam, A.; Pandey, A.K.; Dubey, R.S. Azadirachta indica and Ocimum sanctum leaf extracts alleviate arsenic toxicity by reducing arsenic uptake and improving antioxidant system in rice seedlings. Physiol. Mol. Biol. Plants 2020, 26, 63–81.
- Mahajan, P.; Singh, H.P.; Kaur, S.; Batish, D.R.; Kohli, R.K. β-Pinene moderates Cr (VI) phytotoxicity by quenching reactive oxygen species and altering antioxidant machinery in maize. Environ. Sci. Pollut. Res. 2019, 26, 456–463.
- Farias, G.J.; Frescura, D.V.; Boligon, A.A.; Trapp, C.K.; Andriolo, L.J.; Tedesco, B.S.; Bernardy, K.; Schwalbert, R.; Del Frari, K.B.; Carey, M.; et al. Chemical properties and protective effect of Rosmarinus officinalis: Mitigation of lipid peroxidation and DNA-damage from arsenic exposure. J. Appl. Bot. Food Qual. 2018, 91, 1–7.