4.1. Current Potential Biomarkers for the Screening, Diagnosis, and Prognosis of Retinal Vascular Diseases
Currently, great challenges remain in the screening, diagnosis, and prognosis of retinal vascular diseases. In general, the screening of retinal vascular diseases depends on direct and indirect ophthalmoscopy or fundus photographic technologies. However, these methods are less likely to be carried out owing to the long annual screening period required for susceptible populations, emerging economic burden levels, and the scarcity of trained vitreoretinal specialists [
54,
55]. Although ophthalmoscopy might be sufficient for a basic diagnosis of AMD, fluorescein angiography (FFA) is the gold standard for detecting and differentiating these diseases, as this method allows clinicians to accurately determine the status and circulation of retinal vessels in order to uncover earlier diagnostic pathological indicators (e.g., microaneurysms, hemorrhages, small veins, and so on [
56]), monitor disease progression through repeated examinations every 3–6 months for staging inference, and perform patient stratification through specific imaging findings. Nevertheless, its invasiveness and cumulative radiation toxicity limit its use in patients with cardiovascular and renal diseases, which are the most common comorbidities of retinal vascular diseases.
Severe proliferative vitreoretinopathy and neovascularization are common terminal changes in retinal and choroidal vascular diseases, and current therapeutic strategies have prolonged the time it takes patients to reach the terminal stages of these diseases. Nevertheless, a significant number of patients with poor treatment responses rapidly progress to the terminal stage within a short period. Presently, optical coherence tomography (OCT) and visual acuity are the main tools used to monitor the treatment responses and predict the prognosis of retinal and choroidal vascular diseases; however, inexperienced doctors are prone to generating inaccurate inferences through these methods. Therefore, there is an urgent need to discover more scientific and stable biomarkers for the screening, diagnosis, and prognosis of retinal vascular diseases.
Recent studies have reported potential biomarkers for resolving the problems stated above. Here, we discuss the roles of a classic biomarker for retinal vascular diseases, the vascular endothelial growth factor (VEGF) family. In retinal vascular diseases, retinal hypoxia and hypoperfusion trigger elevated levels of the VEGF family, including VEGF-A, VEGF-B, and placental growth factor (PlGF) in the intraocular environment, eventually leading to angiogenesis in the vascular and avascular zones, which is also well known as neovascularization [
57,
58,
59]. As important stable biomarkers for the development stage and prognosis of retinal vascular diseases, it has been confirmed that a significant correlation exists between high levels of VEGF-A in aqueous and vitreous fluids and neovascularization of the retina, disc, angle, and iris in the proliferative DR (PDR) stage as well as in ischemic retinopathies and ischemic RVO [
57,
60,
61].
Intravitreal injection of anti-VEGF agents is the first-line treatment strategy for ischemic retinal vascular diseases. Despite it being considered as the most promising biomarker, the VEGF family fails to comprehensively explain retinal vascular diseases. Other biomarkers, such as elevated levels of various cytokines, interleukin (IL)-1β, IL-2, IL-4, IL-5, IL-6, IL-10, interferon-γ, tumor necrosis factor-α, and VEGF, were observed in the aqueous humor or vitreous of patients with DR [
62,
63,
64], AMD [
65], RVO [
66], and ROP [
67]. However, all of these biomarkers were derived from a series of positive findings from faint single-aspect evidence sources, and further validation is necessary. Therefore, suitable and stable biomarkers for retinal vascular diseases remain elusive. Multi-omics approaches may simply be used to identify more precise, data-supported, stable, specific, and sensitive biomarkers with lower expenses, increased user-friendliness, and easier accessible detection methods by creating multi-dimensional global evidence chains from suitable tissues.
4.2. Omics-Based Biomarkers Discoveries
Newly-developed “omics-based biomarkers discovery” approaches may be potential solutions to the issues discussed above in previous sections, as these approaches have been frequently applied to screen out potential clinical detection indicators for cancers [
68,
69], type II diabetes [
70], cardiac vascular diseases [
71], central nervous system diseases [
72], and autoimmune diseases [
73]. In these approaches, as illustrated in
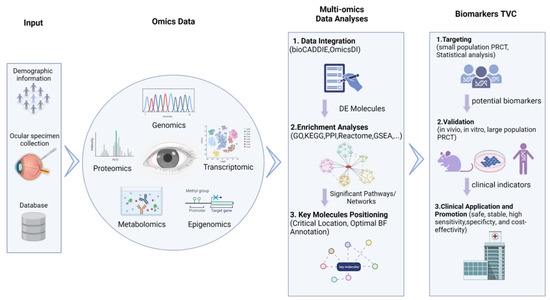
Figure 2, omics data derived from biological specimen sequencing experiments or public databases are integrated together in an untargeted and unbiased manner (i.e., the “hypothesis-free method”) for further analyses. These methods have shown great promise in the identification of precise, specific, and sensitive candidate biomarkers for the clinical detection of retinal vascular diseases to reduce the prevalence of low vision and blindness.
Figure 2. Multi-omics approach to identify biomarkers for retinal vascular diseases.
The flow chart illustrates the omics data-based biomarker discovery process, including omics data collection, multi-omics data integration analysis, biomarker targeting, validation, and clinical application.
4.2.1. Source of Omics Data
Regarding biological specimens for screening, early detection, and long-term monitoring of retinal vascular diseases, biomarkers in tears, saliva, sweat, and other systemic specimens have practical significance mainly due to the non-invasive collection processes and high concentrations of proteins and metabolites of these samples [
74,
75,
76,
77]. Moreover, if omics-based biomarker discovery is conducted using omics data from multiple studies, different types of omics data should be retrieved and downloaded from specified public databases, such as the recommended genomics databases (GWAS catalog:
https://www.ebi.ac.uk/gwas/ (accessed on 24 December 2022) GWAS central:
https://www.gwascentral.org/ (accessed on 24 December 2022) and dbGaP:
https://dbgap.ncbi.nlm.nih.gov/ (accessed on 24 December 2022)); transcriptomics databases (GEO:
https://www.ncbi.nlm.nih.gov/geo/ (accessed on 24 December 2022), Expression Atlas:
https://www.ebi.ac.uk/gxa (accessed on 24 December 2022), and ArrayExpress:
https://www.ebi.ac.uk/arrayexpress/ (accessed on 24 December 2022)); proteomics databases (Peptide Atlas:
http://www.peptideatlas.org/ (accessed on 24 December 2022), ProteomicsDB:
https://www.proteomicsdb.org/ (accessed on 24 December 2022), Human Proteome Map:
http://www.humanproteomemap.org/ (accessed on 24 December 2022), and Human Proteome Atlas:
http://v13.proteinatlas.org/ (accessed on 24 December 2022)); and metabolomics databases (Human Metabolome:
https://hmdb.ca/ (accessed on 24 December 2022) and MetabolomeExpress:
https://www.metabolome-express.org/ (accessed on 24 December 2022)).
4.2.2. Multi-Omics Data Integration Analysis
After obtaining a variety of omics data, the critical step of multi-omics data integration is the identification and visualization of differentially expressed (DE) molecules, which can be accessed by auxiliary analysis platforms such as the Biomedical and Healthcare Data Discovery Index Ecosystem (bioCADDIE) [
78] and Omics Discovery Index (OmicsDI) [
79]. Next, the DE molecules are enriched in the biological functional analyses to generate significant pathways and relation networks (target relation or interaction relation) with the support of some advanced repositories or tools for enrichment analyses, including gene ontology (GO), gene set enrichment analysis (GSEA), Reactome, Kyoto Encyclopedia of Genes and Genomes (KEGG), protein–protein interaction (PPI) databases, and use of the Kinase Target Network and Transcription Factor Network. In the subsequent analyses, the key molecules with significant optimal biological functional (BF) annotations, critical locations in the related networks, and targeted molecular pathways can be determined through various eye and vision studies.
4.2.3. Biomarker Targeting, Validation, and Clinical Detection
A prospective randomized controlled trial (PRCT) based on a small cohort in combination with special statistical approaches (e.g., Cox regression model, time-dependent receiver operating characteristic (ROC) curves, and Kaplan–Meier survival analyses) can be designed to estimate the prediction effectiveness, sensitivity, and specificity of potential biomarkers (the key output molecules from the previous step). Potential biomarkers with high sensitivity and specificity should be targeted as appropriate clinical indicators. However, the concrete mechanisms affecting retinal vascular diseases along with the stability, safety, and cost-effectiveness of candidates should be validated in vivo, in vitro, and in large-population PRCTs. Subsequently, some accurate, evidence-supported, stable, specific, and sensitive biomarkers with low costs and that can be detected using easily accessible detection methods can be applied and promoted in clinical screening, diagnosis, and prognosis determination.
4.3. Omics Biomarkers of Retinal Vascular Diseases
Recently, several omics studies have been conducted to identify potential biomarkers for understanding retinal vascular diseases. To deepen the practical value of the “omics-based biomarker discovery” approaches proposed in the previous section, we analyzed multiple omics data of retinal vascular diseases from specific data repositories. In the literature pertaining to omics data, some omics biomarkers for the screening, diagnosis, and prognosis of DR, AMD, RVO, and ROP were also determined.
4.3.1. Omics Biomarkers in DR
Multiple omics biomarkers of DR can be determined through screening, diagnosis, and prognosis methods. In addition, details regarding the accession number, type, originated specimen, and concrete clinical application value of omics data were extracted from the corresponding literature, as shown in
Supplementary Table S1.
Regarding DR screening, Guo et al. [
80] designed a case-control study on serum metabolomics of 69 pairs of type 2 diabetic patients (T2DM) with and without DR, where thiamine and tryptophan metabolic disorders, downregulated trehalose, and upregulated choline and indole derivatives were targeted as potential metabolic biomarkers. In addition, in a retrospective case-control epigenomic study, two serum epigenetic biomarkers, namely cg02873163 and cg11343894 in S100A13, were used to stratify patients who developed DR with a DM duration of <3 years and those without DR for more than 30 years after being diagnosed with DM [
81].
From the perspective of DR diagnosis, Skol et al. [
82] combined insightful genomics and transcriptomics data with patient clinical phenotyping information (no diabetes, no diabetic retinopathy, and proliferative diabetic retinopathy) using summary data-based Mendelian randomization analysis. Their study found that genetic variation and elevated expression of the folliculin (
FLCN) gene demonstrated a latent contribution to DR diagnosis and staging. In another study, integration analyses of metabolomic and lipidomic data revealed that four metabolic biomarkers, namely 2,4-dihydroxybutyric acid (DHBA), 3,4-DHBA, ribonic acid, and ribitol, and two lipid biomarkers, namely LDL cholesterol and triglycerides, were associated with different DR stages [
53]. Moreover, proteomic biomarkers such as IL-2/-5/-18/-13, TNF, and MMP-2/-3/-9 in tears have been utilized in non-invasive strategies for the diagnosis and staging of DR in recent research [
83].
In the case of DR prognosis, recent research has identified several diagnostic and prognostic miRNAs for recurrent vitreous hemorrhage in PDR patients through angiogenic miRNA profiling, including miRNA-19a and -27 as putative diagnostic vitreous biomarkers for PDR and elevated levels of miRNA-20a and -93 as prognostic biomarkers for recurrent hemorrhage incidence in PDR [
84]. DR, a long-term progressive disease, necessitates frequent follow-ups and intervention throughout a patient’s lifespan. Therefore, with the intention of developing a non-invasive and reliable monitoring method, a study proposed that biomarkers screened out through metabolic and lipid phenotype evaluations of tear fluids and sweat (such as taurine) may be a promising tool in the prognostic assessment and monitoring treatment efficacy of diabetic retinopathy [
85].
In addition to the above, researchers have emphasized the potential roles of metabolic biomarkers in the clinical diagnosis of DR. Li et al. [
86] summarized and tabulated diagnostic and severity-associated metabolic biomarkers from pre-DR, NPDR, mild PDR, and PDR samples over the past 10 years, including pyruvate, Asp, glycerol, cholesterol, Cer (d18:1/24:0), ChE 20:3, and many other metabolites. In addition, Jian et al. [
87] systemically reviewed the latest advances in metabolomics (from nuclear magnetic resonance (NMR) to liquid chromatography-mass spectrometry (LC-MS)) and the potential diagnostic metabolic biomarkers (12-hydroxyeicosatetraenoic acid (12-HETE), 2-piperidone, linoleic acid, nicotinuric acid, ornithine, and phenylacetylglutamine in human serum) from 39 original articles and then integrated the GWAS and metabolome data to predict molecular therapeutic targets (arginase 1, nitric oxide synthase 1, and phosphodiesterase). Furthermore, Hou et al. [
88] summarized the findings of nine metabolomics studies and identified several potential diagnostic biomarkers, including L-glutamine, L-lactic acid, pyruvic acid, acetic acid, L-glutamic acid, D-glucose, L-alanine, L-threonine, citrulline, L-lysine, and succinic acid, as well as the significant metabolic pathways involved in amino acid and energy metabolism.
Radiomics biomarkers are highly promising for DR diagnosis and prognosis. Afarid et al. [
89] analyzed the radiomics parameters in patients without clinical signs of DR compared to healthy individuals to determine several radiomic signs. Notably, enlarged foveal avascular zones (FAZs), reduced vascular density volumes in the macular area, and significant deviations of FAZ shape parameters (convexity and frequency domain irregularity) can be regarded as early screening identifiers of DR patients. Other Optical Coherence Tomography Angiography (OCTA) studies have also indicated that the size and shape of the FAZ may be correlated with a patient’s DR severity and grade [
90,
91]. Additionally, the retinal thickness measured by OCT B-scans [
92] and retinal leakage index dynamics on ultra-widefield fluorescein angiography images [
93] can be utilized to predict the response to intravitreal anti-VEGF treatment.
4.3.2. Omics Biomarkers in AMD
Multi-omics data in combination with clinical phenotyping data could offer a vast amount of reliable digital indicators to advance research on AMD disease. Several omics, proteomics, metabolomics, and epigenomics technologies have been integrated to screen for biomarkers associated with AMD, and the results revealed that some compounds associated with the oxidative stress pathway, complement system, and lipid metabolism pathway were regarded as the most appropriate candidates [
94]. In addition, the other omics biomarkers of AMD screening, diagnosis, and prognosis uncovered from omics research data from different databases are summarized in
Supplementary Table S2.
In the context of AMD screening, several serum transcriptomic indicators, including hsa-let-7a-5p, hsa-let-7d-5p, hsa-miR-23a-3p, hsa-miR-301a-3p, hsa-miR-361-5p, hsa-miR-27b-3p, hsa-miR-874-3p, and hsa-miR-19b-1-5p, were more highly expressed in the serum of patients with dry AMD (
n = 12) compared to wet AMD patients (
n = 14) and healthy controls (
n = 10), indicating that these dysregulated biomarkers may be potential diagnostic or screening tools for at-risk populations [
95]. In a clinical serum metabolomics study [
96], elevated serum concentrations of phosphatidylcholine, docoxahexaenoic acid, and eicosatetraenoic acid in choroidal neovascularization cases (CNV-AMD,
n = 20) and polypoidal choroidal vasculopathy cases (PCV,
n = 20) compared to healthy controls (
n = 20) showed putative roles in identifying AMD patients among vulnerable populations.
Regarding AMD diagnosis, epigenomic research [
97] has revealed that hypomethylation of the IL17RC promoter in AMD patients (
n = 202) compared to controls (
n = 96) can result in elevated concentrations of IL17RC proteins and messenger RNAs in the peripheral blood as well as in the affected retina and choroid. This suggests that the DNA methylation pattern and serum concentrations of IL17RC may be regarded as candidate biomarkers for the diagnosis of AMD. Another DNA methylation locus was recently reported, namely age-related maculopathy susceptibility 2 (ARMS2), and it has been shown that its CpG sites are significantly correlated with the risk SNP (rs10490924) genotype in these loci, thereby contributing to the development and progression of AMD [
98]. Schori et al. [
99] profiled proteins in the vitreous of AMD patients (dry AMD:
n = 6; neovascular AMD:
n = 10) using liquid chromatography coupled mass spectrometry. Specifically, upregulated cholinesterase (CHLE) in dry AMD patients and elevated ribonuclease (RNAS1) and serine carboxypeptidase (CPVL) in both types of AMD patients were identified as latent diagnostic biomarkers.
With regard to AMD prognosis and treatment response monitoring, the metabolites and proteins in systemic specimens may be suggestive of long-term benefits. Coronado et al. [
100] designed a pilot prospect study to estimate the protein profiles of AMD patients with good responses to anti-VEGF agents (
n = 5), poor responses to anti-VEGF agents (
n = 5), and of healthy controls (
n = 5). In their study, several dysregulated proteins were speculated to be potential biomarkers to distinguish AMD patients with different responses to anti-VEGF agents, and among these, LCN1 and the heavy chain component of IGHM were upregulated in the good response group and downregulated in the anti-VEGF poor response group. In addition, plasma kallikrein (KLKB1), kinase insert domain receptor (KDR), and vascular endothelial growth factor receptor 1 (VEGFR-1) were downregulated in the good response group and upregulated in the anti-VEGF poor response group.
The application of metabolomics has been highlighted in AMD research, and several reviews have summarized the potential metabolic biomarkers and pathways associated with AMD, emphasizing their roles in the construction of diagnostic or predictive models and new therapeutic targets for AMD [
101,
102,
103]. Hou et al. [
101] identified 108 dysregulated serum or plasma metabolites from the literature as potential candidate biomarkers and significantly enriched them in the adenosine, hypoxanthine, tyrosine, phenylalanine, creatine, citrate, carnitine, proline, and maltose pathways. Brown et al. [
102] also summarized a list of metabolites from systemic samples from other studies, including phosphatidylcholine, C3 carnitine, 2-ω-carboxyethyl pyrrole (CEP), and lysoPE, proposing that biomarkers in alternative biofluids such as tears, saliva, and vitreous may be of practical value in the diagnosis, staging, and prognosis determination of AMD due to the blood–retinal barrier. In another study, Deng et al. [
104] conducted untargeted metabolomics analyses of AMD samples and identified two metabolites that were differentially distributed between PCV and CNV. They found that the two metabolites, hyodeoxycholic acid and L-tryptophanamide, were differentially distributed between PCV and CNV, and the researchers investigated the genetic association of these metabolites with other metabolites in wet AMD. Liu et al. [
105] revealed distinct metabolic pathways in wet AMD, PCV, and pathological myopia, in which serum metabolomes were profiled using gas chromatography coupled with time-of-flight mass spectrometry (GC-TOFMS). In addition, multivariate statistical methods and data mining were used to interpret the macular neovascularization of the samples. Personalized pathway dysregulation score measurement using the Lilikoi package in R revealed that the pentose phosphate pathway and mitochondrial electron transport chain were the most important pathways in AMD. Furthermore, purine metabolism and glycolysis were identified as the major disturbed pathways in PCV, whereas altered thiamine metabolism and purine metabolism may contribute to PM phenotypes.
Serum metabolomics is a powerful tool for characterizing metabolic disturbances in macular neovascular disease. Li et al. [
86] summarized metabolomics-based clinical studies of AMD, and identified metabolic diagnostic and prognostic biomarkers/pathways in early, intermediate, and late AMD. Additionally, they reviewed some new therapeutic targets based on recent metabolomic findings, such as the pyruvate dehydrogenase kinase/lactate axis and findings related to taurine.
4.3.3. Omics Biomarkers in RVO and ROP
After DR, RVO is the second most common retinal vascular disorder and can be divided into central retinal vein occlusion (CRVO) and branch retinal vein occlusion (BRVO) [
3]. Proteomics research has revealed some significantly dysregulated molecules secondary to RVO. Yao et al. [
106] profiled the proteins in the aqueous humor of patients with cataract controls and BRVO and found that several downregulated proteins, including fibroblast growth factor 4 precursor, hepatoma-derived growth factor isoform a, and α-crystallin A, may serve as biomarkers for the development of macular edema in RVO. A recent study reported several upregulated proteins in the retina of a laser-induced BRVO model, including laminin subunit β-2, laminin subunit γ-1, lipocalin-7, nidogen-2, osteopontin, integrin-β1, isoform 2 of α-actinin-1, and talin-2. This study also suggested that proteins involved in focal adhesion signaling and major extracellular matrix remodeling processes are associated with neovascularization and macular edema following BRVO [
107]. In addition, other studies have illustrated that proteomic changes following aflibercept intervention in experimental CRVO [
108] and BRVO models [
109] can be applied to evaluate subjects’ responses to the intervention.
In their study, Xing et al. [
110] conducted metabolomics analyses of the aqueous humor in patients with RVO and identified differential profiles of metabolites between mild and severe macula edema (ME). In severe RVO-ME, compared with mild RVO-ME, the degradation and biosynthesis of valine, leucine, and isoleucine; histidine metabolism; beta-alanine metabolism; and pantothenate and coenzyme A biosynthesis significantly differed. Furthermore, receiver operating characteristic (ROC) curve analysis revealed that adenosine, threonic acid, pyruvic acid, and pyro-L-glutaminyl-l-glutamine could differentiate RVO-ME from controls with an area under the curve (AUC) of >0.813. Urocanic acid, diethanolamine, 8-butanoylneosolaniol, niacinamide, paraldehyde, phytosphingosine, 4-aminobutyraldehyde, dihydrolipoate, and 1-(beta-D-ribofuranosyl)-1,4-dihydronicotinamide had an AUC >0.848 for distinguishing mild RVO-ME from severe RVO-ME.
Even with the best neonatal care and management strategies, ROP remains the major cause of blindness and visual disability in infancy [
111]. Fortunately, advanced omics technology has provided new possibilities for the screening, diagnosis, prognosis, and management of ROP. For instance, Good et al. [
112] utilized genomics to identify genetic factors to screen for potential treatment targets and explain the progression of ROP development. Furthermore, proteomics will serve as useful determinants of disease states and stages as well as diagnostic and prognostic tools for ROP. In another study, Danielsson et al. [
113] profiled the serum proteins of 14 extremely preterm infants at postnatal days 1, 7, 14, and 28 and at postmenstrual ages of 32, 36, and 40 weeks, demonstrating that 11 proteins associated with ROP were potential biomarkers for predicting ROP. These eleven proteins were significantly enriched in angiogenesis, neurogenesis, osteogenesis, immune, and lipid metabolism functions and included AGER, ANGPT1, amyloid-beta precursor protein (APP), CD40 ligand (CD40LG), SLAM family member 5 (CD84), GDF2, heparin-binding EGF-like growth factor (HBEGF), macrophage metalloelastase (MMP12), peptidyl-prolyl cis–trans isomerase B (PPIB), plasminogen activator inhibitor 1 (SERPINE1), and thrombomodulin (THBD). Another proteomic study screened a total of 498 dysregulated proteins during vessel regression (PN12) and 345 dysregulated proteins during neovascularization (PN17) in the retinas of OIR pups that were involved in angiogenesis-related processes such as matrix remodeling, cell migration, adhesion, and proliferation. In addition, the mRNA expression in a human retinal endothelial cell (HREC) ROP model confirmed 56/69 neovascular-specific proteins, with 23 showing the same expression trends as OIR neovascular retinas. Finally, RNAi and transfection overexpression studies demonstrated that VASP and ECH1, which showed the highest levels in hypoxic HRECs, promoted human umbilical vein (HUVEC) and HREC cell proliferation, while SNX1 and CD109, which showed the lowest levels, inhibited their proliferation [
114].
Zhou et al. [
115] identified distinct profiles of plasma metabolites in patients with treatment-requiring ROP (TR-ROP) and controls. They also conducted metabolomic analyses in a murine ROP model and indicated that “protein digestion and absorption” and “aminoacyl-tRNA biosynthesis” were the most enriched pathways of the altered metabolites. These results demonstrated that the metabolomic profiles changed in the plasma of TR-ROP patients, and the altered metabolites could serve as potential biomarkers for the diagnosis and prognosis of TR-ROP patients. These findings illustrate that these proteins may act as potential biomarkers of vascular-related disorders. Moreover, a targeted blood metabolomic study reported dysregulated metabolites, including glycine, glutamate, leucine, serine, piperidine, valine, tryptophan, citrulline, malonyl carnitine (C3DC), and homocysteine, between 41 ROP and 40 non-ROP infants. In particular, high blood levels of C3DC and glycine can be promising screening biomarkers for ROP [
116].
4.3.4. Opportunities and Challenges That Coexist in Multi-Omics Integration Approaches Related to Biomarker Discoveries
Although a large number of omics biomarkers for the screening, diagnosis, and prognosis of retinal vascular diseases have been reported, there is still a lack of suitable candidates with strong evidence and stability for further validation and clinical detection, as these involve processes that require considerable manpower, financial costs, and time. Integration analysis of multi-omics data can optimize the biomarker discovery process by enhancing the evidence chain, minimizing the investment in manpower, materials, and time, and quantifying its stability in specimens.
The integration of distinct omics data and phenotypic information from previous studies should be a highly cost-effective method with which to discover candidate clinical indicators by enlarging the sample size, enhancing the statistical power, and shortening the validation process. In addition, the application of multi-omics methods in retinal vascular diseases is helpful for ophthalmic diseases as well as for systemic vascular diseases. As the only visible blood vessel in the human body, the early detection of retinal vascular lesions can indicate other systemic vascular diseases, and its lesions may be a representation of systemic vascular diseases. Through certain approaches using ophthalmological instruments, suspicious lesions can be screened for, and the root cause of the disease can be determined. In addition, the related changes in epigenetics, proteomics, and metabolomics can be unearthed through multi-omics methods. Finally, the metabolome may be a more accessible biomarker discovery means, as it can provide precise phenotype outcomes based on patient biology data.
The discovery of multi-omics biomarkers remains in its infancy, and great challenges exist in the associated practical application processes. The main challenges lie in the establishment or acquisition of detailed long-term clinical and phenotypic information datasets for further confirmatory studies, and this inevitably requires cooperation among multiple centers. Moreover, complex data integration requires exquisite programming technology and a solid foundation of clinical, genetic, and biological knowledge to discriminate key molecules from large sources of information. Therefore, the maturation of multi-omics biomarker discovery methods still requires a long-term development period.
This entry is adapted from the peer-reviewed paper 10.3390/cells12010103