Two biological mechanisms can alter the Na balance and hydromineral homeostasis [
7]. The first is largely determined by the intake of NaCl, which accumulates in the extracellular space and leads to the displacement of water from cell to extracellular space, producing cell dehydration. In this situation, the appetite for Na is inhibited and its excretion is increased (natriuresis), while the intake of water is enhanced and the excretion of fluid is reduced (antidiuresis). Conversely, a reduction in ECF osmolality/natremia leads to the inhibition of water intake and promotion of diuresis, increasing the appetite for Na and suppressing natriuresis. The second mechanism results from the loss of intravascular fluid (hypovolemia), one of the components of ECF. Diarrhea, vomiting, hemorrhages, sweating, renal disease, and cardiovascular disorders are sometimes accompanied by the loss of intravascular volume. Hypovolemia triggers compensatory behavioral and physiological reactions to conserve Na and body fluids. These include a thirst and appetite for Na, especially for isotonic drinks containing the same sodium chloride concentration as intravascular fluid (0.15 M), as well as antidiuretic and antinatriuretic responses to retain water and Na. Conversely, ECF hypervolemia reduces the intake of water and Na and increases diuresis and natriuresis.
2. Behavioral Mechanism: Salt Intake
The Na ion [Na
+] is an essential micronutrient that cannot be generated by endogenous processes and must be obtained through the diet [
9]. The need to maintain an appropriate Na balance has led living organisms to develop mechanisms that identify the presence of Na the instant it enters the body [
10]. The first level of detection is located in the oral cavity, from where it accesses the digestive system and is absorbed in the blood.
2.1. Detection and Processing in the Oral Cavity
Na is detected by the taste system within the oral cavity. The mineral generates a salty taste that guides and maintains its consumption [
11,
12,
13,
14]. The gustatory stimulus used as prototype to study the perception of this taste was table salt (NaCl), physiologically the most important salt in the diet, which induces the purest salty taste [
11,
15], and biologically the most relevant to maintain biochemical balance [
16].
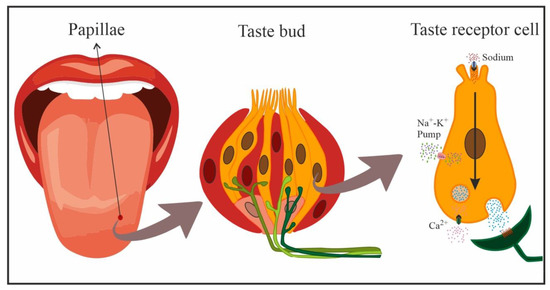
In mammals, gustatory receptors are observed throughout the oral cavity, especially on the tongue, where they are located on protuberances designated papillae. There are four types of papillae according to their shape and localization: fungiform (anterior two-thirds of the tongue), foliate (side toward the back), circumvallate (on the back), and filiform (anterior two-thirds of the tongue). Cells that detect NaCl are mainly located in fungiform papillae [
12,
17]. In general, gustatory papillae can contain several hundreds of taste buds, onion-shaped organs (
Figure 2) that house taste receptor cells (50–100 cells each), i.e., the cells responsible for detecting and transducing taste stimuli (transforming chemical signals into neural activity). In the taste bud, these cells are mechanically held together by tight junctions that separate apical areas from the remaining basolateral region, forming an almost impermeable barrier. Only the apical membrane above the tight junctions (2–3% of the total membrane area of the cell) is exposed to saliva, while the basolateral area remains in a relatively constant interstitial milieu [
12,
13,
14,
18].
Figure 2. Schematic representation of cells in a taste bud, and transduction of NaCl taste in a taste receptor cell (yellow: taste receptor cells, red: support cells, pink: immature precursor cells). Taste receptor cells form synapses with afferent nerve fibers that penetrate the taste bud, crossing the basal membrane.
Taste receptor cells are bipolar neuroepithelial cells with finger-like projections (microvilli) on their apical end that project towards the oral cavity. These microvillar processes protrude into the oral milieu through a small opening, the bud pore, in order to sample the chemical environment. When salty stimuli dissolved in saliva come into contact with these microvilli, the taste cells generate a biological signal that is transmitted to afferent neural fibers that penetrate the base of taste buds and extensively ramify. These afferent nerve fibers form synapses with taste receptor cells and respond to excitatory neurotransmitters released by the latter [
19], relaying this information to the brain to code the taste sensation [
10,
14].
There appear to be two types of salty taste receptor cells in the lingual epithelium. One of them, specialized in coding the perceptual quality of “salty” taste, is highly selective for Na, being activated by low–medium concentrations of NaCl and blocked by lingual application of amiloride (a diuretic drug). The other type is activated by high (aversive) concentrations of NaCl (>150 mM), is amiloride-insensitive, and is non-selective for Na (other salts are equally effective). The latter type of taste receptor cells appears to be innervated by neural pathways associated more with aversion than with taste coding [
12,
14,
19,
20].
Taste reception begins when chemicals interact with taste receptors located at the apical segment of the taste cells. The receptors that mediate the transduction of salty taste are ion channels [
12,
19], designated epithelial Na channels (ENaCs), which are specialized in the detection or recognition of sodium chloride [
11,
12,
15,
16,
19]. Interestingly, and as expected, it has been demonstrated that the gustatory response to Na in deficient animals can be augmented by some of the hormones involved in its regulation, e.g., aldosterone or angiotensin II (Ang II), through an increase in the number of active amiloride-sensitive Na channels in the apical membrane of taste cells [
9,
21,
22].
With the increase in salivary Na concentration, this cation directly enters cells through amiloride-sensitive ENaCs on the apical membrane, following electrochemical gradients. The increase in positive charges inside the taste receptor cell creates a depolarization of the cell membrane that generates action potentials passively conducted towards the basolateral region of the taste receptor cell in a dose-dependent manner, i.e., the action potential frequency is dependent on the stimulus intensity. Here, the opening of voltage-activated Ca
2+ gives rise to the influx of Ca
2+ ions and the release of the corresponding neurotransmitter onto postsynaptic gustatory afferent fibers (
Figure 2) that carry the taste signal to higher-order systems [
10,
16]. Detection by the nonspecific amiloride-insensitive mechanism is less well known and appears to take place below the apical membrane and taste cell tight junctions (basolateral detection) [
13,
16].
NaCl taste afferents are the peripheral processes of pseudounipolar neurons located in geniculate cranial ganglia. The chorda tympani nerve, with cell bodies in the geniculate ganglion, appears to be mainly responsible for transmitting information on the presence of Na in the oral cavity to the brain [
18,
23,
24]. Thus, bilateral transection of this nerve severely disrupts the ability to discriminate Na from other salts [
25]. In humans, the salty taste is generated at concentrations above 40 mM, a very similar threshold to that in rats (30 mM) but much higher than that in mice (10 mM). The rise in salivary Na
+ concentration also increases the neural response (stimulus dependent), meaning that the Na concentration can be estimated as a function of the neural discharge rate. This capacity allows individuals to ingest the appropriate amount of NaCl to maintain homeostasis, i.e., around 1% NaCl (0.17 M), in the ECF [
10,
20,
26].
Accordingly, the chorda tympani nerve appears to contain two types of fibers that differ in their sensitivity to NaCl; one type responds solely to salty taste (Na
+ and Li
+; NaCl specialist, highly specific fibers), while the other responds to a wide range of acid and salty stimuli (NaCl generalist), including Na [
3,
13,
18]. Activity of the former but not the latter is blocked by applying amiloride on the tongue in various animal species, as expected [
10].
Chorda tympani nerve fibers with oral saltiness information terminate in a highly specific region of the rostral nucleus of the solitary tract (rNST), especially at its most anterior pole and lateral division [
18,
27,
28].
2.2. Sodium Detection and Processing in the Gastrointestinal System
Various studies have demonstrated that the most caudal levels of the digestive system are also prepared for Na detection (visceral sensory signals) [
29,
30,
31,
32,
33].
Information on the entry of Na can be transmitted from the gastrointestinal system to the CNS in a rapid manner, via neural mechanisms, or more slowly, via the circulatory system (humoral pathway) [
29,
32,
33,
34,
35,
36].
The neural pathway is vagal in nature [
37], and Na
+ detection by subdiaphragmatic vagal afferents is well documented [
33,
38]. The presence of NaCl-sensitive receptors has been demonstrated in the gastrointestinal mucosa [
39,
40,
41,
42] and at hepatic level [
43,
44]. These receptors appear to be connected to small-diameter fibers, mostly unmyelinated vagal fibers that respond to the presence of hypertonic NaCl with a mean latency of around 10–20 s [
39,
40,
41,
42]. The first relay of visceral-vagal information from post-oral segments of the digestive system takes place in caudal segments of the nucleus of the solitary tract (NST), from which it is remitted to other brain structures [
27,
45,
46].
Na absorption into the circulatory system largely takes place in the distal small bowel and the colon [
47,
48,
49]. In these digestive segments, Na
+ is taken up through absorptive cells of the epithelium, which connect on one side with the intestinal lumen (apical region, brush border) and on the other (basolateral region) with blood and lymphatic capillaries. In the small intestine, these cells are organized into digitiform projections or intestinal villi that significantly augment (around 600-fold) the absorption surface area; however, this organization is less marked towards distal ends of the intestine and is virtually absent at the most distal region of the colon [
50].
Na absorption is transcellular in the intestine. In a first step, the ion penetrates from the intestinal lumen into the cytoplasm of the absorptive epithelial cell following electrochemical gradients and strongly assisted by Na-dependent transporters (e.g., Na
+/glucose and Na
+/cotransporter amino acids or Na–hydrogen exchangers (NHEs), or by Na reuptake through ENaCs mainly present in the distal colon [
50,
51]. Once within the epithelial cells, Na
+ is actively transported towards paracellular spaces by the Na
+/K
+ ATPase pump in the basal and lateral membranes of the cells, avoiding retrograde diffusion, from where it enters the bloodstream [
47,
48,
50].
The modality of Na
+ absorption and the intestinal segment in which it occurs mainly depends on the circumstances. In this way, it appears to occur preferentially in the small intestine after meals, coupled with nutrient absorption, nutrient-dependent Na absorption, Na
+/glucose, and Na
+/cotransporter amino acids. In contrast, it largely takes place in the ileum and colon through NHE or ENaCs during interdigestive periods [
50,
52]. Under Na deficiency conditions, hormones such as aldosterone can increase Na uptake in the intestinal epithelium, stimulating the expression of the transporters that participate in its absorption (especially at colon level). Consequently, there is virtually no Na loss under these circumstances, with <0.5% of the intestinal content of Na being excreted via the feces under normal conditions [
47,
51,
53,
54].
Once in the circulatory system, Na is widely distributed throughout the organism, accessing circumventricular brain structures, which lack a blood–brain barrier, via fenestrated capillaries [
55,
56], and accessing cerebrospinal fluid (CSF) via ion transporters (e.g., ENaC) in epithelial cells of the choroid plexus [
57,
58,
59,
60]. Blood and CSF both have a [Na+] concentration of 144 mOsm/L [
61].