This paper enlightens a deep understanding of the molecular control of MF in EGMS liens and exploring the regulatory driving forces that function efficiently during plant adaptation under a changing environment.
- Hybrid rice, EGMS, male fertility, sterility
1. Introduction
2. Development of Hybrid Rice Technologies
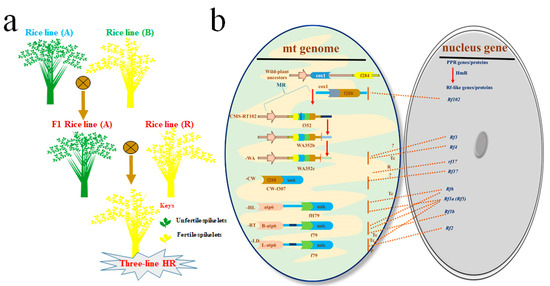
Previous reports highlighted that crop production was not substantial to support increasing population around the world, a lot of regions of the world became the victim of uprising hunger, in the 1950s. Progressively, rice breeding technology has advanced via the introduction of semi-dwarf varieties (HYV) that were high yielding [7]. The maize and wheat enhancement programs paved the way toward high yield and improvement against lodging and disease resistance through genetic manipulation of the semi-dwarf (sd-1) gene among various species. These findings enabled the scientists of the International Rice Research Institute (IRRI) to develop the first semi-dwarf rice that has unique properties such as medium height, lodging resistant, a greater number of panicles and grains leading to high yield. During 1966, there was a more dynamic shift that was attained through the green-revolution by genetic manipulation of the rice IR8 variety. It was harboring the semi-dwarf (sd-1) gene and is known as the miracle rice, termed as international rice 8 (IR8), and enhanced rice yield [8,9]. This discovery and successful manipulation of the sd1 (the semi-dwarf mutant) gene in crops was the first “green-revolution” that facilitated in hunger eradication in the developing countries [10]. To feed the fast-growing population, crop yield was enhanced effectively in several parts of the world by introducing high yield new cultivars during the past few decades. However, rice production with marvelous effects has been in progress since as early as 1926 by investigating heterosis in rice [11,12]. Though the possibility to adopt hybrid-rice (HR) technology was started the first time in 1966 by Yuan Long-ping, later he was pronounced as the father of HR in China [13]. Being a scientist, global FS is an enormous task for human beings. HR gained popularity due to high yield and great advantages as compared to the inbred cultivar/lines [14]. It has also been evidenced that heterosis exploitation is a common phenomenon in crops and the most effective breeding tool against food scarcity worldwide. HR seed production comprises the crossing among two well defined genetically important inbred parental cultivar/lines (one female line and another male line). HR technology provides a better result to improve yield by producing superior quality containing F1 HR over its pure inbred or dwarf lines [15,16]. It is a practical way to enhance rice production by using F1 hybrids, which provided 20–25% more yield benefit over pure rice breeds [13,17,18]. Over time, the tremendous progress in the form of hybrid-breeding technology greatly benefited agriculture by HR with high yield and better tolerance against stressors (e.g., biotic stresses as the diseases, pests, and pathogen infestation and abiotic stresses as the drought, heat, salt, etc.) as compared to the inbred lines/varieties [4,19,20,21]. Rice is a self-pollinated cereal crop; its male fertility (MF, described as the release of the workable gametes or functional pollens that can fertilize female gametes) control demands the male sterility system to generate HR lines/varieties. Male sterility (MS, defined as the production of nonworkable gametes or nonfunctional pollens that can fecundate female gametes), acts as the central player in MF regulation for hybrid-seed production and provides the incredible germplasm to explore rice reproductive development and harness the influence of hybrid-vigor to gain more seed production, as the key breeding tools [22,23]. Overall, MS is grouped into two types such as cytoplasmic-male-sterile (CMS) and environment-sensitive-genic-male-sterility (EGMS) [22]. After the discovery of the male sterility system and its application in HR technology, that originated in China by using male sterility inducing nuclear and cytoplasmic genes to generate cytoplasmic-male-sterile (CMS) lines and CMS also termed as the three-line HR technology, displayed the innovative step towards HR production (Figure 1a) [22,24,25].
- Palanisamy, D.; Marappan, S.; Ponnuswamy, R.D.; Mahalingam, P.S.; Bohar, R.; Vaidyanathan, S. Accelerating hybrid rice breeding through the adoption of doubled haploid technology for R-line development. Biologia 2019, 74, 1259–1269.
- Khush, G.S. Strategies for increasing the yield potential of cereals: Case of rice as an example. Plant Breed. 2013, 132, 433–436.
- Ma, X.; Su, Z.; Ma, H. Molecular Genetic Analyses of Abiotic Stress Responses During plant Reproductive Development; Oxford University Press: Oxford, UK, 2020.
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975.
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273.
- Eshed, Y.; Lippman, Z.B. Revolutions in agriculture chart a course for targeted breeding of old and new crops. Science 2019, 366, eaax0025.
- Kikuchi, F. Semidwarfing genes of high yielding rice varieties in Japan. In Rice Genetics I: (In 2 Parts); World Scientific: Singapore, 1986; pp. 285–295.
- Peng, S.; Tang, Q.; Huang, J.; Zou, Y.; Cui, K.; Zhang, Y.; He, F.; Laza, R.; Visperas, R. Yield Attributes and Nitrogen-Use Efficiency of “Super” Hybrid Rice; Accelerating Hybrid Rice Development, International Rice Research Institute: Manila, Philippines, 2010; pp. 419–428.
- Khush, G.S. What it will take to Feed 5.0 Billion Rice consumers in 2030. Plant Mol. Biol. 2005, 59, 15950.
- Myers, N. The next green revolution: Its environmental underpinnings. Curr. Sci. 1999, 76, 507–513.
- Jones, J.W. Hybrid vigour in rice. Am. Soc. Agron. 1926, 18, 424–428.
- Ramaiah, K. Inheritance of flowering duration in rice. Indian J. Agric. Sci. 1933, 3, 377–410.
- Yuan, L. The execution and theory of developing hybrid rice. Agric. Sci. China 1977, 1, 27–31.
- Weng, J.; Suhai Gu, S.; Wan, X.; Gao, H.; Guo, T.; Su, N.; Lei, C.; Zhang, X.; Cheng, Z.; Guo, X.; et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 2008, 18, 1199–1209.
- فيرماني ، أرز هجين SS. في التقدم في الهندسة الزراعية ؛ إلسفير: أمستردام ، هولندا ، 1996 ؛ المجلد 57 ، ص 377-462.
- فيرماني ، إس إس ؛ الشمس ، ZX ؛ مو ، TM ؛ جوهر ، أ. Mao، CX عقم الذكور في الأرز. في دليل تربية الأرز الهجين المكون من سطرين ؛ المعهد الدولي لبحوث الأرز: لوس بانوس ، الفلبين ، 2003 ؛ ص 5 - 14.
- كروبف ، م. كاسمان ، ك. بينغ ، س. ماثيوز ، ر. Setter ، T. الفهم الكمي لإمكانات العائد. في كسر حاجز العائد ؛ المعهد الدولي لبحوث الأرز: لوس بانوس ، الفلبين ، 1994 ؛ ص 21 - 38.
- تشو ، جي ؛ تشين ، واي. ياو ، دبليو. تشانغ ، سي. شيه ، دبليو. هوا ، ياء ؛ شينغ ، واي. شياو ، ياء ؛ Zhang ، Q. التركيب الجيني لتغاير المحصول في هجين أرز النخبة. بروك. ناتل. أكاد. علوم. الولايات المتحدة الأمريكية 2012 ، 109 ، 15847–15852.
- يانغ ، ر. بياو ، زي. وان ، سي ؛ لي ، جي ؛ روان ، العاشر ؛ Bai، J. تربية مجموعات أرز جابونيكا الهجين المكونة من ثلاثة أسطر مع محتوى نشا عالي المقاومة باستخدام التحديد بمساعدة الواسمات الجزيئية. تربية. علوم. 2020 .
- تشين ، س. تسنغ ، جي ؛ هاو ، م. جيانغ ، هـ. Xiao ، Y. تحسين صهر الأرز ومقاومة الزرع البني لخط PTGMS C815S في الأرز الهجين المكون من سطرين من خلال التحديد بمساعدة العلامة. مول. تربية. 2020 ، 40 ، 21.
- أنصاري ، مور ؛ شاهين ، ت. بخاري ، س. حسنين ، ت. التحسين الجيني للأرز لتحمل الإجهاد الحيوي واللاأحيائي. تركي. جيه بوت. 2015 ، 39 ، 911-919.
- تشين ، إل. Liu ، YG عقم الذكور واستعادة الخصوبة في المحاصيل. Annu. القس بيول النبات. 2014 ، 65 ، 579-606.
- ماير ، إي. جوزيف جوتليب كولروتر مساهمات في علم الأحياء. أوزوريس 1986 ، 2 ، 135–176.
- مروحة ، Y. ؛ Zhang ، Q. التوصيف الجيني والجزيئي لعقم الذكور في فترة الضوء والحساسية الحرارية في الأرز. ريبرود النبات. 2018 ، 31 ، 3–14.
- تشانغ ، ياء ؛ لي ، X.-M. ؛ لين ، H.-X. ؛ تشونغ ، ك. تحسين المحاصيل من خلال مقاومة درجات الحرارة. Annu. القس بيول النبات. 2019 ، 70 ، 753-780.
This entry is adapted from the peer-reviewed paper 10.3390/ijms21217868