Visual illusions fascinate humans, in large part because we realize how such experiences disconnect how we perceive the world from reality. The discovery that other animals also experience some of these illusions has provided a compelling comparative story about the role that perception plays in sometimes misrepresenting the nature of the real world. What has also become apparent from comparative studies is that not all animals experience illusions the same
way, and sometimes the same individual may not experience some illusions while experiencing others. A survey of the results of 14 experiments from 10 different studies with two monkey species that performed numerous illusion tasks reinforces the idea that individual differences are a rich source of information for better understanding any universal principles of visual perception. Future research should focus more effort towards understanding the causes and effects (on other behaviors) of experiencing (or not experiencing) these illusions.
1. Introduction
Perceptual illusions fascinate humans. They are omnipresent in the modern world. Visual illusions are popular on social media platforms, where there are yearly articles on the best illusions from that year [
1], and where one finds endless debate about what people think they may see, and why it is so fascinating (and frustrating) that others do not see the same thing (e.g., is the dress blue or gold? [
2]). Beyond entertainment, illusions are often featured in scientific settings for the public (for example, the Fernbank Museum of Natural History in Atlanta has featured an exhibit on sensory experience and perceptual illusions for many years), and they are even a part of the educational system for many children.
Perceptual illusions serve a purpose beyond entertaining people as as they also help researchers understand the constraints of our perceptual experience, and they inform people about how other cognitive mechanisms interact with sensation and perception to generate such experiences [
6]. Although perceptual illusions are “errors in the machine,” they also are tools for understanding how that machine works more broadly, in the same way that decision-making errors and biases inform people about how researchers frame decisions and calculate the value of options. The ancient Greeks considered the causes and implications of perceptual illusions (e.g., Aristotle and the waterfall effect), and the first scientific description of an illusion was carried out by Necker [
7], followed by numerous other descriptions of illusions shortly after [
8,
9]; for a review, see [
10]. Central to many of these early experiments was an appreciation of individual differences in how people experienced illusions, although formal assessments of what accounted for those differences often was not undertaken.
Given this widespread interest in perceptual illusions among humans, combined with the fascination about the minds of other animals, it is no surprise that studying illusory experiences across species has been a longstanding area of focus in comparative psychology and ethology [
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25,
26]. Although studies of illusory experiences have been conducted with many species, we focus our attention on nonhuman primates, simply for the reason that we know more about those species than others, and because nonhuman primates have been among the most extensively tested species. We highlight specific studies from our laboratory that included tests with rhesus monkeys (
Macaca mulatta) and capuchin monkeys (
Sapajus apella), sometimes in comparison to nearly identical tests given to human adults and human children. The goal in focusing on that work is not to downplay or ignore the many other creative and informative studies of other primates or non-primate species [
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25,
26], but to show that even in a restricted sample, some important points emerge about the need to assess and try to account for individual differences in these illusory experiences. Those differences are present and are often acknowledged and reported in empirical studies, even if there is little attempt made to explain why those differences exist. Some of the earliest papers in comparative psychology made this clear. For example, when chicks were given four tests of different illusions, it was noted in the abstract of that report that marked individual differences in susceptibility were shown for all four of the illusions [
11]. However, there was no indication of what accounted for these differences.
Such individual differences likely offer substantial insight into the nature of perceptual processing mechanisms, not for their universality of operation, but for the way in which those mechanisms interact with variables such as species membership, age, sex, foraging ecology, attentiveness, and the prioritization of so-called top-down versus bottom-up processing of stimulus properties. Although there is still excellent value in charting what new species may be susceptible to different illusory experiences, so as to broaden the phylogenetic map of such experiences, there is equal value in trying to understand why some individuals experience illusions whereas others do not. Additionally, there is great value in understanding why even the same individual may be susceptible to some illusory experiences but not others.
2. Visual Illusions Research
Data were drawn from published papers from the Language Research Center at Georgia State University that included rhesus monkeys and/or capuchin monkeys, and that specifically focused on one or more geometric visual illusions. We included any papers that had one or more primate subjects in them who also were subjects in at least three other experiments from our laboratory that assessed perceptual illusion susceptibility. Any included papers had to report the specific performances of each subject with regards to the illusory outcome (e.g., misperceiving item size or relative quantity), so that we could assign a positive or negative value to that subject for that illusory experience. We chose to classify each subject as either “showing the illusion” or not, rather than trying to quantify the magnitude of the illusion, because the latter effort would have excluded a large number of these papers, and we wanted a larger sample size. This resulted in a total of 10 studies and 14 individual experiments, each of which is described in the linked paper (along with examples of these visual illusions). Across all experiments, a total of 8 rhesus monkeys and 10 capuchin monkeys met criterion for inclusion in this summary by participating in at least four experiments.
Table 1 shows the results of this qualitative survey. Not a single monkey was susceptible to all illusory stimuli to which they were exposed. At the other extreme, one monkey (Hank) showed susceptibility in only one of the nine experiments in which he participated. All other monkeys showed susceptibility to some but not all illusions, and many of them showed susceptibility about half of the time. For those studies in which two experiments were conducted, providing a conceptual replication within-study, there were also changes in susceptibility for the monkeys. In Agrillo et al. [
43], only 1 of 13 monkeys replicated their susceptibility in both experiments. In Agrillo et al. [
44], only two of six monkeys showed the illusory effect in both experiments. In Parrish et al. [
45], only one monkey (Gale) showed the illusory effect in both experiments, and in Parrish and Beran [
52], three of five monkeys replicated the illusory outcome. These qualitative results make clear the variability across illusions within this sample of monkeys, and also the variability within individuals not only for different illusions, but also for the same illusion tested at different times.
Table 1. Susceptibility of each monkey in each experiment in terms of experiencing the illusion in the same direction as for humans.
| Monkey |
Agrillo
et al.,
2014a |
Agrillo
et al., 2014b
Exp 2 |
Agrillo et al., 2014b Exp 3 |
Agrillo
et al.,
2015
Exp 1 |
Agrillo
et al.,
2015
Exp 4 |
Parrish
et al.,
2015
Exp 1 |
Parrish
et al.,
2015
Exp 2 |
Parrish
et al.,
2016
Exp 1 |
Parrish
et al.,
2019 |
Agrillo
et al.,
2019 |
Parrish
et al.,
2020 |
Parrish & Beran 2021
Exp 1 |
Parrish & Beran 2021
Exp 2 |
McKeon
et al.,
2022 |
| Macaques |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Chewie |
|
YES |
NO |
|
|
NO |
YES |
|
YES |
No |
|
YES |
NO |
NO |
| Gale |
|
YES |
NO |
YES |
NO |
YES |
YES |
|
|
|
|
|
|
|
| Han |
|
YES |
YES |
|
|
NO |
YES |
|
YES |
No |
|
NO ^ |
NO ^ |
NO |
| Hank |
|
NO |
NO |
YES |
NO |
NO |
NO ^ |
|
NO |
NO |
|
|
|
NO |
| Lou |
YES |
|
|
YES |
YES |
NO |
YES |
|
NO |
NO |
|
YES |
YES |
NO |
| Luke |
YES |
NO |
NO |
YES |
YES |
|
|
|
NO |
NO |
|
|
|
NO |
| Murph |
YES |
NO |
YES |
NO |
YES |
NO |
YES |
|
NO |
NO |
|
YES |
YES |
NO |
| Obi |
|
|
|
NO |
NO |
NO |
YES |
|
YES |
NO |
|
YES |
YES |
NO |
| Capuchins |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Gonzo * |
|
|
|
|
|
NO |
YES |
YES |
NO |
NO |
|
|
|
|
| Gretel * |
|
|
|
|
|
NO |
YES |
YES |
NO |
NO |
|
|
|
|
| Griffin |
|
YES |
NO |
|
|
NO |
YES |
|
NO |
NO |
YES |
|
|
|
| Liam |
|
NO |
NO |
|
|
NO ^ |
YES |
|
NO |
NO |
YES |
|
|
|
| Logan |
|
YES |
NO ^ |
|
|
NO ^ |
YES |
|
YES |
NO |
NO |
|
|
|
| Mason |
|
|
|
|
|
NO ^ |
YES |
NO |
NO |
NO |
|
|
|
|
| Nala * |
|
NO |
NO |
|
|
NO ^ |
NO ^ |
|
NO |
NO |
YES |
|
|
|
| Nkima |
|
YES |
NO |
|
|
NO |
YES |
|
NO ^ |
NO |
NO |
|
|
|
| Widget * |
|
NO |
NO |
|
|
|
|
|
YES |
NO |
YES |
|
|
|
| Wren * |
|
YES |
NO |
|
|
NO |
YES |
|
YES |
NO |
YES |
|
|
|
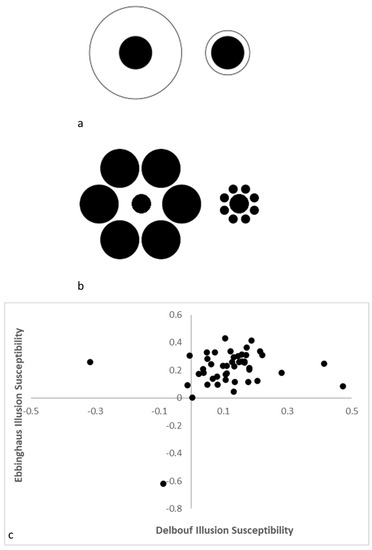
In an unpublished study from our laboratory conducted with adult humans (college students), we found a similar pattern. We presented 46 participants with the Delboeuf illusion (hereafter DBI; [
55]) and the Ebbinghaus illusion (hereafter EBI; [
56]) in the context of having those participants adjust an onscreen dot until they thought it matched the size of a comparison stimulus presented in an illusory context (
Figure 2a,b). Although most participants had a tendency to overestimate the size of the dot presented in a small surrounding circle for the DBI and the size of the dot centered among small surrounding circles in the EBI, there were individuals who did not show this typical susceptibility. The relation of the magnitude of DBI and EBI susceptibility in this sample was nonsignificant, r(44) = 0.23,
p = 0.12, and it accounted for only 5.3% of the variance in the sample (
Figure 2c). As with the monkeys, humans are variable in their susceptibility to closely related illusory experiences, even within the same task given at the same time to the same people.
Figure 2. The Delboeuf (a) and Ebbinghaus (b) illusions in which the central dots surrounded by a smaller perimeter ring or a series of smaller other dots (i.e., the right stimuli in each image) are overestimated relative to when the surrounding ring or series of circles are large. The relative susceptibility to each of these illusions in a sample of 46 adult humans where positive values indicate the typical illusory experience (c).
3. Discussion
The take-home message of this brief survey of a sample of primates, given a moderate range of perceptual illusory experiences, is that there was a “consistent inconsistency” in their susceptibility to such illusions. Across these studies, rarely did any specific primate experience most or all illusions. Rather, the majority of these monkeys sometimes responded as if they experienced a given illusion, although across multiple tests of the same or similar illusions, there was variability at the level of the individual. Additionally, this pattern matched that of an unpublished sample of human participants given two well-known illusions, the Delboeuf illusion and the Ebbinghaus illusion, even though those two illusions were presented within the same task, using the same computerized apparatus. It also matched the consistent reports of large-scale studies with humans in which there were large individual differences in susceptibility [
27,
37,
38,
39,
40].
There are a number of reasons why illusory experiences in nonhuman animals (and humans) may be consistently inconsistent. One strong candidate for accounting for difference across illusions (and studies) is methodological variation. How individuals are trained to perform tasks, how stimuli are created and presented, and what form responses take in tasks all could affect inconsistencies in experiencing illusions across studies (see [
57,
58,
59]). Recognizing the relative priority of global versus local modes of processing visual stimuli in different species and individuals also can account for variation in illusion susceptibility (e.g., [
60,
61,
62,
63,
64,
65,
66]). Differences in psychophysical discriminability capacities also are relevant, as are factors such as discrimination thresholds (acuity in size and numerosity discriminations), motivation to engage with tasks, previous testing histories that may conflict with or support a more illusory experience in newer tasks, and even factors such as how quickly or slowly individuals (and species) respond in tasks. It is likely that all of these factors and others play a role in the species differences and individual differences that are evident in the literature. In some cases, relevant factors could be controlled in ways that better isolate illusion susceptibility, although it is important to also assess illusory experiences in more naturalistic choice and decision setting (e.g., chimpanzees falling prey to the DBI when being allowed to look at and choose food items to immediately consume [
67]).
Although researchers cannot (yet) adequately explain the individual differences seen in monkeys, or other animals, this is an attainable goal, and will allow a productive focus of future research. There is a richness in these individual differences for better understanding the cause(s) of illusory experiences, and more broadly for understanding the different phenomenal worlds of different species, different individuals within a species, and the same individuals tested at different times. Although there are certainly major differences in how some species may perceive and process sensory information relative to other species (e.g., the bat versus the reptile), there may also be substantial intraspecific differences that affect other behavioral and cognitive processes. As just one example, based on some of the studies shown in Table 1, differences in susceptibility to numerical or quantitative illusions could impact foraging behavior, social approach or avoidance behavior, and other natural behaviors that rely on perceiving and representing magnitude information such as quantity or number. The same is true for size-based judgments, which may be affected by those individuals who are more susceptible to illusory experiences.
Most likely, those individual differences do not lead to substantial fitness benefits to those who are less susceptible to illusions, or else there would have been selection against such susceptibility. However, this does not mean that illusory experiences are not detrimental. One could predict that illusory experiences may generate more decision errors in some kinds of choice tasks, or greater likelihood of errors in different kinds of memory tests, and especially those for which one must remember the actual attributes of a stimuli that could be misrepresented due to the illusory experience. As such, documenting and accommodating individual differences in illusory experience could aid in accounting for variance in animal cognition and behavior that has otherwise not been accounted for through the manipulation of other factors.
This entry is adapted from the peer-reviewed paper 10.3390/ani13010022