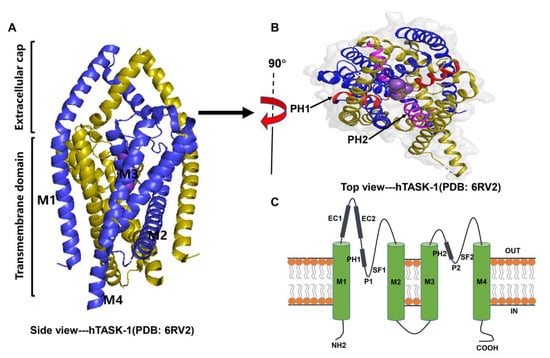
3. The Biological Roles of TASK Channels
3.1. Breathing Rhythm
TASK channels are sensitive to extracellular pH and are expressed in multiple chemosensory nuclei or carotid bodies, which can sense PCO
2 shifts in the early stages of acidosis [
19,
20,
21]. The respiratory stimulant doxapram inhibits the activities of human TASK-1 and TASK-3 channels with effective half doses of 4 and 2.5 µM, respectively, suggesting that TASK channels are involved in regulating breathing rhythm [
22]. Pharmacological and single-channel experiments revealed that TASK-1, TASK-3, and TASK-1/TASK-3 heterodimers play critical roles in the chemosensation of the carotid body and are major sensors of hypoxia and metabolic acidosis [
23]. In neural recordings from electrodes implanted at the carotid body/sinus from TASK-1
−/− mice, a significant reduction (49% and 68%, respectively) in chemoafferent cell firing induced by hypoxia (10% O
2) and hypercapnia (3–6% CO
2) was found along with a blunted ventilatory response to hypoxia. No changes in TASK-3
−/− mice were observed under the same conditions, suggesting that TASK-1 in particular plays a key role in peripheral chemosensing. However, the TASK-3 channel can mediate hypoxic depolarization of normal glomus cells [
24]. Studies using TASK-1 channel knockout (KO) mice also showed that pH sensitivity in serotonergic raphe neurons was abolished but was maintained in the retrotrapezoid nucleus (RTN). Because the RTN is central to the normal ventilatory response to CO
2, this indicates that TASK-1 channels are not involved in the regulation of central respiratory chemosensitivity [
25]. Alkaline-sensitive TASK-2 channels are widely distributed in Phox2b-expressing neurons in the brainstem RTN, which are the principal central respiratory chemoreceptors. TASK-2 KO mice displayed a diminished ventilatory reaction to CO
2/H
+ in vivo, indicating that TASK-2 channels play a major regulatory role in central respiratory chemoreception [
1,
20,
137,
146].
3.2. Pulmonary Artery Hypertension
Hypoxic pulmonary vasoconstriction (HPV) is an autoregulatory mechanism of the pulmonary vessels in response to hypoxia, which can reduce the ratio of ventilation and blood flow in the hypoxic alveolar region to ensure its oxygen supply. Thus, it plays an overriding role in maintaining the local ventilation/blood flow ratio and a constant arterial partial pressure of oxygen. Potassium channels play an important role in pulmonary arterial smooth muscle cells (PASMCs), and reduced potassium channel activity increases their resistance to apoptosis, cell proliferation, and vascular constriction, leading to vascular remodeling [
147,
148,
149]. Hypoxia inhibits certain potassium channel activities, leading to cell membrane depolarization, which activates voltage-dependent calcium channels. This triggers inward extracellular Ca
2+ flow and the contraction of pulmonary vascular smooth muscle, which ultimately causes increased pulmonary vascular resistance and the initiation of HPV. There are five main potassium channels in PASMCs: voltage-gated potassium (K
v) channels, Ca
2+-activated potassium (K
Ca) channels, ATP-sensitive potassium (K
ATP) channels, inward rectifier potassium (K
ir) channels, and K
2P channels [
150,
151]. Human PASMCs express only TASK-1 potassium channels [
26].
3.3. Cardiac Arrhythmia
TASK-1 potassium channels are atrial-specific, and protein blotting experiments have shown that the TASK-1 channel in the human heart is expressed at 14–16-fold higher levels in the atria than that in the ventricles [
152,
153,
154]. The TASK-1 channel is known to be involved in the pathophysiology of cardiovascular diseases [
37,
38,
39]. It has been found that TASK-1 KO mice have a significantly prolonged action potential duration [
40] and that the use of the TASK-1 blocker A293 significantly prolonged the action potential duration in rat ventricular muscle. This suggests that TASK-1 channels are not only involved in maintaining the resting membrane potential of cardiac myocytes but also help in regulating the outward current during the plateau phase of the action potential [
41]. Class III antiarrhythmic drugs prolong action potentials and inhibit atrial and ventricular arrhythmias. Among these drugs, amiodarone inhibits open and closed TASK-1 channels in a concentration-dependent manner, with an IC
50 value of 0.4 μM in oocytes. This is a possible mechanism for the efficacy of these agents [
42,
43].
3.4. Aldosterone Secretion
It has been found that TASK
−/− channel KO mice exhibit non-tumorigenic primary hyperaldosteronism with overproduction of autonomous aldosterone that was neither suppressed by high dietary sodium intake nor corrected using the angiotensin II receptor 1 antagonist candesartan. These results suggested that TASK channels are possible therapeutic targets for primary hyperaldosteronism [
52]. Furthermore, statistical analyses indicated that KCNK3 (TASK-1) variants are associated with hyperaldosteronism and hypertension [
53]. Another study reported that deleting TASK-1 and TASK-3 channels in the zona glomerulosa of the adrenal tissue causes aldosterone-driven angiotensin II-independent hypertension [
54] Several other studies also indicated that TASK channels participate in the modulation of aldosterone secretion [
55,
56,
57].
3.5. Pain
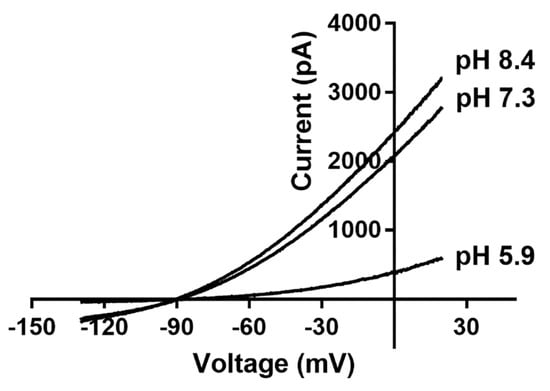
Tissue acidification can cause acute and chronic pain, and TASK channels are very sensitive to extracellular acidification. For example, exposing the channel to an extracellular pH of 6.0 can reduce its current by 96% [
71,
158]. Acid inhibition of TASK channels may be an important mechanism for the sustained depolarization of cells due to tissue acidification. This suggests that they could also be potential targets for tissue acidification-induced nociceptive transmission [
159,
160,
161,
162]. Several studies have shown that TASK channels are closely associated with pain [
58,
59,
72].
Marsh et al. [
60] found that the mRNA levels of TASK channels in the spinal dorsal root ganglion (DRG) were correlated with spontaneous foot lifting after intradermal complete Freund’s adjuvant injections, indicating that TASK channels are associated with inflammation-induced pain. The activity of TASK channels is also altered in pathological models of nerve injury. The mRNA expression of TASK-3 and TWIK-1 channels was found to be downregulated in L4-L5 DRG ipsilateral to the neuropathic lesion one week after spared nerve injury surgery, whereas TASK-1 channel expression remained unchanged [
73]. In addition, it has been reported that the TASK-3 channel co-locates with the transient receptor potential cation channel subfamily M member 8 (TRPM8) in sensory neurons. TRPM8 is a cold- and menthol-activated ion channel that participates in thermosensation [
163]. Liao et al. [
74] designed a peripherally acting selective agonist, CHET3, for the TASK-3 channel and used it to show that the TASK-3 channel is an intrinsic regulator of pain sensation. They proved that CHET3 could attenuate thermal hyperalgesia and mechanical allodynia in different rodent pain models. They also predicted that CHET3 would bind under the SF and close to the M2 and M4 regions, which would alter the gating activity of the channel by affecting the SF conformation. García et al. [
61] found that spinal TASK-1 and TASK-3 channels are involved in the regulation of inflammatory and neuropathic pain and that intrathecal pretreatment with the activator terbinafine could reduce formalin-induced flinching and nociceptive hyperalgesia in rats with neuropathy.
3.6. Anesthetics
3.6.1. Volatile Anesthetics
TASK channels are expressed in CNS sites relevant to anesthetic actions and are the targets of many general anesthetics. Volatile anesthetics were the first group of TASK channel modulators to be extensively studied. Volatile anesthetics, such as halothane and isoflurane, activate TASK channels at clinically relevant concentrations to hyperpolarize cell membranes and reduce excitability [
62]. This is especially true for brainstem motor and thalamocortical neurons and helps explain the loss of consciousness and motor ability that general anesthetics induce [
164,
165] It has been reported that the hyperpolarizing effects of halothane and isoflurane were reduced in TASK-1/TASK-3 KO mice, and the corresponding anesthetic effects of sedation, hypnosis, and immobilization were also diminished [
64]. However, it has also been reported that high concentrations of isoflurane (0.8 mM) activate the TASK-3 channel and the heterodimeric TASK-1/TASK-3 channel, but inhibit the TASK-1 channel. This suggested that different concentrations of volatile anesthetics can regulate the activities of TASK channels in both directions [
65] Studies have shown the presence of a binding region for volatile anesthetics in the structures of TASK channels, which includes the residue M159 at the cytoplasmic terminus of the M3 segment [
66] and, in particular, residues 243 to 248 at the beginning of the cytoplasmic C-terminus, a region also known as the halothane-responsive region [
63,
101]. In addition, it has been shown that halothane and isoflurane can inhibit calcium current in smooth muscle cells of the coronary arteries to directly dilate coronary arteries and increase coronary flow during anesthesia [
166].
3.7. Cancers
The TASK-3 channel is highly expressed in breast cancer, ovarian carcinoma, and melanoma cells, and it has been suggested that it promotes tumor growth and proliferation [
75,
76,
77,
78,
79,
80,
81,
82]. Thus far, only the TASK-3 channel in the K
2P family has been identified to be expressed in the inner mitochondrial membrane, and silencing it induces apoptosis of human melanoma cells [
83,
84]. In addition, a study using a short hairpin RNA (shRNA)-mediated knockdown of the TASK-3 channel confirmed that the TASK-3 channel is involved in migration and cell survival in gastric cancer. Thus, it represents a potential therapeutic target for gastric cancer treatment [
85]. Moreover, the addition of Y4, a monoclonal antibody against the TASK-3 channel extracellular domain, to KCNK9-expressing carcinoma cells can inhibit tumor growth and metastasis [
86]. These confirmed that the TASK-3 channel is a promising target for the treatment of malignancies that express KCNK9 [
87].
3.8. Neurological Activities and/or Disorders
3.8.1. Sleep
Synchronized burst firing and tonic action potential generation in the thalamocortical system of the brain mainly occur during sleep and waking states, respectively [
168]. The switch between these two firing modes is critically modulated by the bidirectional regulation of TASK and TWIK-related K
+ channels (TREK) in thalamic relay neurons.
It has been found that the genetic KO of TASK-1 did not change sleep/wake times in mice [
169]. One study analyzed the locomotor activity and circadian rhythms of TASK-3 KO mice and found that, when compared to wild-type litter controls, both had normal circadian rhythms. However, TASK-3 KO mice had significantly increased nocturnal activity (38%), suggesting that the TASK-3 channel plays an important role in the regulation of sleep [
88]. Another study based on continuous electroencephalogram and electromyogram recordings of TASK-3 KO mice revealed that they had a slower transition from wakefulness to sleep and more fragmented sleep episodes and rapid eye movement (REM) theta wave (4–9 Hz) oscillations during sleep [
89,
90].
3.9. Other Roles
Several studies have shown that the TASK-1 channels found in the pancreas participate in the regulation of glucose homeostasis [
69,
70]. Using the highly specific “RNAscope” method of in situ mRNA hybridization in combination with pharmacological antagonists and genetic deletion tools, Wen et al. [
100] reported the first evidence that the TASK-3 channel is abundantly expressed in retinal ganglion cells and plays a critical role in visual processing in the retina.