Neuropsychiatric disease is a serious public health problem in the world. Depression and anxiety are two of the most common neuropsychiatric diseases, and they usually occur simultaneously or sequentially. There are nearly 322 million people suffering from depression in the world and it has become the second leading cause of years lost to disability worldwide. Depression is characterized by fatigue, anhedonia, sleep disturbance, and self-destructive behavior, which seriously affect patients’ mental health and social functions. The pathology of depression was associated with neuroinflammation, monoamine deficiency, gut microbiota disorder, the hyperactivity of the HPA axis, and so on.
- dietary natural products
- nutrients
- depression
- gut microbiota
1. Epidemiological Studies
2. Experimental Studies
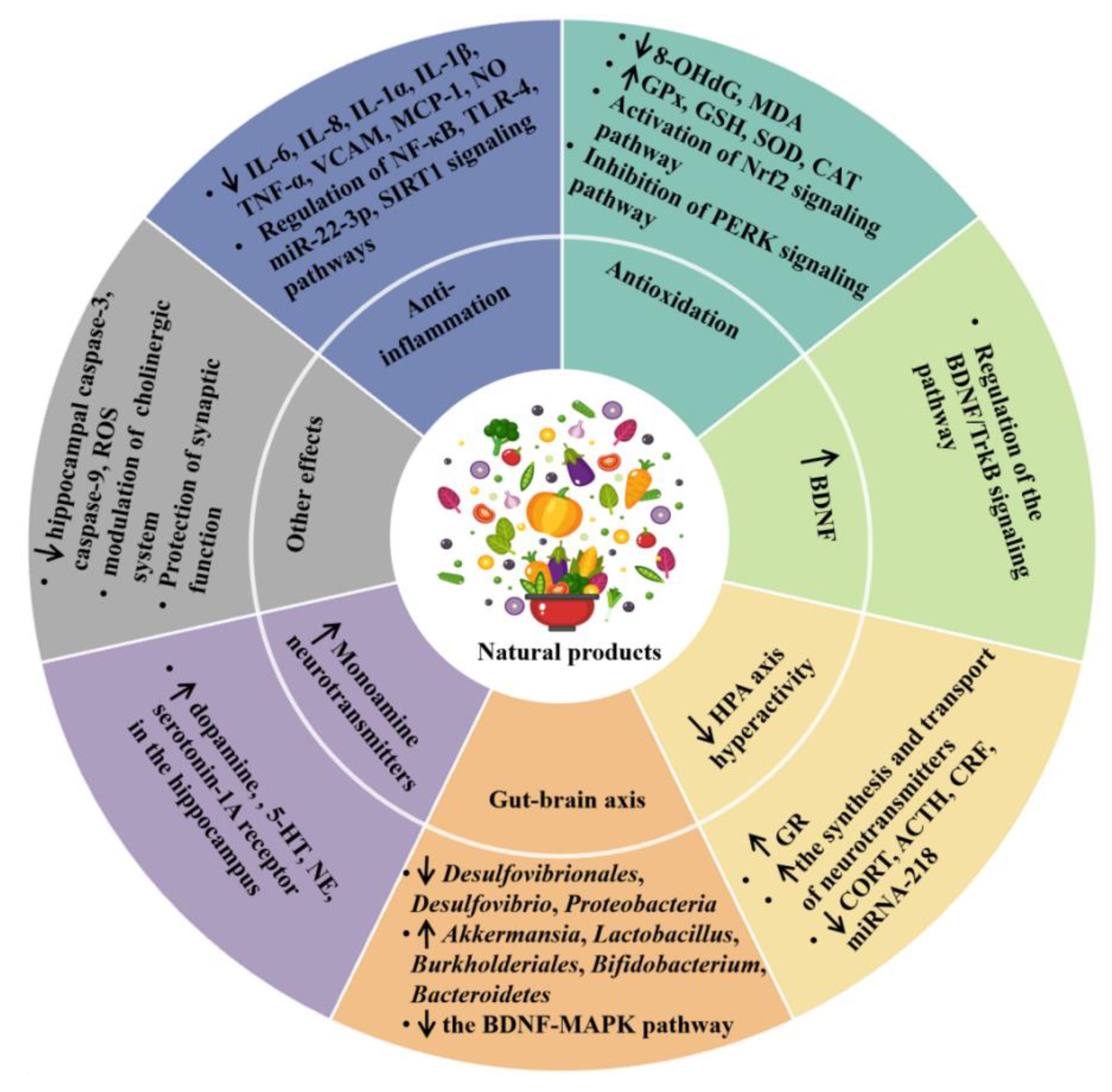
2.1. Anti-Inflammatory Effect
2.2. Antioxidant Effect
2.3. Modulating the Production of Monoamine Neurotransmitters
2.4. Promoting the Production of Neurotrophins
2.5. Inhibition of the HPA Axis Hyperactivity
2.6. Modulation of Microbiota–Gut–Brain Axis
2.7. Other Mechanisms
3. Clinical Trials
This entry is adapted from the peer-reviewed paper 10.3390/antiox11112132
References
- Ng, T.P.; Gao, Q.; Gwee, X.; Chua, D.Q.L. Tea consumption and depression from follow up in the singapore longitudinal ageing study. J. Nutr. Health Aging 2021, 25, 295–301.
- Kim, J.; Kim, J. Green tea, coffee, and caffeine consumption are inversely associated with self-report lifetime depression in the korean population. Nutrients 2018, 10, 1201.
- Liu, Y.Y.; Ju, Y.; Cui, L.L.; Liu, T.; Hou, Y.Y.; Wu, Q.; Ojo, O.; Du, X.J.; Wang, X.H. Association between dietary fiber intake and incidence of depression and anxiety in patients with essential hypertension. Nutrients 2021, 13, 4159.
- de Almeida, T.L.F.; Petarli, G.B.; Cattafesta, M.; Zandonade, E.; Bezerra, O.; Tristao, K.G.; Salaroli, L.B. Association of selenium intake and development of depression in Brazilian farmers. Front. Nutr. 2021, 8, 671377.
- Saghafian, F.; Malmir, H.; Saneei, P.; Milajerdi, A.; Larijani, B.; Esmaillzadeh, A. Fruit and vegetable consumption and risk of depression: Accumulative evidence from an updated systematic review and meta-analysis of epidemiological studies. Br. J. Nutr. 2018, 119, 1087–1101.
- Kim, C.S.; Byeon, S.; Shin, D.M. Sources of dietary fiber are differently associated with prevalence of depression. Nutrients 2020, 12, 2813.
- Arab, L.; Guo, R.; Elashoff, D. Lower depression scores among walnut consumers in nhanes. Nutrients 2019, 11, 275.
- Yang, Y.J.; Je, Y.J. Fish consumption and depression in Korean adults: The Korea National Health and Nutrition Examination Survey, 2013–2015. Eur. J. Clin. Nutr. 2018, 72, 1142–1149.
- Sanchez-Villegas, A.; Alvarez-Perez, J.; Toledo, E.; Salas-Salvado, J.; Ortega-Azorin, C.; Zomeno, M.D.; Vioque, J.; Martinez, J.A.; Romaguera, D.; Perez-Lopez, J.; et al. Seafood consumption, omega-3 fatty acids intake, and life-time prevalence of depression in the PREDIMED-plus trial. Nutrients 2018, 10, 2000.
- Navarro, A.M.; Abasheva, D.; Martinez-Gonzalez, M.A.; Ruiz-Estigarribia, L.; Martin-Calvo, N.; Sanchez-Villegas, A.; Toledo, E. Coffee consumption and the risk of depression in a middle-aged cohort: The SUN project. Nutrients 2018, 10, 1333.
- Yao, S.; Zhang, M.; Dong, S.-S.; Wang, J.-H.; Zhang, K.; Guo, J.; Guo, Y.; Yang, T.-L. Bidirectional two-sample Mendelian randomization analysis identifies causal associations between relative carbohydrate intake and depression. Nat. Hum. Behav. 2022, in press.
- Anjom-Shoae, J.; Sadeghi, O.; Keshteli, A.H.; Afshar, H.; Esmaillzadeh, A.; Adibi, P. Legume and nut consumption in relation to depression, anxiety and psychological distress in Iranian adults. Eur. J. Nutr. 2020, 59, 3635–3645.
- Zhang, X.Y.; Huang, X.Y.; Xiao, Y.; Jing, D.R.; Huang, Y.Z.; Chen, L.P.; Luo, D.; Chen, X.; Shen, M.X. Daily intake of soft drinks is associated with symptoms of anxiety and depression in Chinese adolescents. Public Health Nutr. 2019, 22, 2553–2560.
- Sanchez-Villegas, A.; Zazpe, I.; Santiago, S.; Perez-Cornago, A.; Martinez-Gonzalez, M.A.; Lahortiga-Ramos, F. Added sugars and sugar-sweetened beverage consumption, dietary carbohydrate index and depression risk in the Seguimiento Universidad de Navarra (SUN) Project. Br. J. Nutr. 2018, 119, 211–221.
- Luo, M.; Gan, R.-Y.; Li, B.-Y.; Mao, Q.-Q.; Shang, A.; Xu, X.-Y.; Li, H.-Y.; Li, H.-B. Effects and mechanisms of tea on parkinson’s disease, alzheimer’s disease and depression. Food Rev. Int. 2021, in press.
- Zhou, L.; Wang, T.; Yu, Y.; Li, M.; Sun, X.; Song, W.; Wang, Y.; Zhang, C.; Fu, F. The etiology of poststroke-depression: A hypothesis involving HPA axis. Biomed. Pharmacother. 2022, 151, 113146.
- Ren, Y.F.; Sun-Waterhouse, D.; Ouyang, F.X.; Tan, X.T.; Li, D.P.; Xu, L.H.; Li, B.; Wang, Y.L.; Li, F. Apple phenolic extracts ameliorate lead-induced cognitive impairment and depression- and anxiety-like behavior in mice by abating oxidative stress, inflammation and apoptosis via the miR-22-3p/SIRT1 axis. Food Funct. 2022, 13, 2647–2661.
- Jeon, S.W.; Kim, Y.-K. Inflammation-induced depression: Its pathophysiology and therapeutic implications. J. Neuroimmunol. 2017, 313, 92–98.
- Norris, G.T.; Kipnis, J. Immune cells and CNS physiology: Microglia and beyond. J. Exp. Med. 2019, 216, 60–70.
- Choi, J.E.; Borkowski, K.; Newman, J.W.; Park, Y.S. N-3 PUFA improved post-menopausal depression induced by maternal separation and chronic mild stress through serotonergic pathway in rats-effect associated with lipid mediators. J. Nutr. Biochem. 2021, 91, 108599.
- Dong, X.F.; Lu, K.; Lin, P.C.; Che, H.X.; Li, H.Y.; Song, L.; Yang, X.H.; Xie, W.C. Saccharina japonica ethanol extract ameliorates depression/anxiety-like behavior by inhibiting inflammation, oxidative stress, and apoptosis in dextran sodium sulfate induced ulcerative colitis mice. Front. Nutr. 2021, 8, 784532.
- Huang, F.R.; Liu, X.J.; Xu, S.; Hu, S.T.; Wang, S.S.; Shi, D.B.; Wang, K.C.; Wang, Z.X.; Lin, Q.Q.; Li, S.; et al. Prevotella histicola mitigated estrogen deficiency-induced depression via gut microbiota-dependent modulation of inflammation in ovariectomized mice. Front. Nutr. 2022, 8, 805465.
- Yun, S.W.; Kim, J.K.; Han, M.J.; Kim, D.H. Lacticaseibacillus paracasei NK112 mitigates Escherichia coli-induced depression and cognitive impairment in mice by regulating IL-6 expression and gut microbiota. Benef. Microbes 2021, 12, 541–551.
- Wang, J.P.; Xu, S.; Chen, X.H.; Wang, L.; Li, J.Y.; Li, G.J.; Zhang, B.L. Antidepressant effect of EGCG through the inhibition of hippocampal neuroinflammation in chronic unpredictable mild stress-induced depression rat model. J. Funct. Foods 2020, 73, 104106.
- Bhatt, S.; Nagappa, A.N.; Patil, C.R. Role of oxidative stress in depression. Drug Discov. Today 2020, 25, 1270–1276.
- Ou, S.S.; Fang, Y.C.; Tang, H.; Wu, T.; Chen, L.Z.; Jiang, M.; Zhou, L.Q.; Xu, J.; Guo, K.H. Lycopene protects neuroblastoma cells against oxidative damage via depression of ER stress. J. Food Sci. 2020, 85, 3552–3561.
- Moosa, A.; Farzand, A.; Ahmad, T.; Khan, S.A.; Gleason, M.L.; Abbas, H.; Khan, W.A.; Jabbar, A.; Mohsan, M. First report of fusarium rot of phalsa (Grewia asiatica) caused by Fusarium solani in Pakistan. Plant Dis. 2019, 103, 2476–2477.
- Imran, I.; Javaid, S.; Waheed, A.; Rasool, M.F.; Majeed, A.; Samad, N.; Saeed, H.; Alqahtani, F.; Ahmed, M.M.; Alaqil, F.A. Grewia asiatica berry juice diminishes anxiety, depression, and scopolamine-induced learning and memory impairment in behavioral experimental animal models. Front. Nutr. 2021, 7, 587367.
- Di Lorenzo, A.; Sobolev, A.P.; Nabavi, S.F.; Sureda, A.; Moghaddam, A.H.; Khanjani, S.; Di Giovanni, C.; Xiao, J.B.; Shirooie, S.; Sokeng, A.J.T.; et al. Antidepressive effects of a chemically characterized maqui berry extract (Aristotelia chilensis (Molina) Stuntz) in a mouse model of post-stroke depression. Food Chem. Toxicol. 2019, 129, 434–443.
- Cigliano, L.; Spagnuolo, M.S.; Boscaino, F.; Ferrandino, I.; Monaco, A.; Capriello, T.; Cocca, E.; Iannotta, L.; Treppiccione, L.; Luongo, D.; et al. Dietary supplementation with fish oil or conjugated linoleic acid relieves depression markers in mice by modulation of the nrf2 pathway. Mol. Nutr. Food Res. 2019, 63, 1900243.
- Marathe, S.V.; D’Almeida, P.L.; Virmani, G.; Bathini, P.; Alberi, L. Effects of monoamines and antidepressants on astrocyte physiology: Implications for monoamine hypothesis of depression. J. Exp. Neurosci. 2018, 12, 1179069518789149.
- Lu, X.M.; Ce, Q.; Jin, L.; Zheng, J.; Sun, M.; Tang, X.; Li, D.; Sun, J. Deoiled sunflower seeds ameliorate depression by promoting the production of monoamine neurotransmitters and inhibiting oxidative stress. Food Funct. 2021, 12, 573–586.
- Li, G.J.; Yang, J.; Wang, X.; Zhou, C.; Zheng, X.Y.; Lin, W.H. Effects of EGCG on depression-related behavior and serotonin concentration in a rat model of chronic unpredictable mild stress. Food Funct. 2020, 11, 8780–8787.
- Zhang, L.L.; Yang, Z.Y.; Ren, J.N.; Fan, G.; Pan, S.Y. Dietary essential oil from navel orange alleviates depression in reserpine-treated mice by monoamine neurotransmitters. Flavour Frag. J. 2019, 34, 252–259.
- Wu, Z.; Wang, P.Y.; Pan, D.D.; Zeng, X.Q.; Guo, Y.X.; Zhao, G.S. Effect of adzuki bean sprout fermented milk enriched in gamma-aminobutyric acid on mild depression in a mouse model. J. Dairy Sci. 2021, 104, 78–91.
- Sahay, A.; Kale, A.; Joshi, S. Role of neurotrophins in pregnancy and offspring brain development. Neuropeptides 2020, 83, 102075.
- Huang, Y.J.; Lu, K.H.; Lin, Y.E.; Panyod, S.; Wu, H.Y.; Chang, W.T.; Sheen, L.Y. Garlic essential oil mediates acute and chronic mild stress-induced depression in rats via modulation of monoaminergic neurotransmission and brain-derived neurotrophic factor levels. Food Funct. 2019, 10, 8094–8105.
- Lim, D.W.; Han, T.; Um, M.Y.; Yoon, M.; Kim, T.E.; Kim, Y.T.; Han, D.; Lee, J.; Lee, C.H. Administration of Asian herb bennet (Geum japonicum) extract reverses depressive -like behaviors in mouse model of depression induced by corticosterone. Nutrients 2019, 11, 2841.
- Fang, J.L.; Luo, Y.; Jin, S.H.; Yuan, K.; Guo, Y. Ameliorative effect of anthocyanin on depression mice by increasing monoamine neurotransmitter and up-regulating BDNF expression. J. Funct. Foods 2020, 66, 103757.
- Zhao, Y.; Wang, Q.; Jia, M.; Fu, S.; Pan, J.; Chu, C.; Liu, X.; Liu, X.; Liu, Z. (+)-Sesamin attenuates chronic unpredictable mild stress-induced depressive-like behaviors and memory deficits via suppression of neuroinflammation. J. Nutr. Biochem. 2019, 64, 61–71.
- Aygun, H.; Akin, A.T.; Kizilaslan, N.; Sumbul, O.; Karabulut, D. Probiotic supplementation alleviates absence seizures and anxiety- and depression-like behavior in WAG/Rij rat by increasing neurotrophic factors and decreasing proinflammatory cytokines. Epilepsy Behav. 2022, 128, 108588.
- Bin-Jaliah, I.; Al-Hash, F.H.; Abbas, A.M.; Nasif, K.A.; Sakr, H.F. Effect of resveratrol on neurotrophic factors and depression-like behavior in a rat model of male hypogonadism. Int. J. Pharmacol. 2021, 17, 38–47.
- Bao, A.-M.; Swaab, D.F. The human hypothalamus in mood disorders: The HPA axis in the center. IBRO Rep. 2019, 6, 45–53.
- Wang, C.C.; Du, L.; Shi, H.H.; Ding, L.; Yanagita, T.; Xue, C.H.; Wang, Y.M.; Zhang, T.T. Dietary EPA-enriched phospholipids alleviate chronic stress and LPS-induced depression- and anxiety-like behavior by regulating immunity and neuroinflammation. Mol. Nutr. Food Res. 2021, 65, 2100009.
- Jia, D.; Dou, Y.H.; He, Y.S.; Zhou, X.X.; Gao, Y.; Ma, M.; Wu, Z.Z.; Li, W.M. Saponin extract of Baihe-Zhimu Tang ameliorates depression in chronic mild stress rats. J. Funct. Foods 2020, 68, 103905.
- Choi, J.E.; Kim, E.Y.; Park, Y. N-3 PUFA improved pup separation-induced postpartum depression via serotonergic pathway regulated by miRNA. J. Nutr. Biochem. 2020, 84, 108417.
- Iegaki, N.; Narita, Y.; Hattori, N.; Hirata, Y.; Ichihara, K. Royal jelly reduces depression-like behavior through possible effects on adrenal steroidogenesis in a murine model of unpredictable chronic mild stress. Biosci. Biotechnol. Biochem. 2020, 84, 606–612.
- Tao, J.; Li, S.; Gan, R.-Y.; Zhao, C.-N.; Meng, X.; Li, H.-B. Targeting gut microbiota with dietary components on cancer: Effects and potential mechanisms of action. Crit. Rev. Food Sci. Nutr. 2020, 60, 1025–1037.
- Li, H.-Y.; Zhou, D.-D.; Gan, R.-Y.; Huang, S.-Y.; Zhao, C.-N.; Shang, A.; Xu, X.-Y.; Li, H.-B. Effects and mechanisms of probiotics, prebiotics, synbiotics, and postbiotics on metabolic diseases targeting gut microbiota: A narrative review. Nutrients 2021, 13, 3211.
- Li, B.; Mao, Q.; Zhou, D.; Luo, M.; Gan, R.; Li, H.; Huang, S.; Saimaiti, A.; Shang, A.; Li, H. Effects of tea against alcoholic fatty liver disease by modulating gut microbiota in chronic alcohol-exposed mice. Foods 2021, 10, 1232.
- Li, B.-Y.; Xu, X.-Y.; Gan, R.-Y.; Sun, Q.-C.; Meng, J.-M.; Shang, A.; Mao, Q.-Q.; Li, H.-B. Targeting gut microbiota for the prevention and management of diabetes mellitus by dietary natural products. Foods 2019, 8, 440.
- Zhang, Y.-J.; Li, S.; Gan, R.-Y.; Zhou, T.; Xu, D.-P.; Li, H.-B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519.
- Luo, M.; Zhou, D.-D.; Shang, A.; Gan, R.-Y.; Li, H.-B. Influences of food contaminants and additives on gut microbiota as well as protective effects of dietary bioactive compounds. Trends Food Sci. Technol. 2021, 113, 180–192.
- Cao, S.-Y.; Zhao, C.-N.; Xu, X.-Y.; Tang, G.-Y.; Corke, H.; Gan, R.-Y.; Li, H.-B. Dietary plants, gut microbiota, and obesity: Effects and mechanisms. Trends Food Sci. Technol. 2019, 92, 194–204.
- Capuco, A.; Urits, I.; Hasoon, J.; Chun, R.; Gerald, B.; Wang, J.K.; Ngo, A.L.; Simopoulos, T.; Kaye, A.D.; Colontonio, M.M.; et al. Gut microbiome dysbiosis and depression: A comprehensive review. Curr. Pain Headache Rep. 2020, 24, 36.
- Liu, Y.; Wu, Z.F.; Cheng, L.; Zhang, X.; Yang, H.N. The role of the intestinal microbiota in the pathogenesis of host depression and mechanism of TPs relieving depression. Food Funct. 2021, 12, 7651–7663.
- Yang, Y.; Zhao, S.; Yang, X.; Li, W.; Si, J.; Yang, X. The antidepressant potential of Lactobacillus casei in the postpartum depression rat model mediated by the microbiota-gut-brain axis. Neurosci. Lett. 2022, 774, 136474.
- Gu, F.; Wu, Y.Y.; Liu, Y.; Dou, M.; Jiang, Y.S.; Liang, H. Lactobacillus casei improves depression-like behavior in chronic unpredictable mild stress-induced rats by the BDNF-TrkB signal pathway and the intestinal microbiota. Food Funct. 2020, 11, 6148–6157.
- Tian, P.J.; Wang, G.; Zhao, J.X.; Zhang, H.; Chen, W. Bifidobacterium with the role of 5-hydroxytryptophan synthesis regulation alleviates the symptom of depression and related microbiota dysbiosis. J. Nutr. Biochem. 2019, 66, 43–51.
- Sun, Y.; Geng, W.T.; Pan, Y.J.; Wang, J.J.; Xiao, P.; Wang, Y.P. Supplementation with Lactobacillus kefiranofaciens ZW3 from Tibetan Kefir improves depression- like behavior in stressed mice by modulating the gut microbiota. Food Funct. 2019, 10, 925–937.
- Yun, S.W.; Kim, J.K.; Lee, K.E.; Oh, Y.J.; Choi, H.J.; Han, M.J.; Kim, D.H. A probiotic Lactobacillus gasseri alleviates Escherichia coli-induced cognitive impairment and depression in mice by regulating IL-1 beta expression and gut microbiota. Nutrients 2020, 12, 3441.
- Liu, Z.G.; Li, L.; Ma, S.B.; Ye, J.; Zhang, H.B.; Li, Y.T.; Sair, A.T.; Pan, J.R.; Liu, X.N.; Li, X.; et al. High-dietary fiber intake alleviates antenatal obesity-induced postpartum depression: Roles of gut microbiota and microbial metabolite short-chain fatty acid involved. J. Agric. Food Chem. 2020, 68, 13697–13710.
- Song, J.; Zhou, N.A.; Ma, W.N.; Gu, X.Y.; Chen, B.Z.; Zeng, Y.; Yang, L.; Zhou, M.M. Modulation of gut microbiota by chlorogenic acid pretreatment on rats with adrenocorticotropic hormone induced depression-like behavior. Food Funct. 2019, 10, 2947–2957.
- Han, S.K.; Joo, M.K.; Kim, J.K.; Jeung, W.; Kang, H.; Kim, D.H. Bifidobacteria-fermented red ginseng and its constituents ginsenoside Rd and protopanaxatriol alleviate anxiety/depression in mice by the amelioration of gut dysbiosis. Nutrients 2020, 12, 901.
- Braidy, N.; Liu, Y. Can nicotinamide riboside protect against cognitive impairment? Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 413–420.
- Jiang, Y.S.; Liu, Y.; Gao, M.Q.; Xue, M.L.; Wang, Z.L.; Liang, H. Nicotinamide riboside alleviates alcohol-induced depression-like behaviours in C57BL/6J mice by altering the intestinal microbiota associated with microglial activation and BDNF expression. Food Funct. 2020, 11, 378–391.
- Wang, L.; Wu, X.J.; Ma, Y.H.; Li, X.; Zhang, J.F.; Zhao, L.C. Supplementation with soy isoflavones alleviates depression-like behaviour via reshaping the gut microbiota structure. Food Funct. 2021, 12, 4995–5006.
- Hao, W.Z.; Ma, Q.Y.; Tao, G.; Huang, J.Q.; Chen, J.X. Oral coniferyl ferulate attenuated depression symptoms in mice via reshaping gut microbiota and microbial metabolism. Food Funct. 2021, 12, 12550–12564.
- Chen, Y.L.; Xiao, N.; Chen, Y.X.; Chen, X.Y.; Zhong, C.F.; Cheng, Y.Y.; Du, B.; Li, P. Semen Sojae Praeparatum alters depression-like behaviors in chronic unpredictable mild stress rats via intestinal microbiota. Food Res. Int. 2021, 150, 110808.
- Maehata, H.; Kobayashi, Y.; Mitsuyama, E.; Kawase, T.; Kuhara, T.; Xiao, J.Z.; Tsukahara, T.; Toyoda, A. Heat-killed Lactobacillus helveticus strain MCC1848 confers resilience to anxiety or depression-like symptoms caused by subchronic social defeat stress in mice. Biosci. Biotechnol. Biochem. 2019, 83, 1239–1247.
- Chen, S.X.; Guo, W.Y.; Qi, X.X.; Zhou, J.Y.; Liu, Z.Q.; Cheng, Y.Y. Natural alkaloids from lotus plumule ameliorate lipopolysaccharide-induced depression-like behavior: Integrating network pharmacology and molecular mechanism evaluation. Food Funct. 2019, 10, 6062–6073.
- Fan, C.Q.; Li, Y.; Lan, T.; Wang, W.J.; Mao, X.Q.; Yu, S.Y. Prophylactic treatment of curcumin in a rat model of depression by attenuating hippocampal synaptic loss. Food Funct. 2021, 12, 11202–11213.
- Xiong, R.-G.; Zhou, D.-D.; Wu, S.-X.; Huang, S.-Y.; Saimaiti, A.; Yang, Z.-J.; Shang, A.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Health benefits and side effects of short-chain fatty acids. Foods 2022, 11, 2863.
- Tang, C.-F.; Wang, C.-Y.; Wang, J.-H.; Wang, Q.-N.; Li, S.-J.; Wang, H.-O.; Zhou, F.; Li, J.-M. Short-chain fatty acids ameliorate depressive-like behaviors of high fructose-fed mice by rescuing hippocampal neurogenesis decline and blood-brain barrier damage. Nutrients 2022, 14, 1882.
- Johnston, C.S.; Jasbi, P.; Jin, Y.; Bauer, S.; Williams, S.; Fessler, S.N.; Gu, H.W. Daily vinegar ingestion improves depression scores and alters the metabolome in healthy adults: A randomized controlled trial. Nutrients 2021, 13, 4020.
- Poorbaferani, F.; Rouhani, M.H.; Heidari, Z.; Poorbaferani, M.; Safavi, S.M. Flaxseed oil supplementation on severity of depression and brain-derived neurotrophic factor: A randomized, double blind placebo controlled clinical trial. Int. J. Food Prop. 2020, 23, 1518–1526.
- Xue, L.L.; Zhang, J.; Shen, H.X.; Ai, L.; Wu, R.R. A randomized controlled pilot study of the effectiveness of magnolia tea on alleviating depression in postnatal women. Food Sci. Nutr. 2020, 8, 1554–1561.
- Fisk, J.; Khalid, S.; Reynolds, S.A.; Williams, C.M. Effect of 4 weeks daily wild blueberry supplementation on symptoms of depression in adolescents. Br. J. Nutr. 2020, 124, 181–188.
- Zhang, X.M.; Chen, S.B.; Zhang, M.; Ren, F.Z.; Ren, Y.M.; Li, Y.X.; Liu, N.; Zhang, Y.; Zhang, Q.; Wang, R. Effects of fermented milk containing Lacticaseibacillus paracasei strain shirota on constipation in patients with depression: A randomized, double-blind, placebo-controlled trial. Nutrients 2021, 13, 2238.
- Kim, H.K.; Kim, S.H.; Jang, C.S.; Kim, S.I.; Kweon, C.O.; Kim, B.W.; Ryu, J.K. The combined effects of yogurt and exercise in healthy adults: Implications for biomarkers of depression and cardiovascular diseases. Food Sci. Nutr. 2018, 6, 1968–1974.
- Ali, S.; Corbi, G.; Maes, M.; Scapagnini, G.; Davinelli, S. Exploring the impact of flavonoids on symptoms of depression: A systematic review and meta-analysis. Antioxidants 2021, 10, 1644.
- Marx, W.; Lane, M.; Rocks, T.; Ruusunen, A.; Loughman, A.; Lopresti, A.; Marshall, S.; Berk, M.; Jacka, F.; Dean, O.M. Effect of saffron supplementation on symptoms of depression and anxiety: A systematic review and meta-analysis. Nutr. Rev. 2019, 77, 557–571.