Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Endocrinology & Metabolism
虽然异柠檬酸脱氢酶已为人所知数十年,但它们在肿瘤中的机制正逐渐变得清晰。人体细胞中的三种异柠檬酸脱氢酶亚型在亚细胞定位、分子结构、辅因子要求和催化机制方面有所不同。在这里,我们回顾了异柠檬酸脱氢酶在新陈代谢中的作用,提供了与癌症相关的异柠檬酸脱氢酶的当前概述,并指出了未来的研究方向,以更好地了解健康和疾病中的复杂生物学。
- isocitrate dehydrogenase (IDH)
- cancer metabolism
- oxidative decarboxylation
- nicotinamide adenine dinucleotide phosphate(NADPH)
- α-ketoglutarate(α-kg)
一、简介
异柠檬酸脱氢酶(IDH)在癌症中的研究分为野生型IDH和突变型IDH [ 1 ]。许多以前的研究都将肿瘤视为一个单一的实体,而不区分突变型和野生型 IDH。然而,现在人们普遍认为,野生型 IDH和突变型 IDH肿瘤具有根本不同 [ 1 ]。近年来,肿瘤IDH的研究主要集中在突变类型,许多有效的突变型IDH抑制剂已应用于临床并取得了可喜的成果[ 1 ]。突变体 IDH 减少 α-KG 的产生,减少来自异柠檬酸和 NADP +的 NADPH, 并催化 α-KG 转化为 D-2-羟基戊二酸 (D-2HG) [ 2 ]。提出升高的 D-2HG 可促进肿瘤发生 [ 1 ]。IDH1 和 IDH2 突变导致活性位点替换,从而深刻影响 IDH 活性、细胞代谢和癌症发展[ 1、3、4 ]。同时,近年来野生型IDH在肿瘤中的研究也取得了重大进展。多项研究表明,肿瘤中野生型 IDH表达增加的预后比突变型 IDH增加的预后更差[ 5 ]。90% 的胶质瘤为野生型 IDH,预后比突变型 IDH差[5 ]。野生型 IDH 酶在促进肿瘤增殖和治疗复发方面起着关键作用。因此,阐明野生型IDH在肿瘤中的作用机制至关重要。
在正常情况下,细胞需要足够的氧气浓度 (O 2 ) 进行有氧呼吸,产生三磷酸腺苷 (ATP),这是生物体最直接的能量来源。尽管高度增殖的肿瘤细胞消耗大量葡萄糖,但它们产生ATP的效率很低[ 6、7、8、9 ]。这是因为快速增殖的细胞不需要产生大量 ATP 来执行其细胞功能,但必须合成生物分子,如核苷酸、氨基酸和脂肪酸,以促进细胞分裂和生长[ 6、7、8 ]。还原羧化为这种合成代谢过程提供了捷径[10 , 11 ]。IDH1和IDH2介导α-KG在还原羧化反应中直接生成异柠檬酸[ 3 ],有学者将此反应过程描述为还原TCA或“逆时针” TCA [ 12,13,14 ]。有趣的是,一些研究主张 IDH1 和 2 催化的还原羧化作用无关紧要,因为它会将 α-KG 转化回异柠檬酸,从而形成“无效循环”[ 9 ]。
许多研究表明,它是一种重要的代谢途径,尤其是在快速增殖的肿瘤细胞中。例如,发现当肿瘤细胞缺乏谷氨酰胺或在不含 α-KG 的培养基中生长时,肿瘤细胞的生长速度会降低 [ 15 , 16 ]。肿瘤细胞对谷氨酰胺的消耗增加导致谷氨酰胺衍生的异柠檬酸盐增加,进而产生更多的柠檬酸盐 [ 10 , 11 ]。柠檬酸盐不能自由穿过线粒体内膜,必须通过柠檬酸苹果酸丙酮酸循环进入细胞质 [ 17 ]。ATP 柠檬酸裂解酶催化柠檬酸可逆转化为乙酰辅酶 A,后者是脂肪酸合成的原料 [ 17]. 脂肪酸是新膜形成的关键,也是细胞快速增殖所必需的 [ 10 ]。IDH介导的异柠檬酸氧化脱羧也具有重要意义。一方面,它是TCA循环的重要组成部分,为呼吸链生成ATP提供氢原子。另一方面,NAD(P) + /NAD(P)H 氧化还原比率转变被改变以适应细胞代谢的变化以执行其他功能,这将在下面讨论 [ 18 ]。
人类异柠檬酸脱氢酶 (IDH) 基因编码同工酶 IDH1、2 和 3,它们催化异柠檬酸和 α-酮戊二酸 (α-KG) 的转化,并且是正常哺乳动物代谢所必需的。异柠檬酸脱氢酶 1 和 2 催化异柠檬酸可逆转化为 α-KG。异柠檬酸脱氢酶 3 是介导三羧酸 (TCA) 循环中异柠檬酸生成 α-KG 的关键酶。在TCA循环中,异柠檬酸脱氢酶催化的脱羧反应介导异柠檬酸转化为α-KG并伴有脱氢作用,这一过程通常称为氧化脱羧。α-KG和CO 2形成6-C异柠檬酸盐IDH催化的反应称为还原羧化。这种 IDH 介导的可逆反应在肿瘤细胞中非常重要。
2. IDHs的催化机制和酶结构
三种 IDH 亚型在人类细胞中的表达在亚细胞定位、分子结构、辅因子需求和催化机制方面有所不同(图 1)[ 19 ]。IDH1 位于细胞质和过氧化物酶体中,而 IDH2 和 IDH3 位于线粒体中 [ 20 ]。IDH1和IDH2(EC 代码:1.1.1.42)具有相当大的序列相似性(在人类中有 70% 的同一性)并催化相同的反应[ 20、21 ]。IDH1 和 IDH2 可催化NADPH 和 CO 2将αKG 还原羧基化为异柠檬酸,从而催化可逆反应 [ 19 , 20 , 21]. 通过对大鼠肝脏中的 TCA 循环中间体进行同位素分析,也观察到了可逆的 CO 2固定 [ 22 ]。
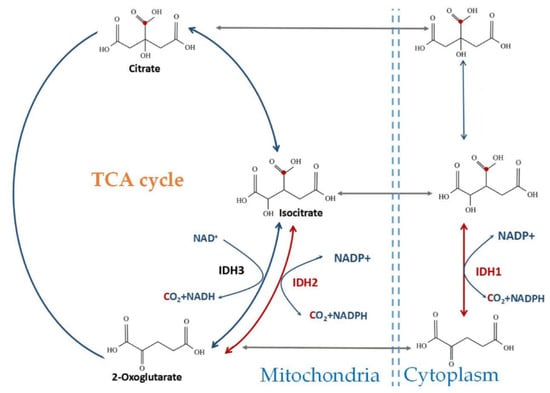
图 1.野生型 IDH 催化的亚细胞定位和化学反应。IDH1 位于胞质溶胶中;IDH2 和 IDH3 位于线粒体基质中。IDH1/2 可逆地将异柠檬酸氧化为 α-KG 和 CO 2并形成 NADPH。IDH3 不可逆地将异柠檬酸氧化为 α-KG,同时形成 NADH 作为 TCA 循环的一部分。
IDH1 作为同型二聚体存在,每个 IDH1 单体具有一个钳结构域、两个大结构域和两个小结构域。大结构域和小结构域由 β-折叠 [ 23 ] 连接。IDH2 和 IDH1 是结构非常相似的同工酶 [ 20 ]。在TCA循环中,IDH3(EC代码:1.1.1.41)以NAD +为辅因子催化反应形成α-KG和NADH。该反应是不可逆的,IDH3 受柠檬酸盐、二磷酸腺苷和三磷酸腺苷的变构调节 [ 24 ]。α-KG 进一步代谢为琥珀酸,而 NADH 被电子传递链氧化 [ 25]. IDH3 异二聚体由 αβ- 和 αγ- 亚基组成,它们组装形成 α2βγ 异四聚体 [ 24 ]。IDH3α-亚基由IDH3A基因(染色体 15q25.1)编码,β-亚基由IDH3B基因(染色体 20p13)编码,γ-亚基由IDH3G基因(染色体 Xq28)编码 [ 24 ]。α亚基是催化亚基[ 26 ],β和γ亚基是调节亚基[ 27 ]。αβ和αγ单独具有基础活性,但IDH3的全部活性需要这两个异二聚体的组装和协同作用[ 24 ]。
This entry is adapted from the peer-reviewed paper 10.3390/cancers14235779
This entry is offline, you can click here to edit this entry!