Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Mechanics
The design of bird diverters should be based upon the perception of birds, not the perception of humans, but until now it is human vision that has guided diverter design. Aspects of bird vision pertinent to diverter design are reviewed.
- bird diverters
- vision
- light level
1. Vision and Visual Capacities
Vision is the prime sense used by animals to retrieve information about the world that surrounds them. At its most fundamental, vision has evolved to provide information about where objects are over a range of distances [3]. This is true for humans as much as it is for all other species, but the information that can be extracted about objects, and where they are, varies significantly between species. Furthermore, in every species, the accuracy of spatial information also varies significantly with light level, and hence time of day. Animals, including humans, are hostage to the sun and the moon as regards the information that they have available to them about the world
Vision is a complex and multifaceted sense, making it impossible to give a simple summary of the visual ability of any species. To describe vision, it is necessary to break it down into various “visual capacities” which need to be investigated and described separately. The most notable of these visual capacities are spatial resolution, colour vision, speed of vision, and field of view. The first three of these are highly dependent upon the light level and upon position within the field of view. Under natural conditions, light levels change dramatically within the daily cycle (typically 3000-fold between noon and twilight, and up to 100,000-fold at night depending upon the moon’s presence and phase [1]. This means that the ability to retrieve information about objects from a scene varies throughout every daily cycle to a significant degree.
2. Changing Flight Behaviour
Like vision, behaviour is complex and multifaceted, there are multiple ways of describing and measuring behaviour. Power line diverters seek to change a quite specific part of a bird’s repertoire at a particular location. While diverters seek to alter the flight path of birds it is necessary to recognize that flight, like vision, is multifaceted and differs between species. Each bird species has a specific flight pattern, manoeuvrability, and characteristic speed of powered flight [4,5]. Bird species also differ in the time of day at which they are most active or undertake flights in their daily cycle. One species will fly in twilight, while another will not fly until the sun is well above the horizon, and some species will fly during nighttime. Some have flight which is slow and highly manoeuvrable, while others fly fast and cannot readily change their trajectories [5].
3. Will a Diverter Divert?
The application of knowledge of vision and flight behaviour to the design of bird diverters can be reduced to one key question: will the diverting device be detected at a distance that should allow a timely change in flight path? Key to answering this question is an understanding of which species a diverting device is aimed at. Additionally, at which locations, at what time of day, and under what weather conditions should the device have its desired effect? Weather conditions and light levels become important because they will determine what can be detected and at what distance.
In this context, it is worth reflecting upon our own perception of objects. What we see/detect, and how we interpret a natural scene, depends crucially upon the light level and weather conditions. A telling example of how these conditions are relevant to our behaviour, indeed our well-being, is car driving. What we detect, and how we interpret information when driving, has been subject to much research [2]. When driving we are constantly seeking new information about objects at a distance from us, towards which we are travelling. Whether we detect an object, and whether we interpret it correctly, depends significantly upon light levels and weather conditions [6,7].
Safe driving depends upon constant updates of information about the road ahead and interpreting changes correctly. Particularly important is the fact that we often, perhaps typically, rely upon partial information and frequently drive beyond our perceptual limit [7]. That is, we often do not have full information to determine what lies ahead, but we rely upon the predictability of roads and their standard design features to allow us to make judgements based on partial information.
Usually, we manage to behave correctly even though we have partial information from our vision. However, if conditions become exacting, and the acquisition of information is impaired, accidents can occur, even partial information may not be available and correct interpretations are not possible. In these more exacting circumstances, we need repeated information to warn us that the world ahead is changing before we fully recognize the true nature of the situation and change behaviour accordingly.
There would seem to be parallels here with the situations posed to birds by power lines and other obstacles that intrude into the air space in which they fly. Under these circumstances, birds are also likely to be relying upon partial or incomplete spatial information, as some species are known to do when foraging [1,8]. Natural objects that intrude into a bird’s open flight space are rare and it is reasonable that the bird will often predict that there is little likelihood of an obstacle ahead. Thus, warning of a power line ahead may require significant and apparently excessive signals, much as is required to warn us that an obstacle lies ahead when driving on an open or fast road. Furthermore, when environmental conditions reduce the information available to the birds (particularly when light levels are low, and visibility is reduced) it may require significant and repeated signals to warn the birds of the obstacle and change their behaviour.
4. Visual Spatial Resolution and Light Level
In all animals, visual-spatial resolution decreases significantly with light level [3]. The examples of visual acuity in a range of bird species given in Table 1 apply to high daytime light levels, approximately those that occur in open habitats around midday under the full sun. Changes in spatial resolution within the range of light levels experienced during the day are typically small (heavy cloud reduces light levels by about 10-fold), however, as light levels fall into twilight a significant decrease in resolution occurs (Figure 1). At sunset, light levels have fallen by about 300-fold compared with noon and fall by another 100-fold during the twilight period. These changes in light level have a significant impact on spatial resolution (Figure 1).
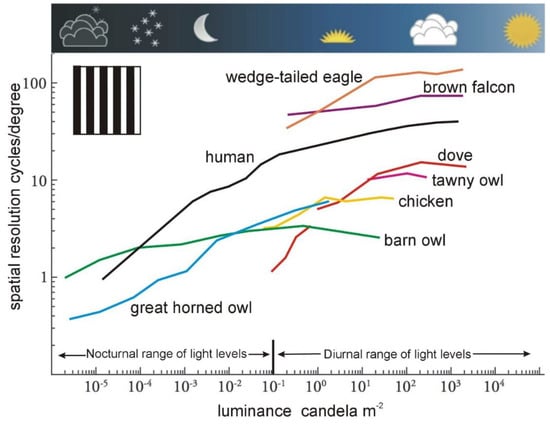
Figure 1. Resolution and light levels. The effect of light levels on visual resolution is significant. Acuity is often measured or estimated at high daytime light levels, but in a few bird species acuity has been measured across almost the full range of naturally occurring light levels. In some species, acuity has been determined across the narrower range of the light levels that occur in daytime and in some species across the daylight-twilight range. This figure shows some key points about the effect of light levels on spatial resolution. In all species shown here acuity has been determined using behavioural training techniques. Although the maximum acuities of these different species are significantly different (for example, the maximum acuity of the eagle is 60 times higher than that of the dove) it is also clear that in all species acuity decreases considerably as natural light levels fall. The only species in which there is not a steep decline in acuity are Western Barn Owls Tyto alba, but even in Barn Owls acuity shows a significant fall as light levels decrease to the lower ranges of night-time. In some species, the decline is particularly steep and may be a significant reason why some daytime birds, such as doves, go to roost as light levels fall. (From [1], Figure 2.16).
Table 1. Spatial resolution (visual acuity) in a sample of bird species and in humans. All measures refer to acuity at high daytime light levels and indicate the best performance (finest spatial detail that can be resolved using high contrast stimuli). To compare between species simply divide their acuity values. Thus, the goose’s resolution is about 7 times higher than that of a young human. Expressed another way, a young human should be able to detect a given object 7 times further away than a goose, while an older human can detect the same object 3 times further away.
| Species | Visual Acuity (Minutes of arc) |
Source |
|---|---|---|
| Wedge-tailed Eagle Aquila audax | 0.2 | [9] |
| Brown Falcon Falco berigora | 0.4 | [10] |
| Rock Dove Columba livia | 1.7 | [11] |
| Budgerigar Melopsittacus undulatus | 2.6 | [12] |
| Canada Goose Branta canadensis | 3.1 | [13] |
| Bourke’s Parrot Neopsephotus bourkii | 3.2 | [12] |
| Great Horned Owl Bubo virginianus | 4 | [14] |
| House Sparrow Passer domesticus | 6.3 | [15] |
| Human (young) | 0.4 | [3] |
| Human (mature/with spectacles) | 1.0 | [3] |
A good rule of thumb is that visual resolution during twilight can be reduced by up to 10-fold compared with maximum resolution (acuity) [2]. This means that an animal would have to be 10 times closer to detect a given object at twilight than at midday. While this is confirmed by our everyday experiences, it applies equally to birds and will be behaviorally significant to birds which readily fly during twilight [16].
This reduction in resolution with light level applies to all diurnally active birds (Figure 1), and it has been argued [1] that many birds go to roost at dusk primarily because their visual resolution drops sufficiently that they cannot negotiate obstacles safely. (Note that owls have a different set of sensory and behavioural strategies that allow them to be nocturnally active within complex, woodland-type, habitats. However, their spatial resolution is low (Table 1 and Figure 1) and they may rely on only partial visual information when active at nighttime light levels [1].
5. Fields of View
The human field of view is unusual. Few other animal species exhibit the same features. Two eyes look straight out from the face. This results in a view of the environment that at any one instant takes in little more than half of the world about the head. The world always appears to lie in front of us and very little can be detected above and below, even in the frontal field. Moreover, the direction in which spatial resolution is highest lies directly ahead and we see less detail in our visual periphery [1].
This is quite unlike the field of view of birds. In all bird species the eyes are placed laterally in the skull (Figure 2) and together view a large sector of the world about the head (Figure 3). Some species, including some ducks, achieve total panoramic vision. That is, they see 360 degrees around the head and the full hemisphere above it. On the other hand, some of the larger birds of prey (Accipitridae) and bustards (Otididae) have been shown to have such extensive blind areas above their heads that when they pitch their head forwards to scan the ground below, they are flying blind in their direction of travel (Figure 3) [17]. Furthermore, in birds, the direction of highest spatial resolution projects approximately along the optic axis of each eye (Figure 2) with the result that birds have two directions of highest acuity, one on either side of the head, not directly in front of it.
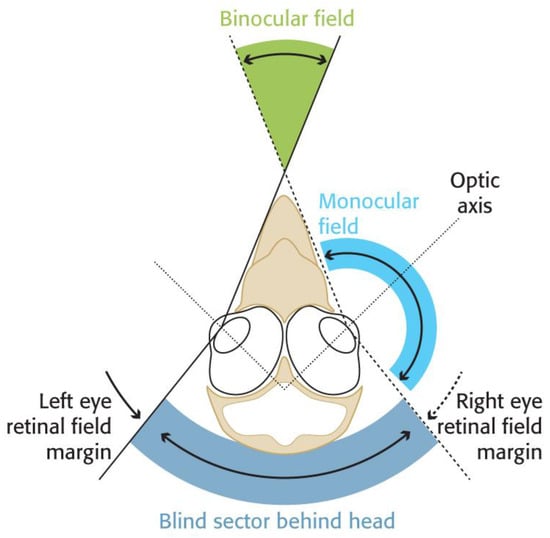
Figure 2. A diagrammatic section through the head of a bird showing a typical arrangement of eyes in the skull and how the visual fields of each eye combine. In all birds the eyes project laterally so that the axes of the eyes always diverge, no birds have forward-facing eyes. The field of view of each eye is combined to give the total field of view of the bird and to give a sector in front of the head where the fields of the two eyes overlap to give a binocular field. A wide degree of variation in these basic arrangements is found in birds and gives different degrees of binocular overlap and different width blind areas behind and above the head. Just small variations in the width of the field of view of each eye, and of eye position in the skull, can result in large differences in visual fields between species. The optic axis of each eye defines the approximate direction of highest visual acuity. Hence, birds have at least two areas of high spatial resolution that project into their lateral fields of view. (Diagram by the author and drawn by Nigel Hawtin (nigelhawtin.com, accessed on 5 December 2022)).
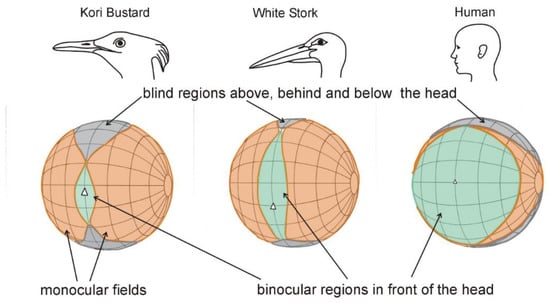
Figure 3. Depictions of the total visual fields of White Storks Ciconia ciconia, Kori Bustards Ardeotis kori, and Humans in which the key features of the field are shown as projected onto the surface of a sphere that surrounds the head. This presentation emphasizes how different the visual fields of these three species are. This is despite the visual fields of their individual eyes being similar. It is clear that differences in eye position in the skull, as well as differences in the visual fields of individual eyes, can result in quite different ways in which birds can extract information from the world about their heads. Kori Bustards are an example of a species which loses forward vision when the head is pitched forward to scan the ground below. White Storks and humans, on the other hand, maintain forward vision when the head is pitched forward through a large angle (Modified from [1], Figure 2.14).
The visual significance of these arrangements is that in the majority of birds visual information can be gained at any instant from an extensive volume about their head and that there are at least two regions of enhanced resolution in their visual field. However, these regions project laterally rather than directly forwards. The result of this is that to examine an object with the highest acuity, a bird is required to turn its head and appears to look at the object “sideways” rather than examine it with the head facing forwards. The main differences between the human and bird view of the world can be summed up as: humans have the impression that they move forward “into” the world and are usually moving towards a point where their spatial resolution is highest. Birds, on the other hand, can be considered to be flowing “through” the world. For the majority of birds, objects appear in front, flow past, and disappear behind, and the region of highest spatial resolution is lateral to the head rather than directly in front.
The crucial role of binocular vision in birds is not in the detailed examination of something that lies ahead [8,18]. Objects of interest tend to be detected laterally using the region of highest resolution, and visual control is subsequently passed to the binocular field [19]. The binocular field functions primarily in the control of the direction of travel towards an object, and in determining the time to contact that object, rather than in determining the distance to that object. Both the direction of travel and time to contact are determined directly from the pattern of the optical flow of images across the retina in each eye [20,21], and it is in the binocular region of each eye that the flow pattern expands symmetrically about the direction of travel [21,22,23].
The region of binocular vision in birds is relatively narrow (typically between 20 and 40 degrees wide) and vertically long (100 to 180 degrees) [1]. In Canada Geese, the binocular field that lies directly ahead of a bird in flight is about 22 degrees wide [24]. In some ducks, the binocular field is only 10 degrees wide, but in some birds of prey and passerine species, it is 40–50 degrees wide. See [1] for a comparative table of binocular field width in a range of bird species. In humans, the binocular field is 120 degrees wide (Figure 3).
This entry is adapted from the peer-reviewed paper 10.3390/birds3040028
This entry is offline, you can click here to edit this entry!