Skin is the largest organ of the human body and is a great shield, as it protects it from external infections (environmental and chemical pollutants) as well as from UV irradiation. However, it is vulnerable since its degradation can occur both due to extrinsic and intrinsic factors, leading to early aging. Among all, extrinsic skin aging, called photoaging, is a remarkable result of oxidative stress caused by UV irradiation. In addition, reactive oxygen species (ROS) have also been found to contribute to skin aging, as they are produced in skin cells through UV irradiation, although at low concentrations they could be beneficial for some signaling pathways. Environmental and chemical pollutants also produce ROS, triggering a number of pathologies. Skin’s connective tissue includes a number of constituents, including collagen fibrils, elastic fibers, glycoproteins, and glycosaminoglycans. Among all, proteins like elastin, collagen, the glycosaminoglycan hyaluronic acid, and a polymeric pigment called melanin play pivotal roles in the regulation of skin’s elasticity as well as its protection against UV irradiation.
1. Human Neutrophil Elastase (HNE)-A Serine Protease
Neutrophil elastase [EC 3.4.21.37] (known as Human Leukocyte Elastase (HLE) or elastase 2) is a glycoprotein belonging to chymotrypsin serine proteases family [
16]. The active form of HNE can be found at a concentration of ~5 mM in neutrophil lysosomes (azurophils), which act as HNE carriers [
17]. HNE has specificity for protein hydrolysis and is responsible for the degradation of proteins located in the azurophils, as well as extracellular matrix (ECM) proteins [
17]. HNE hydrolyzes all the prime proteins constituting the connective tissue, such as collagen-IV, elastin, fibronectin, proteoglycans, keratins, and other ECM proteins [
18,
19]. However, elastase shows great specificity in degrading elastin, a protein responsible for the elasticity in arteries, lungs, ligaments, and skin [
18,
20,
21]. When bacteria and microorganisms invade the ECM through wounds, the mechanism of inflammation is activated, leading to the secretion of HNE, resulting in tissue repair, proving the vital role of elastin [
21,
22]. Along with collagenase, elastase regulates skin tissue homeostasis. In contrast, if there is no homeostatic control, elastase degrades several connective tissue proteins [
23], causing early skin aging. Furthermore, elastase plays a vital role in conditions damaging the lung tissues, leading to the development of many inflammatory disorders, such as acute respiratory distress syndrome, pulmonary emphysema, lung inflammation, and even rheumatoid arthritis.
HNE is a 30-kD glycoprotein consisting of 218 amino acids and four disoulfide bridges (Cys: 42–58, 136–201, 168–182 and 191–220) that form two stable antiparallel cylindrical domains of beta-barrels [
24]. HNE shows specificity in degrading peptide bonds between small hydrophobic amino acids. The potent catalytic activity of HNE depends on a catalytic triad that contains the amino acids Histidine (His57), Serine (Ser195), and Aspartate (Asp102). HNE’s structure consists of five distinguished hydrophobic pockets, four substrate-binding pockets (S2, S3, S4, and S5), and one catalytic pocket (S1) [
24,
25]. The S1 pocket is hemispheric and hydrophobic and binds to medium-sized aliphatic amino acids. The S2 pocket has a bowl-like shape and prefers medium-sized aliphatic amino acids like Pro. S3, S4, and S5; the pockets take part in the bond formation of HNE with the appropriate ligand. HNE’s catalytic mechanism is similar to that of the homologous serine protease chymotrypsin [
26,
27].
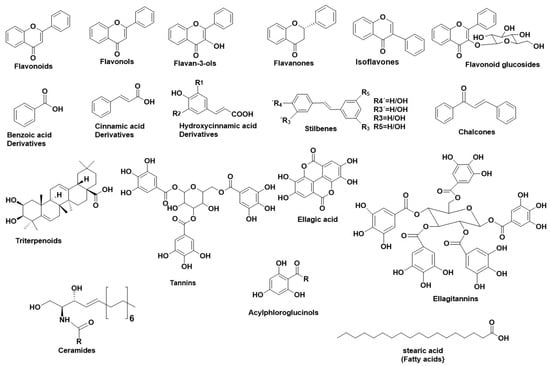
Figure 1 illustrates the main structural scaffolds that have been marked as potent HNE inhibitors. Polyphenols [
40] phenolic acids, cinnamic acids (and derivatives), caffeic acids (and derivatives), [
18,
41,
42,
43] stilbenes, chalcones, elagitannins [
44] pentacyclic triterpenes, fatty acids [
45], ceramides [
46], and acylphloroglucinols have been studied extensively for their anti-HNE properties. The characteristic flavonoid structure and the number of the -OH groups affect their interaction with the HNE’s active site [
47]. Gallic acid derivatives, which were estimated as potent elastase inhibitors, interacted with HNE’s active site through hydrophobic forces and hydrogen bonds [
28]. Caffeic acid derivatives, such as bornyl caffeate [
40,
41] (a bicyclic caffeic acid derivative), also inhibited HNE efficiently (IC
50 = 1.6 μΜ). Additionally, in silico studies revealed an interaction between the caffeic acid moiety with the amino acids of the “oxyanion hole” of HNE
40,. Similarly, bornyl coumarate and bornyl ferulate (hydroxycinnamic acid derivatives) interacted with HNE’s active site as bornyl caffeate [
40,
41].
N-octylcaffeic acid (an alkylated caffeic acid derivative) also demonstrated strong HNE inhibition (IC
50 = 1 μΜ). The silico studies showed that
N-octylcaffeic acid interacted with HNE’s catalytic triad [
42]. The natural products that have been studied for their inhibitory properties towards HNE can be explored in the new developed Server and Database called ANTIAGE-DB, through the link
https://bio-hpc.ucam.edu/anti-age-db
Figure 1. Main structural scaffolds which have been marked as potent HNE inhibitors.
2. Hyaluronidase (Hyal)—A Glycosyl Hydrolase
Hyalouronidases (Hyals) [E.C. 3.2.1.35] consist of a subclass of glycosyl hydrolases mainly responsible for degrading the glycosaminoglycan Hyaluronic Acid (HA) [
60] by cleaving its
β-1→4 glycosidic bonds into oligomers. HA can be found in abundance in synovial fluid, in cartilage (mainly in the soft part of connective tissue), and in the vitreous part of the eye [
61]. Furthermore, HA can be found in the extracellular matrix as a connecting fluid of protein filaments, collagen fibers, and connective tissue cells [
62].
Studies made on Hyals by Karl Meyer (1971) [
63] differentiated them into three groups according to their reaction mechanism: (1) bacterial hyaluronidases (E.C. 4.2.99.1) (endo-
β-acetyl-hexosaminidases), (2) hyaluronidases (E.C. 3.2.1.36) (endo-
β-glucuronidases), and (3) mammalian hyaluronidases (E.C. 3.2.1.35) (hydrolases, producing tetrasaccharides and hexasaccharides).
Hyals contain five homologous molecules expressed in the human gene: Hyal-1, Hyal-2, Hyal-3, Hyal-4, and Hyal-5 (SPAM-1 or PH-20), as well as a pseudogene (pHYAL1) [
60]. Hyal-1, Hyal-2, and Hyal-3 are found in chromosome 3p21.3 [
64], whereas Hyal-4, PHYAL1, and SPAM1 are found in chromosome 7q31.3 [
65]. Hyal-1, Hyal-2, and PH-20 regulate HA’s stability [
66], whereas Hyal-3, Hyal-4, and PHYAL1 do not affect HA’s metabolism [
67]. Among all Hyals, human Hyal-1 is abundant in the somatic tissue and in human serum (~60 ng/mL) as well as in liver, kidney, heart, plasma etc. This lysosomal enzyme contains three
N-glycosylated sites at places N-99, N-216, and N-350 [
68] and degrades HA into GlcA-GlcNAc-GlcA-GlcNAc tetrasacharides [
61]. According to numerous studies, Hyal-1 is expressed in various cancer cells, like in the prostate, bladder, and brain [
69,
70], and is also connected to the human cancer cell lines MCF-7 and MDA-MB-231 [
71]. Hyal-1 is of great interest, as a well-developed study is determining compounds which act as natural Hyal-1 inhibitors against diseases like arthritis and gingivitis [
72,
73]. Hyal-2 is widely found in connective tissue at high concentration and decomposes HA into oligomers of 50 saccharide units (~20 kDa) [
74]. The activity of Hyal-2 ranges between various pH values, whereas Hyal- 1 and Hyal-3 are active in acidic conditions. Hyal-4 behaves also as a chondroitinase, using Chondroitin (Ch) and Chondroitin Sulfate (ChS) as substrates. Additionally, SPAM1 is located in the acrosome of spermatids and enables the adhesion of sperm to the cumulus mass protecting the ovum [
74,
75].
Hyals’ catalysis depends on an acidic active site Glu-131, which donates H [
60]. Hyal’s active site also contains Tyr75 Trp141, Tyr201, Tyr208, Tyr210, Tyr247, Tyr261, Tyr286, and Trp321, which are responsible for its catalysis. The active site, positively charged hydrophobic amino acids enable the complete interaction not only with HA, but also with other negatively charged substrates (e.g., Chondroitin and Chondroitin Sulfate) [
74]. Furthermore, Hyal’s structure contains two arginines (Arg134 and Arg265), which interact with -COOH groups of HA, as well as a Ser245, which is stable and enables bond formation with the −OH group of Tyr202 [
61,
68,
74]. Hyal-4 has Cys263 instead of Tyr247, which enables the interaction with Chondroitin and Chondroitin Sulfate [
74].
Numerous studies regarding the anti-Hyal potency of plants, their extracts, or their isolated metabolites have showed that the most potent natural inhibitors are terpenes, flavonoids and alkanoids [
60].
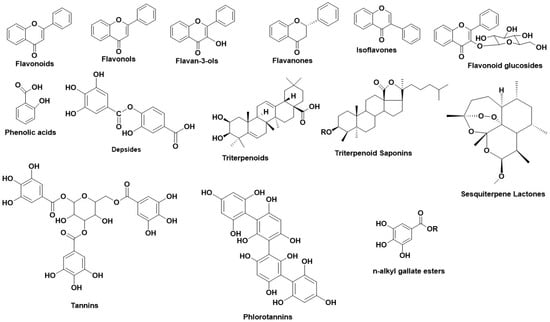
Figure 2 illustrates the main structural scaffolds which have been marked as efficient Hyal inhibitors.
The natural products that have been studied for their inhibitory properties towards Hyal can be explored in the new developed Server and Database called ANTIAGE-DB, through the link https://bio-hpc.ucam.edu/anti-age-db.
Figure 2. Main structural scaffolds which have been marked as potent Hyal inhibitors.
3. Tyrosinase—A Polyphenol Oxidase
Tyrosinases (catechol oxidases, catecholases, diphenol oxidases,
o-diphenolases, phenolases and polyphenol oxidases) [ΕC 1.14.18.1] are copper-containing glycoproteins and can be found in various active forms in living organisms (mammals, plants, fungi and animal tissues) [
86]. The most vital activity of Tyrs is their implication in melanin and other pigments synthesis, as well as the brown color in fruits and vegetables, caused by the oxidation of the present Tyr in their flesh with the atmospheric molecular oxygen. In humans, Tyrs are located in the melanosomes, which are synthesized in the skin melanocytes. Tyrs have a double enzymatic activity, in the presence of O
2: (1) they act as monophenolases (cresolases) and hydroxylate monophenols in their ortho-position and (2) they act as diphenolases (catecholases) and oxidize
o-diphenols into
o-quinones. Tyr oxidizes L-tyrosine into dopaquinone, a product which leads to the formation of eumelanin (brown or black melanin) and pheomelanin (red to yellow melanin) [
86].
Specifically, Tyr structure consists of three domain parts. The central, and most characteristic domain, is the active site binuclear copper moiety, which enables the bond formation between Tyr and molecular O
2, as well as with Tyr’s substrate [
87]. The other domains contain six histidines, which form bonds with the cuprous pair inside the active site. The Tyr structure is stabilized through disoulfide bridges, formed by cysteines around the active site. Tyrs from different plants differ in their number of cysteines; human and mouse Tyrs have 17 cysteines, whereas plant Tyrs have 11 cysteines. Furthermore, plant Tyrs contain one cysteine in their Carboxylic terminal moiety, whereas Tyrs from
N. crassa,
A. bisporus, and prokaryotic Tyrs have 0 or 1 cysteines [
88].
Numerous studies revealed various natural metabolites as Tyr inhibitors, along with their interacting properties with Tyr’s active site through combined studies on docking simulations and kinetics [
89]. The structure of Tyr is similar to hemoqyanin, and a lot of correlated results regarding the binding properties of Tyr can be excluded [
89,
90,
91]. Wang et al., developed a fluorescent probe with which they determined the mechanism of Tyr reaction [
92] finding that the inhibitors react with Tyr irreversibly [
86]. Tyr inhibition is achieved either by covering the active site or by producing undesirable dopaquinone derivatives or by completing multisite reactions. For this reason, several studies have been performed regarding the inhibitory activity of natural compounds, their type of inhibition, including also their interaction profile.
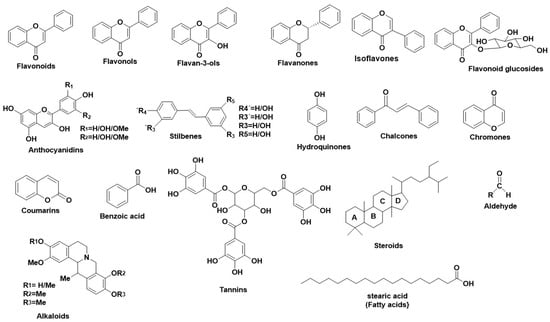
Figure 3 illustrates the main structural scaffolds which have been marked as efficient Tyr inhibitors. The natural products that have been studied for their inhibitory properties towards Tyr can be explored in the new developed Server and Database called ANTIAGE-DB, through the link
https://bio-hpc.ucam.edu/anti-age-db.
Figure 3. Main structural scaffolds which have been marked as potent Tyr inhibitors.
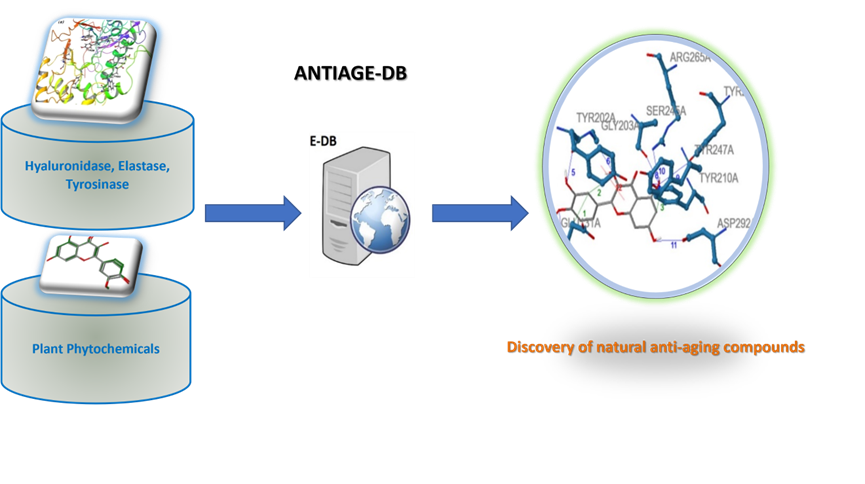
The Database ANTIAGE-DB has been developed to allow a flexible and easy interrogation of the inhibition properties of natural products as also to perform in real time in silico prediction of targeted compounds that can interact with the studied enzymes (HNE, Hyal and Tyr). Also, the Server provides information on the atomic level interactions of the targeted compounds with the three enzymes (HNE, Hyal and Tyr) as also the calculated binding affinity of these targeted compounds towards each enzyme (HNE, Hyal and Tyr) (Figure 4).
Figure 4: The ANTIAGE-DB.
This entry is adapted from the peer-reviewed paper 10.3390/antiox11112268