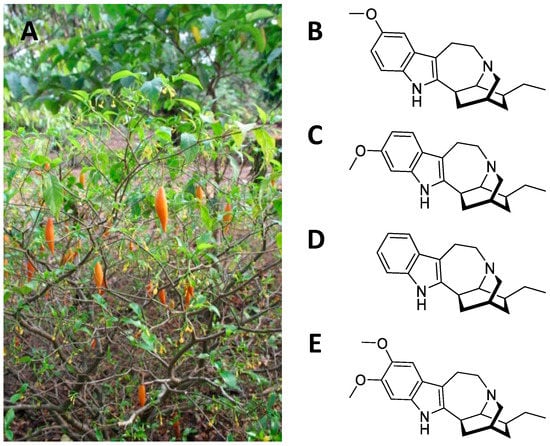
Figure 4. (
A) Three-dimensional model representing hSERT (PDB ID: 6DZZ). Residues coloured in blue are those located in a zone within 5 Å of the serotonin. (
B) Magnification depicting ibogaine (in purple) and serotonin (in orange) fitted into the central binding pocket of hSERT. The image was obtained by aligning the PDB ID 6DZZ and 7MGW structures. The highlighted residues are those most relevant to the interaction of the two molecules [
57]. The superimposition (RMSD 1.152 Å) was completed using the MatchMaker tool in UCSF Chimera [
70]. The artworks were produced using the same software.
2.4. Therapeutic Hypotheses
Over the recent years, ibogaine has been taken into consideration as a therapeutic agent, able to treat or assist the treatments for drug addictions such as heroin [71], cocaine [72], methamphetamine [73], cannabis and crack [74]. The advantage of the use of ibogaine as a therapeutic for drug addiction resides in the fact that a single large dose seems effective in blocking withdrawal and reduces cravings in drug-dependent individuals [72], contrary to standard pharmacological treatments which typically require a prolonged administration. Indeed, ibogaine administration, unlike commonly used treatments such as methadone, shortens the time needed for withdrawal to 2–3 days, effectively speeding up the detoxification process [72].
Along with the growing use of psychedelics in the treatment of psychiatric disorders, research about the therapeutic effects of ibogaine has begun to take its first steps in this direction. For example, Fernandes-Nascimento et al. (2022) recently reported a single case study of ibogaine microdosing for the treatment of type II bipolar disorder (BPD) [78]. Microdosing, a growing trend in the psychedelics field, is defined as recurrent administration of small doses of psychedelic drugs, with little to no identifiable acute drug effects, with the intention of improving mental health and general wellbeing, enhance cognitive performances and boost mood [79,80]. It reported a significant improvement of symptoms in a 47-year-old woman with a 20-year history of BPD treated for 60 days, with two daily administrations of ibogaine (4 mg per administration, approximately 1% of a full conventional single dose). This finding is consistent with previous preclinical studies undertaken in rats [81], which characterized behavioural effects induced by acute ibogaine and noribogaine (20 and 40 mg/kg, i.p., single injections for each dose) in rats, assessing depression-like symptoms using the forced swim test. Both drugs induced a dose- and time-dependent antidepressant effect, without significant changes in locomotor activity.
3. Echinocactus williamsii
3.1. Ethnobotany
The genus
Echinocactus includes a dozen perennial cacti originating from Central America and South America with a globular or cylindrical structure, with slow growth, which can reach a diameter of 90 cm and develop a fleshy root [
82,
83,
84]. Specifically,
Echinocactus williamsii (
Figure 5A), also called peyote, is a small cactus that grows in the Rio Grande valley and neighbouring deserts [
82,
83,
84]. The psychoactive alkaloids are concentrated mainly in the epigeal part [
85]. In particular, the “vegetable buttons” known as mescal correspond to the tips of the cactus cut and left to dry in the sun and subsequently used by shamans for mystical-religious rites [
8,
86,
87,
88,
89]. Furthermore, in some Central American locations it is eaten fresh, dried in paste or as tea [
8]. About 20 alkaloids are extracted from
Echinocactus williamsii, the most important of which is mescaline [
85] (
Figure 5B). Mescaline is an alkaloid, biosynthetically derived from amino acids, with a chemical structure similar to monoamine neurotransmitters, and psychomimetic activity [
85,
90]. It is used by the natives of Central America both for its exciting and intoxicating properties and for the attractive hallucinations it causes [
8,
86,
87]. In particular, it is capable of altering the basal psychic state leading to a condition of psychomotor arousal with euphoria and lively abnormal psychosensory manifestations (olfactory, auditory and above all visual hallucinations), symptoms of depersonalization and alterations in space-time orientation [
90,
91]. Some hallucinatory phenomena, called synesthetic phenomena, are characteristic, consisting in the fact that a perception, for example auditory, causes lively chromatic sensations. The substance is active at a dose between 100 and 400 mg per person orally [
90,
92].
Figure 5. Flowered Echinocactus williamsii (A); chemical structure of mescaline (B).
3.2. Central Nervous System Pathways
Regarding the pharmacological aspects, mescaline is considered an agonist of the 5-HT2A and 5-HT2C receptors of serotonin, and of the α2A adrenergic receptor [90,93,94,95]. In contrast, mescaline shows low affinity for other serotonergic receptors and for dopamine receptors and monoamine transporters [85,91,93,94,95]. The hallucinogenic effects appear to be regulated by mescaline agonism towards the 5-HT2A receptor [85,95]. However, it has been observed in animal models that blockade of dopamine receptors with haloperidol can halt the characteristic effects of mescaline, suggesting the additional involvement of other neuronal pathways [91].
3.3. Structural and Computational Studies
Navarro et al., after having generated and validated a 5-HT2A receptor model by homology modelling studies, performed in-silico experiments, including docking, to evaluate the interaction of mescaline with this target [
101]. Molecular docking studies have been carried out in two different ways. Firstly, using a protocol that involved a rigid docking mode (RRA), which gives the ligand greater degrees of conformational freedom, something that the receptor is not granted. The second docking was carried out in flexible mode (FRA) where, in addition to the ligand, degrees of freedom are also granted in the 4 Å vicinity of the binding site of the receptor.
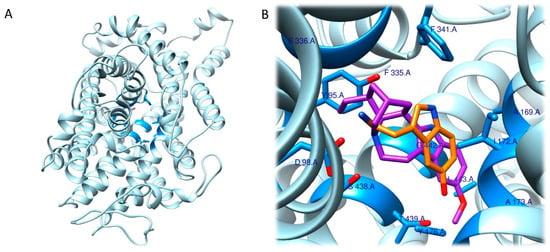
As expected, RRA and FRA methods displayed two different interaction patterns. Interactions obtained with RRA included one weak π–alkyl interaction between the aromatic ring of the ligand and Val156 (5.18 Å), three strong π–σ interactions between the aromatic ring and Val235 (2.77 Å), a hydrogen bond between one H atom from the NH3+ group and the backbone of Phe234 (2.26 Å), and another hydrogen bond between the O atom from the 5-OMe substituent and Ser239 (2.61 Å) (Figure 6A,B).
Figure 6. Three-dimensional model depicting the 5-HT2A receptor (PDB ID 7WC4). Residues coloured in pink are those involved in interactions with mescaline determined by the RRA method (
A,
B) and the FRA method (
C,
D) [
70,
101].
The FRA method provided different results in terms of interaction distances and involved residues. More specifically, one weak, four intermediate and one strong interaction were identified. The weak π–π interaction was identified between the aromatic ring and Phe339 (5.05 Å). The four intermediate interactions comprehend a π–π interaction between the aromatic ring and Val156 (4.83 Å), a H bond between the O atom (as an acceptor) from the 5-OMe substituent and the backbone of Ser239 (3.55 Å), a H bond between the 3-OMe substituent and Asp155 (3.34 Å), and a H bond between the 4-OMe substituent and Ser159 (3.50 Å). The strong interaction detected is a salt bridge interaction between one H atom from the NH3+ group and Asp231 (2.41 Å). Of note, the common residues involved according to both methods were only Val156 and Ser239 (Figure 6C,D).
3.4. Therapeutic Hypotheses
Mescaline, along with other psychedelics such as LSD and psilocybin, has been widely employed in psychiatric and therapeutic contexts, before its use was restricted by the Schedule I of the UN Convention on Drugs in 1967 [91]. Due to its regulation, clinical research over recent decades has been heavily limited [90]. Preclinical studies using animal models have been employed to investigate the effect of mescaline on behaviour. As mentioned previously, mescaline is able to induce an increase in locomotor activity and exploratory behaviour, although some studies report a dose-dependent reduction with an inverted U-shape curve (locomotion and exploratory behaviour increase at lower doses and decrease at higher doses) [102,103,104]. Mescaline is also able to induce an increase in animal reactivity [105].
4. Psychotria Viridis
4.1. Ethnobotany
About 1400 shrubby species that grow in tropical areas around the world belong to the
Psychotria genus [
108,
109,
110,
111,
112,
113,
114,
115]. In particular,
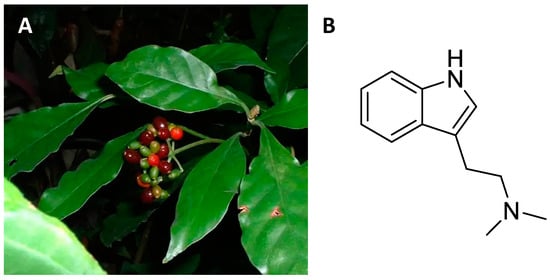
Psychotria viridis (
Figure 7A) is a member of the rubiaceae family that grows spontaneously in the humid areas of Central and South America [
116]. The leaves of
Psychotria viridis are rich in N,N-dimethyltryptamine (DMT,
Figure 7B), a psychedelic alkaloid [
117]. Depending on the different American ethnic groups, the dried leaves can be smoked or added to a drink that also contains
Banisteroriopsis caapi leaves [
118]. This drink is called
ayahuasca and is used by shamans to contact deceased ancestors [
119]. The hallucinogenic effect of ayahuasca is due to the presence of carbolines (harmine and harmaline) which inhibit the action of monoamine oxidase and enhance the action of DMT [
9,
10,
118,
119].
Figure 7. Psychotria viridis with red infructescences (A); chemical structure of N,N-dimethyltryptamine (B).
4.2. Central Nervous System Pathways
Like most psychedelics, DMT is considered a partial agonist of serotonin receptors. Receptor binding studies have shown a DMT affinity to the 5-HT2A receptor of around 75 nM [
117,
120]. However, the 5-HT2A receptor appears to be necessary but not sufficient to explain the hallucinogenic phenomena reported in previous studies [
121,
122]. Indeed, the anxiolytic response observed in mouse models appears to be mediated by DMT binding to 5-HT1D and 5-HT3 receptors [
117]. The psychedelic response to DMT can also be mediated by interaction with 5-HT1A and 5-HT2C receptors, although the head twitch response in mice is blocked only by 5-HT2A receptor antagonists [
120,
121].
DMT can also regulate the activity of ionotropic NMDA receptors directly, by modulating memory and learning processes, or indirectly, by activating the sigma-1 receptor [
129,
130,
131,
132]. The sigma-1 receptor is a chaperonin localized in the endoplasmic reticulum of cells of the cerebral or peripheral tissues [
133]. Given the widespread distribution of the sigma-1 receptor, it has been studied in various diseases and neurobiological conditions such as addiction, depression, amnesia, pain, stroke and cancer [
133]. DMT binds to the sigma-1 receptor at micromolar concentrations, contributing to the psychedelic response [
130,
132]. Agonism at the sigma-1 receptor is also involved in neurotrophic and neuroprotective processes [
134,
135]. Although totally convincing data on the direct involvement of DMT in the neuroprotective activity of the sigma-1 receptor has not been reported, it cannot be excluded [
130,
136].
4.3. Structural and Computational Studies
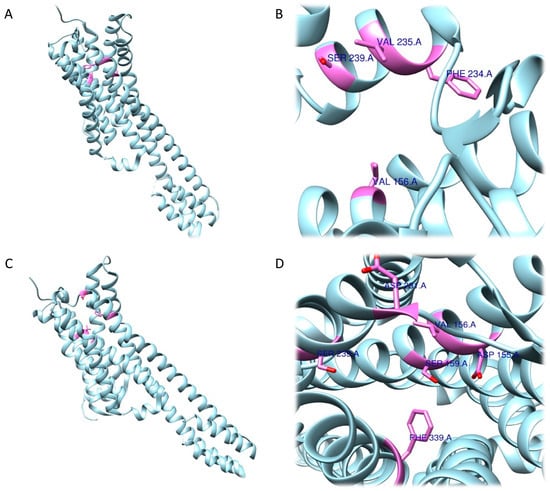
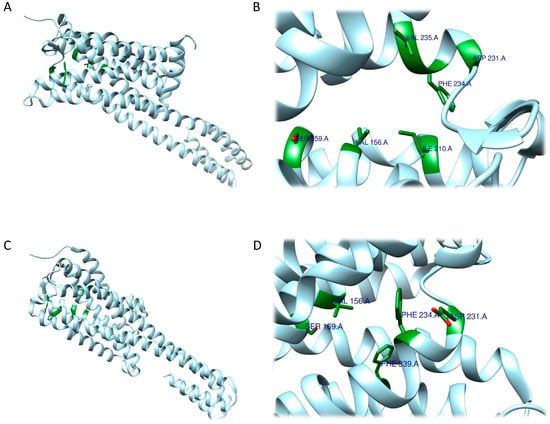
The RRA shows a π–σ interaction with Ile210 (2.83 Å), a H bond between the H(N) atom of the indole moiety and Asp231 (2.17 Å), a H bond between NHMe2 and Ser159 (3.43 Å), an amide–π stacking interaction with Phe234 (3.98 Å), an amide–π stacking interaction with Phe234 (4.12 Å), a π–alkyl interaction with Val235 (4.95 Å) and Val156 (5.01 Å), and a π–alkyl interaction with Ile210 (5.00 Å) and Val156 (4.95 Å). Once the interactions were identified, they were classified according to the distance between the atoms involved as strong (distance ≤ 3 Å), intermediate (distance between 3–5 Å), and weak (distance ≥5 Å). Overall, two strong, five intermediate, and two weak interactions were detected (Figure 9A,B).
Figure 9. Three-dimensional model depicting 5-HT2A receptor (PDB ID 7WC4). Residues coloured in green are those involved in interactions with DMT determined by the RRA method (
A,
B) and FRA method (
C,
D) [
70].
FRA simulations showed a H bond of the H(N) from the indole moiety with Ser159 (2.3 Å), an attractive charge interaction of the N atom from NHMe2 with Asp231 (5.46 Å), a H bond between the H(N) from NHMe2 with an oxygen atom of Phe234 (2.90 Å), a π–alkyl interaction with Val156 (5.27 Å), a π–π interaction with Phe339 (5.32 Å) and another π–π interaction with Phe339 (4.84 Å). FRA results also show the presence of additional interactions between Asp231 and Phe234 and the NHMe2 group of the DMT molecule (Figure 9C,D).
At a molecular level, the mechanism of the interaction between DMT and 5-HT1B is not yet fully known. In this regard, the aim of the work was to understand how the presence of DMT could somehow influence the stability of the receptor at the atomic level. To overcome the limitation of the current treatments there is an ongoing search for molecules that can contribute to the treatment of disorders such as anxiety and depression [
143]. The resolved structure of the receptor (PDB ID: 4IAR) was used for molecular docking studies. The binding energy obtained from the docking was −6.65 ± 0.07 kcal/mol, comparable to that computed for serotonin (−6.50 ± 0.14 kcal/mol), a result that indicates a good value of binding energy. By comparing the behaviour of DMT and serotonin, the schloars also confirmed that the two molecules share most interactions with the 5-HT1B receptor.
4.4. Therapeutic Hypotheses
Although DMT is known for its psychedelic properties, latest research has shown its clinical utility, addressing a variety of medical conditions (especially those of the central nervous system). As a matter of fact, in the last decade DMT has been investigated for its neuroprotective effects, which are thought to be mediated by binding with the sigma-1 receptor [145]. In-vitro experiments have displayed DMT’s cytoprotective effect against hypoxia: diverse cell types (human cortical neurons derived from iPSC, dendritic cells, monocyte-derived macrophages) cultured in severe hypoxic conditions have been treated with DMT. Results suggest that DMT is able to prevent cellular stress and robustly boosts cell survival [146]. DMT is also able to activate the subgranular zone of the dentate gyrus in-vivo, a part of the hippocampal formation in which most adult neurogenesis takes place. This neurogenic propriety seems to have a robust functional effect, since DMT-treated mice (2 mg/kg, i.p., for 21 days) performed better in memory and learning tasks, for which the hippocampus plays an essential role [148]. The increased neurogenesis and subsequent cognitive improvement provided by DMT treatment could have profound implications for neurodegenerative diseases such as Parkinson’s (PD) and Alzheimer’s (AD) and for medical conditions defined by significant loss of neural cells in affected areas of the CNS, such as stroke. Previous literature [149,150,151,152] indicates that neurogenesis and neuroprotective factors could be implemented as a valid tool in the therapeutic strategies for psychiatric and neurological disorders, aiding in the restoration and preservation of residual functionality in the cerebral regions affected by the disease. This concept has recently been assessed using DMT-containing Ayahuasca in an in-vitro model of Parkinson’s disease.
Apart from clinical and preclinical studies, ayahuasca’s therapeutic potential seems to be confirmed by several observational studies, conducted on participants taking ayahuasca in naturalistic setting [163,164,165,166]. Although the clear limitation of observational studies, it is important to notice that this field of literature is unanimous in confirming the potential healing properties of ayahuasca and DMT, reporting robust improvements in psychopathological symptoms, boost in overall mood and improvements in mental health well-being that are persistent over time after a single dose.
5. Psilocybe Cubensis
5.1. Ethnobotany
Psilocybe mushrooms are Basidiomycota members of the family Strophariaceae [
171]. The species of this genus are cosmopolitan, the best known are the
P. cubensis (
Figure 11A) and the
P. Mexicana, which grow in Central America and have been historically used for shamanic rites [
172]. Mushrooms are prepared in various ways depending on the shaman’s preferences [
86]. They can be eaten fresh, dried, or infused and trigger hallucinogenic responses in users. The ritualistic use of Psilocybe mushrooms in Mesoamerica is documented by the 14th century Codex “Yuta Tnoho” or “Vindobonensis Mexicanus I”, which depicts a sacred ceremony where deities consume sacred mushrooms prior to the first dawn [
173]. However, the discoveries of the Tassili mural in Algeria, reporting fungi associated with
P. mairei, and the Selva Pascuala mural in Spain, a rock painting representing fungoid figures that have been associated with
P. hispanica, date the use of these natural products back 7000–9000 years [
174].
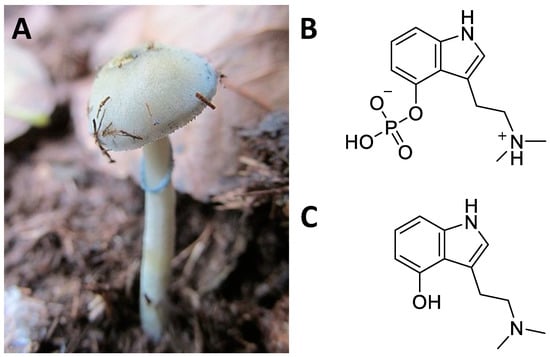
Figure 11. Psilocybe cubensis (A); chemical structure of psilocybin (B); psilocin (C).
5.2. Central Nervous System Pathways
After oral administration of increasing doses of psilocybin, it loses its phosphate group and is totally converted to psilocin in the acidic environment of the stomach or by alkaline phosphatase in the intestine and kidneys [
182,
183,
184]. Therefore, evaluations of the pharmacological profile of psilocybin have been performed with its main derivative [
182]. Psilocin is identified in plasma within half an hour of administration and reaches peak concentrations within three hours [
184]. Plasma AUC increases in proportion to dose, indicating linear pharmacokinetics in doses between 0.3 and 0.6 mg/kg of psilocybin [
183,
184]. The average bioavailability of psilocin is around 50% and its average half-life is around three hours. More than 80% of psilocin is metabolized by glucuronidation and released in the urine as psilocin-O-glucuronide [
183,
184].
Psilocin has also shown a good receptor affinity for the 5-HT2A receptor with the EC50 for receptor occupancy around 2 μg/L [
185,
186,
187]. However, psilocin is considered a partial agonist of the 5-HT2A receptor. Indeed, compared with serotonin, psilocin shows an efficiency of less than 40% in the Ca
2+ mobilization assay [
188]. The link with the 5-HT2A receptor is responsible for the “mystical” hallucinatory effects induced by psilocin [
185]. However, a certain psilocin receptor non-specificity common to many psychedelics has been confirmed. In increasing order of affinity, psilocin can also bind to 5-HT2B, 5-HT1D, dopamine D1, 5-HT1E, 5-HT1A, 5-HT5A, 5-HT7, 5-HT6, D3, 5-HT2C and 5-HT1B receptors [
185,
188].
Simultaneously with the psychedelic effects, psilocin has shown antidepressant and anxiolytic effects at the basis of its therapeutic use. Indeed, psilocin, acting on serotonergic receptors, deactivates or normalizes the hyperactivity of the medial prefrontal cortex which is typically hyperactive during depressive phenomena [
32,
189]. This antidepressant action also seems to involve limbic areas including the amygdala, which is considered the centre of perception and processing of emotions. Psilocin causes an indirect variation of the dopaminergic and serotonergic tone in a differentiated way in some mesolimbic areas of the brain [
191].
5.3. Structural and Computational Studies
Concerning the 5-HT2A receptor, Cao et al. (2022) resolved and deposited the structure of psilocin-5-HT2A complex (PDB ID: 7WC5) [194,195]. The work performed by Cao et al. showed that, in addition to the central binding pocket, psilocin is also able to interact with a second binding site where the indole group fits into a pocket, described as an extended binding site, mainly containing hydrophobic residues. A salt bridge is formed between the basic nitrogen and the Asp155 residue and a hydrogen bond between Asn352 and the OH- group in the indole of psilocin was detected. Subsequently, during the analysis of the interactions between the 5-HT2A receptor and psilocin, additional residues involved in the formation of the complex, such as Val156, Phe339, Asn363, Leu228, and Trp151, were identified [195].
The study of the role of psilocybin and of its active metabolite then continued, though focused on another receptor, 5-HT2C. In this case, molecular docking was enrolled, and Gumpper et al. (2022) showed that psilocin is also able to bind to the binding pocket of the receptor [196]. Here the compound establishes bonds with a salt bridge with Asp134 through the amine compound, a π–stacking interaction between the tryptamine ring and Phe328, and another bond between the indole -OH and Asn331. Further, other interactions with Trp130 and Phe327 were identified in the assembly.
5.4. Therapeutic Hypotheses
Psilocybin is a medium-lasting, well-tolerated classic psychedelic that is potentially safe and effective [
189,
197,
198]. Among psychedelics, psilocybin represents the path-opening drug in modern psychedelic-supervised therapy. In 2021, the FDA twice designated psilocybin the designation of “breakthrough therapy”, to accelerate its drug development and review process. At first, the FDA supported psilocybin for the treatment of depression and severe treatment-resistant depression. Subsequently, the FDA supported the Compass Pathways company in testing psilocybin as a promising therapeutic option for major depressive disorder (MDD). After the conclusion in 2022 of this clinical trial, in which 233 participants were enrolled, the scholars were able to demonstrate that a single administration, though of a high dosage of psilocybin (25 mg), was able to reduce the Montgomery–Åsberg Depression Rating Scale (MADRS), a clinical parameter used to establish the depression severity, by 12 points [
199].
Although the safety and efficacy of psilocybin as an antidepressant are the focus of most recent clinical trials (more than 100 clinical trials have been registered), there are still no definitive data that can establish whether psilocybin can offer substantial clinical improvement over existing antidepressant therapies [
200,
201]. It has recently been reported that the antidepressant response of psilocybin is not statistically stronger than the conventional antidepressant escitalopram [
202].
Again, the FDA supported an investigational new drug (IND) clinical trial to explore how psilocybin-assisted therapy impacts the treatment of anorexia nervosa (ClinicalTrials.gov Identifier: NCT04505189). For the study, a small group of patients with a primary diagnosis of anorexia nervosa as defined by DSM-V criteria will take part in eight study visits, including three psilocybin dosing sessions with varying doses up to the maximum of 25 mg per single session.
Studies are underway that aim to assess whether psilocybin is more feasible, tolerable, and efficacious for the treatment of post-traumatic stress disorders when administered alone or in combination with assisted therapy.
6. Claviceps Purpurea
6.1. Ethnobotany
Claviceps purpurea is a fungus, belonging to the Ascomycetes Clavicipetali, which infests cereal crops and in particular rye [
209] (
Figure 15A). The sclerotium,
Secale cornutum, is the richest part of the alkaloids (
Figure 15B). These include ergolinic alkaloids derived from lysergic acid such as ergotamine, ergometrine, ergocristine, ergocriptine, and ergoconine [
210]. Alkaloids have found multiple applications in the cardiovascular and gynaecological fields [
211]. However, the greatest impact on the CNS came with the semi-synthetic synthesis of D-lysergic acid diethylamide (LSD,
Figure 15C), deriving from lysergic acid. Indeed, in 1938 the chemist Albert Hofmann synthesized LSD for the first time in the Swiss laboratories of the Sandoz AG Pharmaceutical Company, deriving from lysergic acid, and accidentally tested its hallucinatory and psychedelic effects [
212,
213]. LSD has had a major social impact since the 1960s by profoundly influencing Western culture [
212,
213]. The cerebral effects of LSD concern the emotional–ideational aspects and above all sensory perception (colours are perceived more vividly) [
214,
215,
216,
217]. These hallucinogenic effects affect the sight, hearing, touch, and perception of one’s body. Furthermore, subjects who used LSD experienced introspective trips that enabled them to perceive inner problems and reality from other points of view, beyond the usual schemes [
214,
215,
216,
217].
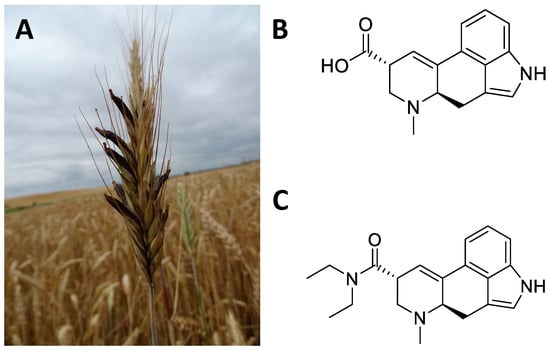
Figure 15. Sclerotium of Claviceps purpurea on an ear of grass (A); chemical structure of lysergic acid (B); lysergic acid diethylamide (synthetic compound) (C).
6.2. Central Nervous System Pathways
LSD is totally absorbed in the intestine after oral administration. Absorption can be affected by the pH of the stomach and duodenum [
218,
219]. Indeed, the administration of LSD with food induces plasma concentrations that are halved when compared to administration on an empty stomach [
220]. The different routes of administration do not determine qualitative differences in the hallucinatory effects of LSD but only in the intensity and speed of onset [
220,
221,
222]. LSD can cross the blood–brain barrier as previously observed in mice, rats, cats and monkeys [
220,
221,
223]. In humans, after an administration of 2 μg/kg i.v. LSD levels were approximately 7 ng/mL after 30 min and disappeared after 10 h [
220]. Some variation was observed between species in the half-life of LSD at the same dosage and route of administration: 7 min in mice, 130 min in cats, 100 min in monkeys, and 175 min in humans [
223]. LSD is almost completely metabolized to 13- and 14-hydroxy-LSD and their conjugates glucuronic acid, 2-oxo-LSD, and nor-LSD and only a small part of the unchanged drug is excreted [
224]. LSD can be metabolized in humans to 2-oxo-LSD and 2-oxo-3-hydroxy-LSD by certain NADH-dependent liver microsomal enzymes [
220,
221,
223]. In addition, lysergic acid ethylamide (originating from dealkylation of the diethylamide radical at position 8 of the side chain), nor-LSD and di-hydroxy-LSD have also been identified in human blood and urine [
218,
219]. These metabolites, and LSD itself, can be identified in urine for up to four days after ingestion.
6.3. Structural and Computational Studies
To assess the action of the LSD, at the level of 5-HT2B receptor, Wacker et al. (2017) characterized the complex through computational studies and structural resolution techniques [
234]. Their analysis revealed that LSD can bind to the receptor within the orthosteric binding site with a volume of 2898.7 Å
3 [
235]. The ligand has been shown to be able to insert itself into the orthosteric binding pocket through a salt bridge that occurs between Asp135 and the basic nitrogen present in the structure. Subsequently, the schloars identified the way in which the ergoline moiety of LSD can establish aromatic contacts with Phe340 and Phe341, and how the nitrogen in the indole group forms a hydrogen bond with Gly221. Another component capable of forming interactions is the diethylamide moiety, which, thanks to one ethyl group, forms non-polar contacts with Leu132 and Trp131, while the other ethyl group interfaces with Leu362. Further interactions detected are Val136, Ser139, Leu209, Phe217, Ser222, Ala225, Trp337, and Asn344.
6.4. Therapeutic Hypotheses
The recent interest in the therapeutic effects of LSD is supported by several preclinical and clinical studies. Concerning preclinical studies, LSD has been tested to treat stress-induced anxiety-like behaviour in mice. In particular, male mice exposed to chronic restrain stress were treated daily with 30 μg/kg LSD for one week and the treatment was able to prevent the stress-induced anxiety-like behaviour, the decrease of cortical spine density and the serotonergic transmission decline [
33]. Interestingly, LSD treatment did not cause any anxiolytic or anti-depressant effect in non-stressed mice. LSD has also been demonstrated to exert antidepressant effects in rodents and to promote neuroplasticity in rat cortical neurons; a single administration of LSD 0.15 mg/kg i.p. was able to significantly reduce depressive-like behaviours in rats five weeks after treatment. Indeed, by activating the TrkB receptor, mTOR and 5-HT2A signalling pathways in the prefrontal cortex, it produced an increase in dendritic arbor complexity, promote dendritic spine growth, and stimulate synapse formation, thus revealing a fast-acting, robust and persistent antidepressant effect [
170,
238]. Additional repeating of LSD administration (0.13 mg/kg) for 11 days was able to reverse deficits in active avoidance learning in bulbectomised rats, a model of depression [
214].
In healthy human subjects, neuroimaging studies have demonstrated that LSD induces increases in functional brain connectivity between the thalamus and sensory–somatomotor cortical regions, and from the thalamus to the posterior cingulate cortex [
216]; on the other hand, it decreased connectivity to the temporal cortex [
216]. These anatomical changes could explain the LSD-induced effect in facilitating a novel experience of the self and its environment and in reducing rigid or ruminative thinking patterns typical of psychiatric disorders [
33].