Aflatoxins are natural toxicants produced mainly by species of the Aspergillus genus, which contaminate virtually all feeds and foods. Apart from their deleterious health effects on humans and animals, they can be secreted unmodified or carried over into the milk of lactating females, thereby posing health risks to suckling babies. Aflatoxin M1 (AFM1) is the major and most toxic aflatoxin type after aflatoxin B1 (AFB1). It contaminates human breast milk upon direct ingestion from dairy products or by carry-over from the parent molecule (AFB1), which is hydroxylated in the liver and possibly in the mammary glands by cytochrome oxidase enzymes and then excreted into breast milk as AFM1 during lactation via the mammary alveolar epithelial cells.
- human breast milk
- aflatoxin M1
- child growth and development
- health risk
- influx/efflux transport
1. Routes of Breast Milk Contamination with AFM1
1.1. Diet
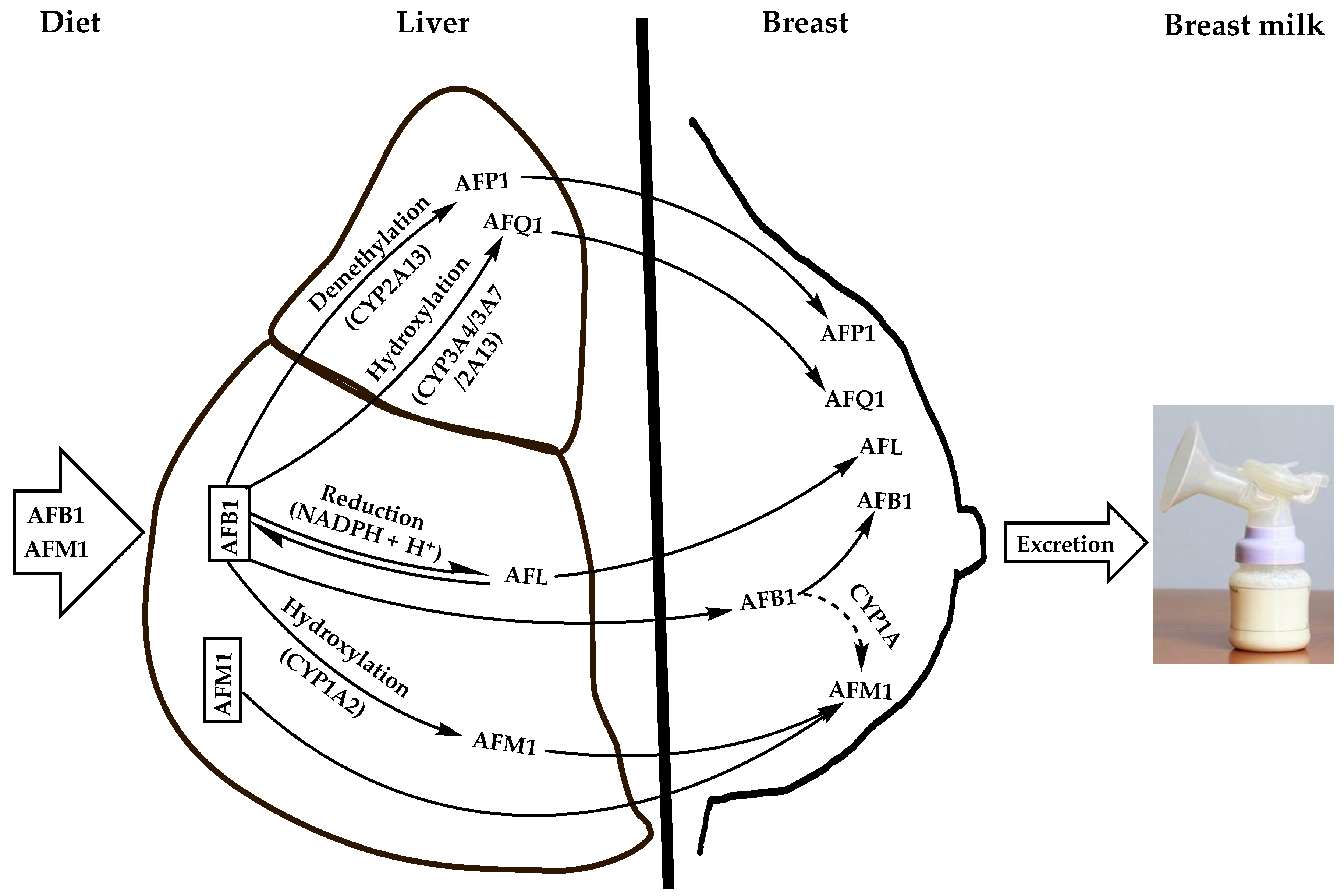
1.2. Feed as an Indirect Source of Human Breast Milk Contamination with AFM1
2. Adverse Health Effects of AFM1 on Infants and Young Children
2.1. Growth Impairment
2.2. Other AFM1-Related Health Issues
This entry is adapted from the peer-reviewed paper 10.3390/ijerph192416792
References
- Deng, J.; Zhao, L.; Zhang, N.Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.S.; Sun, L.H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. 2018, 778, 79–89.
- Bertero, A.; Augustyniak, J.; Buzanska, L.; Caloni, F. Species-specific models in toxicology: In vitro epithelial barriers. Environ. Toxicol. Pharmacol. 2019, 70, 103203.
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423.
- Yiannikouris, A.; Jouany, J.-P. Mycotoxins in feeds and their fate in animals: A review. Anim. Res. 2002, 51, 81–99.
- Ghadiri, S.; Spalenza, V.; Dellafiora, L.; Badino, P.; Barbarossa, A.; Dall’Asta, C.; Nebbia, C.; Girolami, F. Modulation of aflatoxin B1 cytotoxicity and aflatoxin M1 synthesis by natural antioxidants in a bovine mammary epithelial cell line. Toxicol. Vitr. 2019, 57, 174–183.
- Smith, J.W.; Groopman, J.D. Aflatoxins. In Encyclopedia of Cancer, 3rd ed.; Boffetta, P., Hainaut, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 30–43.
- Loveland, P.M.; Wilcox, J.S.; Pawlowski, N.E.; Bailey, G.S. Metabolism and DNA binding of aflatoxicol and aflatoxin B1 in vivo and in isolated hepatocytes from rainbow trout (Salmo gairdneri). Carcinogenesis 1987, 8, 1065–1070.
- Bailey, G.S.; Loveland, P.M.; Pereira, C.; Pierce, D.; Hendricks, J.D.; Groopman, J.D. Quantitative carcinogenesis and dosimetry in rainbow trout for aflatoxin B1 and aflatoxicol, two aflatoxins that form the same DNA adduct. Mutat. Res. 1994, 313, 25–38.
- Carvajal-Moreno, M.; Vargas-Ortiz, M.; Hernández-Camarillo, E.; Ruiz-Velasco, S.; Rojo-Callejas, F. Presence of unreported carcinogens, Aflatoxins and their hydroxylated metabolites, in industrialized Oaxaca cheese from Mexico City. Food Chem. Toxicol. 2019, 124, 128–138.
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7.
- Massey, T.E.; Guindon, K.A. Aflatoxins. In Encyclopedia of Cancer; Schwab, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; p. 101.
- Dohnal, V.; Wu, Q.; Kuča, K. Metabolism of aflatoxins: Key enzymes and interindividual as well as interspecies differences. Arch. Toxicol. 2014, 88, 1635–1644.
- Mykkänen, H.; Zhu, H.; Salminen, E.; Juvonen, R.O.; Ling, W.; Ma, J.; Polychronaki, N.; Kemiläinen, H.; Mykkänen, O.; Salminen, S.; et al. Fecal and urinary excretion of aflatoxin B1 metabolites (AFQ1, AFM1 and AFB-N7-guanine) in young Chinese males. Int. J. Cancer 2005, 115, 879–884.
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological properties and their involvement in cancer development. Toxins 2018, 10, 214.
- Bailey, G.S.; Dashwood, R.; Loveland, P.M.; Pereira, C.; Hendricks, J.D. Molecular dosimetry in fish: Quantitative target organ DNA adduction and hepatocarcinogenicity for four aflatoxins by two exposure routes in rainbow trout. Mutat. Res. 1998, 399, 233–244.
- Karabulut, S.; Paytakov, G.; Leszczynski, J. Reduction of aflatoxin B1 to aflatoxicol: A comprehensive DFT study provides clues to its toxicity. J. Sci. Food Agric. 2014, 94, 3134–3140.
- Caruso, M.; Mariotti, A.; Zizzadoro, C.; Zaghini, A.; Ormas, P.; Altafini, A.; Belloli, C. A clonal cell line (BME-UV1) as a possible model to study bovine mammary epithelial metabolism: Metabolism and cytotoxicity of aflatoxin B1. Toxicon 2009, 53, 400–408.
- IARC. Monographs on the Evaluation of Carcinogenic Risks to Humans: Chemical Agents and Related Occupations; A Review of Human Carcinogens, 100 F; IARC: Lyon, France, 2012.
- Polychronaki, N.; Turner, P.C.; Mykkänen, H.; Gong, Y.Y.; Amra, H.; Abdel-Wahhab, M.; El-Nezami, H. Determinants of aflatoxin M1 in breast milk in a selected group of Egyptian mothers. Food Addit. Contam. 2006, 23, 700–708.
- Carvajal, M.; Rojo, F.; Méndez, I.; Bolaños, A. Aflatoxin B1 and its interconverting metabolite aflatoxicol in milk: The situation in Mexico. Food Addit. Contam. 2003, 20, 1077–1086.
- Warth, B.; Braun, D.; Ezekiel, C.N.; Turner, P.C.; Degen, G.H.; Marko, D. Biomonitoring of Mycotoxins in Human Breast Milk: Current State and Future Perspectives. Chem. Res. Toxicol. 2016, 29, 1087–1097.
- Braun, D.; Abia, W.A.; Šarkanj, B.; Sulyok, M.; Waldhoer, T.; Erber, A.C.; Krska, R.; Turner, P.C.; Marko, D.; Ezekiel, C.N.; et al. Mycotoxin-mixture assessment in mother-infant pairs in Nigeria: From mothers’ meal to infants’ urine. Chemosphere 2022, 287, 132226.
- García-Lino, A.M.; Álvarez-Fernández, I.; Blanco-Paniagua, E.; Merino, G.; Álvarez, A.I. Transporters in the Mammary Gland—Contribution to Presence of Nutrients and Drugs into Milk. Nutrients 2019, 11, 2372.
- Williams, J.A.; Phillips, D.H. Mammary Expression of Xenobiotic Metabolizing Enzymes and Their Potential Role in Breast Cancer. Cancer Res. 2000, 60, 4667–4677.
- Williams, J.A.; Stone, E.M.; Millar, B.C.; Gusterson, B.A.; Grover, P.L.; Phillips, D.H. Determination of the enzymes responsible for activation of the heterocyclic amine 2-amino-3-methylimidazoquinoline in the human breast. Pharmacogenetics 1998, 8, 519–528.
- Kuilman, M.E.M.; Maas, R.F.M.; Fink-Gremmels, J. Cytochrome P450-mediated metabolism and cytotoxicity of aflatoxin B1 in bovine hepatocytes. Toxicol. Vitr. 2000, 14, 321–327.
- Héraud, F.; Barraj, L.M.; Moy, G.G. GEMS/Food Consumption Cluster Diets. In Total Diet Studies; Moy, G.G., Vannoort, R.W., Eds.; Springer: New York, NY, USA, 2013; pp. 427–434.
- Britzi, M.; Friedman, S.; Miron, J.; Solomon, R.; Cuneah, O.; Shimshoni, J.A.; Soback, S.; Ashkenazi, R.; Armer, S.; Shlosberg, A. Carry-over of aflatoxin B1 to aflatoxin M1 in high yielding Israeli cows in mid- and late-lactation. Toxins 2013, 5, 173–183.
- Ayar, A.; Sert, D.; Con, A.H. A study on the occurrence of aflatoxin in raw milk due to feeds. J. Food Safety 2007, 27, 199–207.
- Costamagna, D.; Gaggiotti, M.; Chiericatti, C.A.; Costabel, L.; Audero, G.M.L.; Taverna, M.; Signorini, M.L. Quantification of aflatoxin M1 carry-over rate from feed to soft cheese. Toxicol. Rep. 2019, 6, 782–787.
- Veldman, A.; Meijs, J.A.C.; Borggreve, G.J.; Heeres-van der Tol, J.J. Carry-over of aflatoxin from cows’ food to milk. Anim. Sci. 1992, 55, 163–168.
- Phillips, E.; Turner, P.C.; Ngure, F.M.; Kassim, N.; Makule, E.; Smith, L.E.; Nelson, R.J.; Stoltzfus, R.J. Ethical considerations in the design and conduct of a cluster-randomised mycotoxin mitigation trial in Tanzania. World Mycotoxin J. 2022, 15, 213–222.
- Phillips, E.; Ngure, F.; Smith, L.E.; Makule, E.; Turner, P.C.; Nelson, R.; Kimanya, M.; Stoltzfus, R.; Kassim, N. Protocol for the trial to establish a causal linkage between mycotoxin exposure and child stunting: A cluster randomized trial. BMC Public Health 2020, 20, 598.
- Phillips, E.; Turner, P.C.; Kassim, N.; Makule, E.; Nelson, R.; Ngure, F.; Smith, L.; Stoltzfus, R. Ethical considerations of the trial to establish a causal linkage between mycotoxin exposure and child stunting. Curr. Dev. Nutr. 2021, 5 (Suppl. 2), 678.
- El-Nezami, H.S.; Nicoletti, G.; Neal, G.E.; Donohue, D.C.; Ahokas, J.T. Aflatoxin M1 in human breast milk samples from Victoria, Australia and Thailand. Food Chem. Toxicol. 1995, 33, 173–179.
- Adejumo, O.; Atanda, O.O.; Raiola, A.; Somorin, Y.; Bandyopadhyay, R.; Ritieni, A. Correlation between aflatoxin M1 content of breast milk, dietary exposure to aflatoxin B1 and socioeconomic status of lactating mothers in Ogun State, Nigeria. Food Chem. Toxicol. 2013, 56, 171–177.
- Elaridi, J.; Bassil, M.; Kharma, J.A.; Daou, F.; Hassan, H.F. Analysis of Aflatoxin M1 in Breast Milk and Its Association with Nutritional and Socioeconomic Status of Lactating Mothers in Lebanon. J. Food Prot. 2017, 80, 1737–1741.
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24.
- El-Tras, W.F.; El-Kady, N.N.; Tayel, A.A. Infants exposure to aflatoxin M1 as a novel foodborne zoonosis. Food Chem. Toxicol. 2011, 49, 2816–2819.
- Benkerroum, N. Mycotoxins in dairy products: A review. Int. Dairy J. 2016, 62, 63–75.
- Battacone, G.; Nudda, A.; Palomba, M.; Pascale, M.; Nicolussi, P.; Pulina, G. Transfer of aflatoxin B1 from feed to milk and from milk to curd and whey in dairy sheep fed artificially contaminated concentrates. J. Dairy Sci. 2005, 88, 3063–3069.
- van der Fels-Klerx, H.J.; Camenzuli, L. Effects of Milk Yield, Feed Composition, and Feed Contamination with Aflatoxin B1 on the Aflatoxin M1 Concentration in Dairy Cows’ Milk Investigated Using Monte Carlo Simulation Modelling. Toxins 2016, 8, 290.
- Frobish, R.A.; Bradley, B.D.; Wagner, D.D.; Long-Bradley, P.E.; Hairston, H. Aflatoxin Residues in Milk of Dairy Cows after Ingestion of Naturally Contaminated Grain. J. Food Prot. 1986, 49, 781–785.
- Masoero, F.; Gallo, A.; Moschini, M.; Piva1, G.; Diaz, D. Carryover of aflatoxin from feed to milk in dairy cows with low or high somatic cell counts. Animal 2007, 9, 1344–1350.
- van Eijkeren, J.C.; Bakker, M.I.; Zeilmaker, M.J. A simple steady-state model for carry-over of aflatoxins from feed to cow’s milk. Food Addit. Contam. 2006, 23, 833–838.
- Zarba, A.; Wild, C.P.; Hall, A.J.; Montesano, R.; Hudson, G.J.; Groopman, J.D. Aflatoxin M1 in human breast milk from The Gambia, west Africa, quantified by combined monoclonal antibody immunoaffinity chromatography and HPLC. Carcinogenesis 1992, 13, 891–894.
- Khlangwiset, P.; Shephard, G.S.; Wu, F. Aflatoxins and growth impairment: A review. Crit. Rev. Toxicol. 2011, 41, 740–755.
- da Silva, J.V.; de Oliveira, C.A.; Ramalho, L.N. Effects of Prenatal Exposure to Aflatoxin B1: A Review. Molecules 2021, 26, 7312.
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Humphrey, J.H.; Stoltzfus, R.J. Aflatoxin exposure during pregnancy, maternal anemia, and adverse birth outcomes. Am. J. Trop. Med. Hyg. 2017, 96, 770–776.
- Abdulrazzaq, Y.M.; Osman, N.; Yousif, Z.M.; Trad, O. Morbidity in neonates of mothers who have ingested aflatoxins. Ann. Trop. Paediatr. 2004, 24, 145–151.
- Adhikari, M.; Gita, R.; Berjak, P. Aflatoxin, kwashiorkor, and morbidity. Nat. Toxins 1994, 2, 1–3.
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; de Onis, M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R.; et al. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451.
- Ortiz, J.; Jacxsens, L.; Astudillo, G.; Ballesteros, A.; Donoso, S.; Huybregts, L.; De Meulenaer, B. Multiple mycotoxin exposure of infants and young children via breastfeeding and complementary/weaning foods consumption in Ecuadorian highlands. Food Chem. Toxicol. 2018, 118, 541–548.
- Sherif, S.O.; Salama, E.E.; Abdel-Wahhab, M.A. Mycotoxins and child health: The need for health risk assessment. Int. J. Hyg. Environ. Health 2009, 212, 347–368.
- Hulin, M.; Bemrah, N.; Nougadère, A.; Volatier, J.L.; Sirot, V.; Leblanc, J.C. Assessment of infant exposure to food chemicals: The French Total Diet Study design. Food Addit. Contam. Part A 2014, 31, 1226–1239.
- Lauer, J.M.; Duggan, C.P.; Ausman, L.M.; Griffiths, J.K.; Webb, P.; Wang, J.S.; Xue, K.S.; Agaba, E.; Nshakira, N.; Ghosh, S. Maternal aflatoxin exposure during pregnancy and adverse birth outcomes in Uganda. Matern. Child Nutr. 2019, 15, e12701.
- Turner, P.C.; Collinson, A.C.; Cheung, Y.B.; Gong, Y.Y.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Aflatoxin exposure in utero causes growth faltering in Gambian infants. Int. J. Epidemiol. 2007, 36, 1119–1125.
- JME (UNICEF/WHO/World Bank). Levels and Trends in Child Malnutrition: Key Fndings of the 2021 Edition of the Joint Child Malnutrition Estimates; World Health Organization: Geneva, Switzerland, 2021.
- Boni, S.B.; Beed, F.; Kimanya, M.E.; Koyano, E.; Mponda, O.; Mamiro, D.; Kaoneka, B.; Bandyopadhyay, R.; Korie, S.; Mahuku, G. Aflatoxin contamination in Tanzania: Quantifying the problem in maize and groundnuts from rural households. World Mycotoxin J. 2021, 14, 553–564.
- Ministry of Health, C.D. Gender, Elderly and Children (MoHCDGEC) ; Ministry of Health (MoH) ; National Bureau of Statistics (NBS); Office of the Chief Government Statistician (OCGS); ICF; Tanzania Demographic and Health Survey and Malaria Indicator Survey (TDHS-MIS) 2015–2016. Dar es Salaam Rockville: MoHCDGEC, MoH, NBS, OCGS, and ICF. 2016, p. 591. Available online: https://dhsprogram.com/pubs/pdf/fr321/fr321.pdf (accessed on 12 June 2022).
- UNICEF. The Conceptual Framework of Under-Nutrition. 2022. Available online: https://forum.susana.org/244-theme-1-making-the-link-in-theory-and-practice-where-do-we-stand/17576-the-conceptual-framework-of-under-nutrition (accessed on 12 June 2022).
- Gong, Y.Y.; Cardwell, K.; Hounsa, A.; Egal, S.; Turner, P.C.; Hall, A.J.; Wild, C.P. Dietary aflatoxin exposure and impaired growth in young children from Benin and Togo: Cross sectional study. BMJ 2002, 325, 20–21.
- Maxwell, S.M.; Familusi, J.B.; Sodeinde, O.; Chan, M.C.; Hendrickse, R.G. Detection of naphthols and aflatoxins in Nigerian cord blood. Ann. Trop. Paediatr. 1994, 14, 3–5.
- Magoha, H.; Kimanya, M.; De Meulenaer, B.; Roberfroid, D.; Lachat, C.; Kolsteren, P. Risk of dietary exposure to aflatoxins and fumonisins in infants less than 6 months of age in Rombo, Northern Tanzania. Matern. Child Nutr. 2016, 12, 516–527.
- Dewey, K.G.; Adu-Afarwuah, S. Systematic review of the efficacy and effectiveness of complementary feeding interventions in developing countries. Matern. Child Nutr. 2008, 4 (Suppl. 1), 24–85.
- Panjwani, A.; Heidkamp, R. Complementary feeding interventions have a small but significant impact on linear and ponderal growth of children in low- and middle-income countries: A systematic review and meta-analysis. J. Nutr. 2017, 147, 2169s–2178s.
- Bashiry, M.; Javanmardi, F.; Sadeghi, E.; Shokri, S.; Hossieni, H.; Oliveira, C.A.F.; Mousavi Khaneghah, A. The prevalence of aflatoxins in commercial baby food products: A global systematic review, meta-analysis, and risk assessment study. Trends Food Sci. Technol. 2021, 114, 100–115.
- Humphrey, J.H.; Mbuya, M.N.N.; Ntozini, R.; Moulton, L.H.; Stoltzfus, R.J.; Tavengwa, N.V.; Mutasa, K.; Majo, F.; Mutasa, B.; Mangwadu, G.; et al. Independent and combined effects of improved water, sanitation, and hygiene, and improved complementary feeding, on child stunting and anaemia in rural Zimbabwe: A cluster-randomised trial. Lancet Glob. Health 2019, 7, e132–e147.
- Null, C.; Stewart, C.P.; Pickering, A.J.; Dentz, H.N.; Arnold, B.F.; Arnold, C.D.; Benjamin-Chung, J.; Clasen, T.; Dewey, K.G.; Fernald, L.C.; et al. Effects of water quality, sanitation, handwashing, and nutritional interventions on diarrhoea and child growth in rural Kenya: A cluster-randomised controlled trial. Lancet Glob. Health 2018, 6, e316–e329.
- Luby, S.P.; Rahman, M.; Arnold, B.F.; Unicomb, L.; Ashraf, S.; Winch, P.J.; Stewart, C.P.; Begum, F.; Hussain, F.; Benjamin-Chung, J.; et al. Effects of water quality, sanitation, handwashing, and nutritional interventions on diarrhoea and child growth in rural Bangladesh: A cluster randomised controlled trial. Lancet Glob. Health 2018, 6, e302–e315.
- Hoffmann, V.; Jones, K.; Leroy, J.L. The impact of reducing dietary aflatoxin exposure on child linear growth: A cluster randomised controlled trial in Kenya. BMJ Glob. Health 2018, 3, e000983.
- Benkerroum, N. Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint. Int. J. Environ. Res. Public Health 2019, 16, 3633.
- Saltzmann, J.; Xu, Y.; Gong, Y.Y.; Lindahl, J.; Kersten, S.; Dänicke, S.; Routledge, M.N. Preliminary study on the relationship between aflatoxin-bovine serum albumin adducts in blood and aflatoxin M1 levels in milk of dairy cows. Mycot. Res. 2020, 36, 207–211.
- Chen, G.; Gong, Y.Y.; Kimanya, M.E.; Shirima, C.P.; Routledge, M.N. Comparison of urinary aflatoxin M1 and aflatoxin albumin adducts as biomarkers for assessing aflatoxin exposure in Tanzanian children. Biomarkers 2018, 23, 131–136.
- WHO. Principles for Evaluating Health Risks in Children Associated with Exposure to Chemicals; World Health Organization: Geneva, Switzerland, 2006.
- Sadeghi, N.; Oveisi, M.R.; Jannat, B.; Hajimahmoodi, M.; Bonyani, H.; Jannat, F. Incidence of aflatoxin M1 in human breast milk in Tehran, Iran. Food Control 2009, 20, 75–78.
- Mahdavi, R.; Nikniaz, L.; Arefhosseini, S.R.; Vahed Jabbari, M. Determination of aflatoxin M(1) in breast milk samples in Tabriz-Iran. Matern. Child Health J. 2010, 14, 141–145.
- Montesano, R.; Hainaut, P.; Wild, C.P. Hepatocellular carcinoma: From gene to public health. J. Natl. Cancer Inst. 1997, 89, 1844–1851.
- Gong, Y.Y.; Egal, S.; Hounsa, A.; Turner, P.C.; Hall, A.J.; Cardwell, K.F.; Wild, C.P. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: The critical role of weaning. Int. J. Epidemiol. 2003, 32, 556–562.
- Gong, Y.Y.; Hounsa, A.; Egal, S.; Turner, P.C.; Sutcliffe, A.E.; Hall, A.J.; Cardwell, K.; Wild, C.P. Postweaning exposure to aflatoxin results in impaired child growth: A longitudinal study in Benin, West Africa. Environ. Health Perspect 2004, 112, 1334–1338.
- IARC (International Agency for Research on Cancer). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Lyon, France, 1993; Volume V56, pp. 51–72.
- NIH. Mycotoxin Mitigation Trial. 2022. Available online: https://www.clinicaltrials.gov/ct2/show/NCT03940547 (accessed on 31 August 2022).
- Turner, P.C.; Moore, S.E.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Modification of immune function through exposure to dietary aflatoxin in Gambian children. Environ. Health Perspect. 2003, 111, 217–220.
- Smith, L.E.; Stoltzfus, R.J.; Prendergast, A. Food chain mycotoxin exposure, gut health, and impaired growth: A conceptual framework. Adv. Nutr. 2012, 3, 526–531.
- Tessema, M.; De Groote, H.; Brouwer, I.D.; De Boevre, M.; Corominas, A.V.; Stoecker, B.J.; Stoecker, B.J.; Feskens, E.J.; Belachew, T.; Karakitsou, A.; et al. Exposure to aflatoxins and fumonisins and linear growth of children in rural Ethiopia: A longitudinal study. Public Health Nutr. 2021, 24, 3662–3673.
- Mbuya, M.N.; Humphrey, J.H. Preventing environmental enteric dysfunction through improved water, sanitation and hygiene: An opportunity for stunting reduction in developing countries. Matern. Child Nutr. 2016, 12 (Suppl. 1), 106–120.
- Harper, K.M.; Mutasa, M.; Prendergast, A.J.; Humphrey, J.H.; Manges, A.R. Environmental enteric dysfunction pathways and child stunting: A systematic review. PLoS Negl. Trop. Dis. 2018, 12, e0006205.
- Keusch, G.T.; Rosenberg, I.H.; Denno, D.M.; Duggan, C.; Guerrant, R.L.; Lavery, J.V.; Tarr, P.I.; Ward, H.D.; Black, R.E.; Nataro, J.P.; et al. Implications of acquired environmental enteric dysfunction for growth and stunting in infants and children living in low- and middle-income countries. Food Nutr. Bull. 2013, 34, 357–364.
- Humphrey, J.H.; Jones, A.D.; Manges, A.; Mangwadu, G.; Maluccio, J.A.; Mbuya, M.N.; Moulton, L.H.; Ntozini, R.; Prendergast, A.J. The sanitation hygiene infant nutrition efficacy (SHINE) trial: Rationale, design, and methods. Clin. Infect. Dis. 2015, 61 (Suppl. 7), S685–S702.
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Mbuya, M.N.; Mutasa, K.; Kembo, G.; Stoltzfus, R.J. The Potential Role of Mycotoxins as a Contributor to Stunting in the SHINE Trial. Clin. Infect. Dis. 2015, 61 (Suppl. 7), S733–S737.
- Weber, T.K.; Polanco, I. Gastrointestinal microbiota and some children diseases: A review. Gastroenterol. Res. Pract. 2012, 2012, 676585.
- Sampson, T.R.; Mazmanian, S.K. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe 2015, 17, 565–576.
- Obuseh, F.A.; Jolly, P.E.; Kulczycki, A.; Ehiri, J.; Waterbor, J.; Desmond, R.A.; Preko, P.O.; Jiang, Y.; Piyathilake, C.J. Aflatoxin levels, plasma vitamins A and E concentrations, and their association with HIV and hepatitis B virus infections in Ghanaians: A cross-sectional study. J. Int. AIDS Soc. 2011, 14, 53.
- Onyemelukwe, G.; Ogoina, D.; Ibiam, G.E.; Ogbadu, G.H. Aflatoxins in body fluids and food of Nigerian children with protein-energy malnutrition. Afr. J. Food Agric. Nutr. Dev. 2012, 12, 6553–6566.
- Hatem, N.L.; Hassab, H.M.; Abd Al-Rahman, E.M.; El-Deeb, S.A.; El-Sayed Ahmed, R.L. Prevalence of aflatoxins in blood and urine of Egyptian infants with protein-energy malnutrition. Food Nutr. Bull. 2005, 26, 49–56.
- Coulter, J.B.; Hendrickse, R.G.; Lamplugh, S.M.; Macfarlane, S.B.; Moody, J.B.; Omer, M.I.; Suliman, G.I.; Williams, T.E.; Liverpool/Khartoum Aflatoxin Study Group. Aflatoxins and kwashiorkor: Clinical studies in Sudanese children. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 945–951.
- Hendrickse, R.G.; Coulter, J.B.S.; Lamplugh, S.M.; Macfarlane, S.B.J.; Williams, T.E.; Omer, M.I.A.; Suliman, G.I. Aflatoxins and kwashiorkor: A study In Sudanese children. Br. Med. J. (Clin. Res. Ed.) 1982, 285, 843–846.
- Ortega-Beltran, A.; Bandyopadhyay, R. Contributions of integrated aflatoxin management strategies to achieve the sustainable development goals in various African countries. Glob. Food Secur. 2021, 30, 100559.
- Fakhri, Y.; Rahmani, J.; Oliveira, C.A.F.; Franco, L.T.; Corassin, C.H.; Saba, S.; Rafique, J.; Khaneghah, A.M. Aflatoxin M1 in human breast milk: A global systematic review, meta-analysis, and risk assessment study (Monte Carlo simulation). Trends Food Sci. Technol. 2019, 88, 333–342.